
Forensic DNA Analysis of Mixed Mosquito Blood Meals: STR Profiling for Human Identification
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Mosquitoes
2.2. Experimental Mice
2.3. Blood Samples
2.3.1. Human Blood
2.3.2. Mice Blood
2.4. Mosquito Membrane Feeding
2.5. Experimental Design
2.6. DNA Extraction and Quanitation
2.7. STRs Profiling
2.8. Data Analysis
3. Results
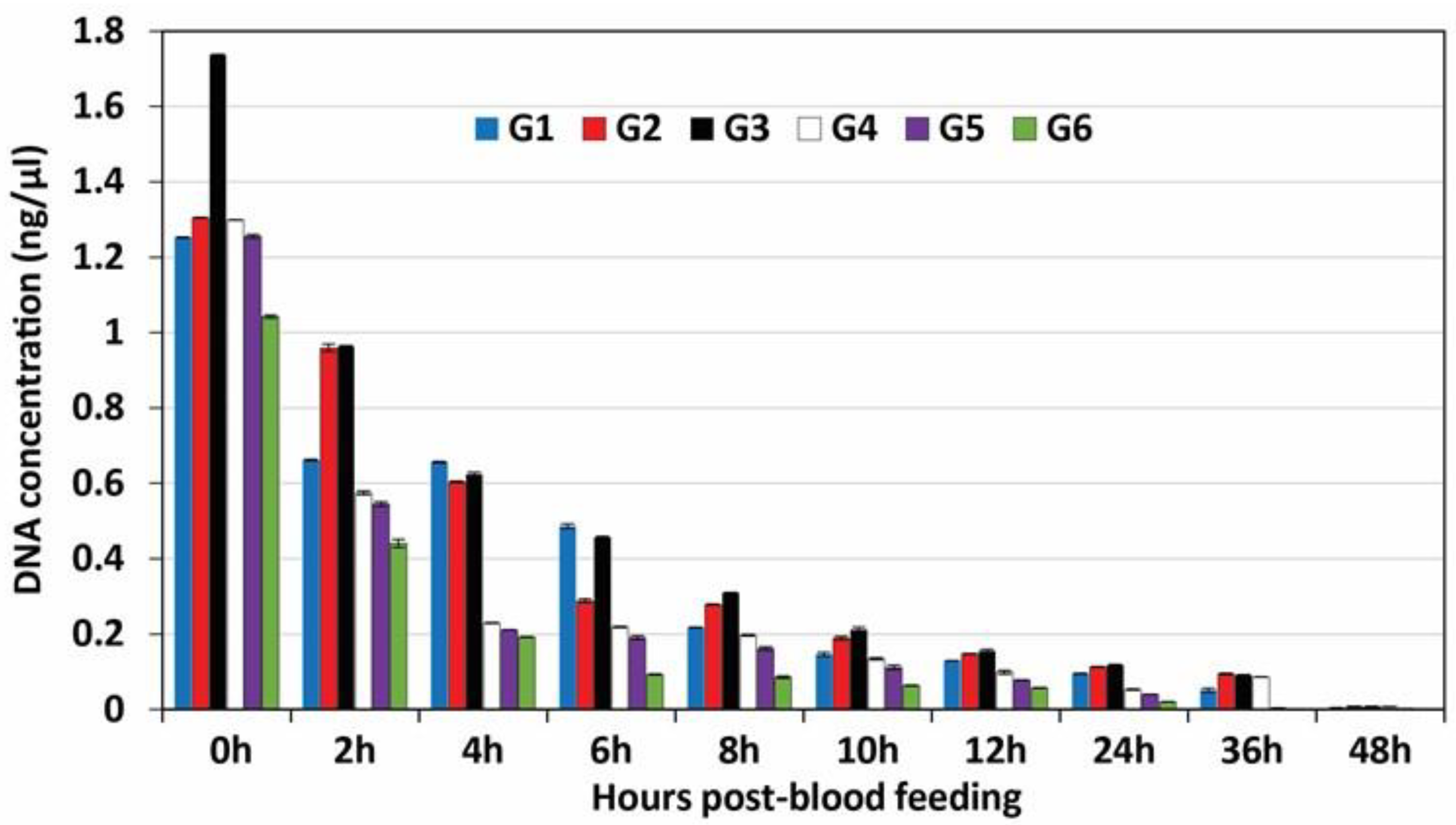
3.1. DNA Quantification
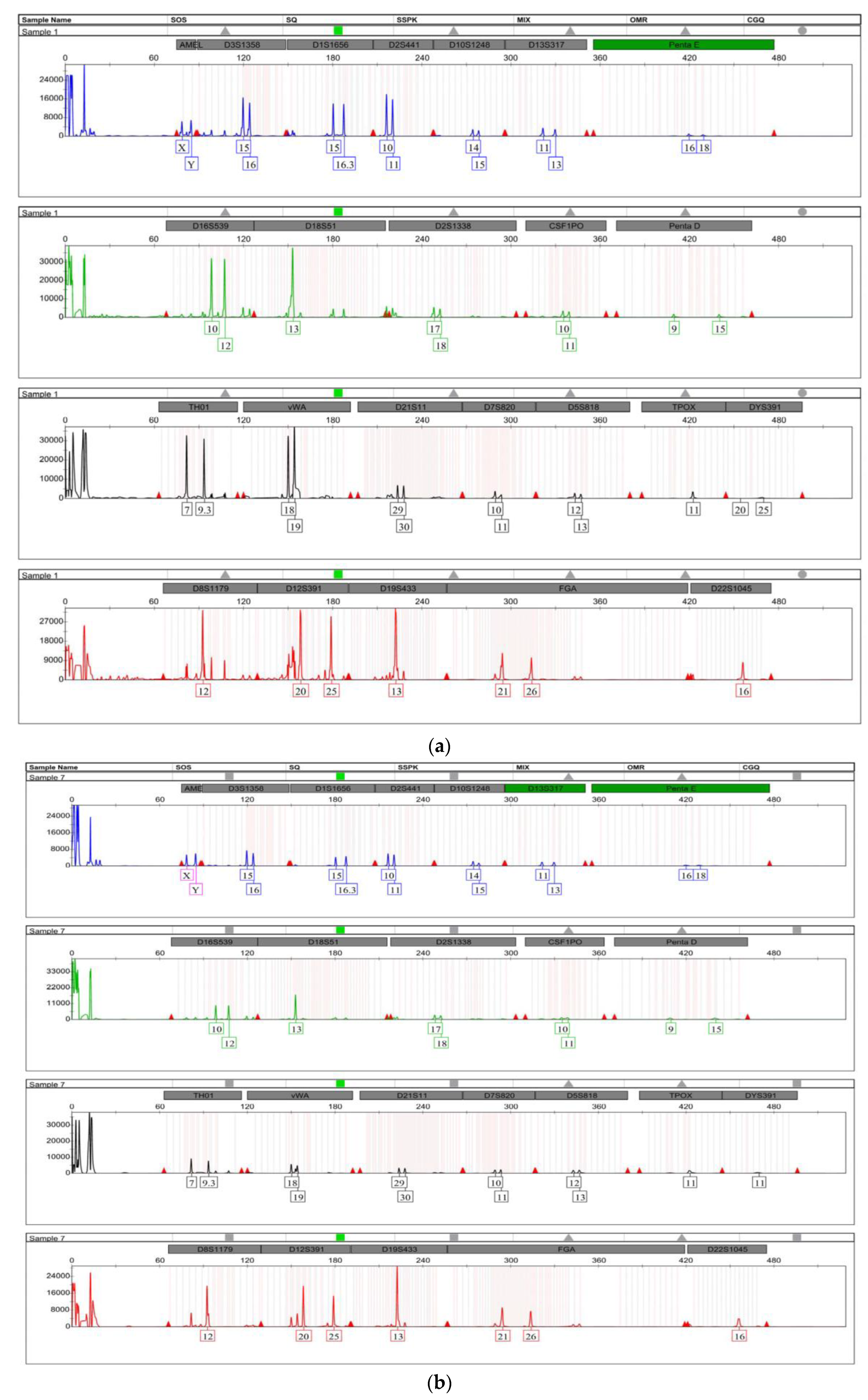
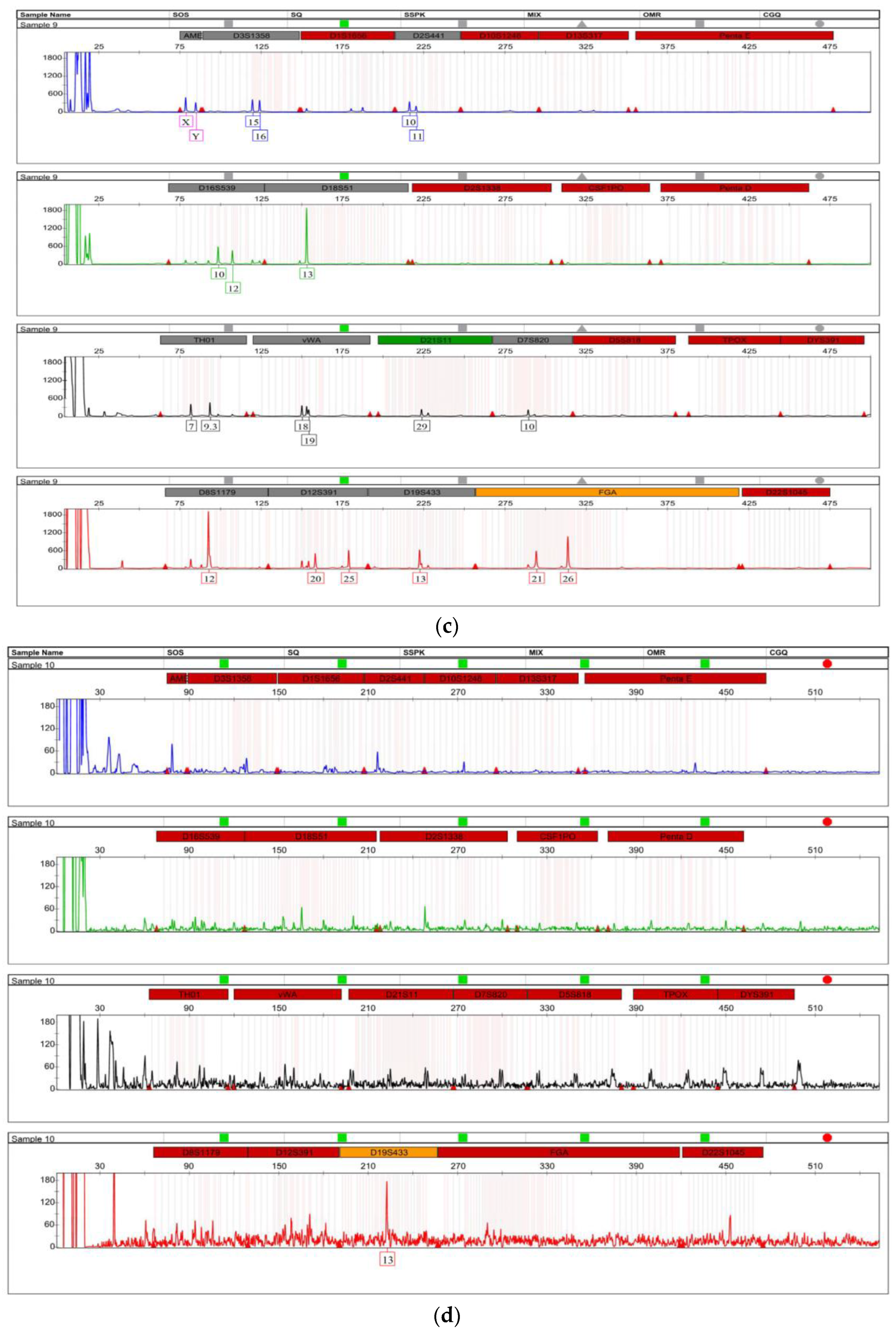
3.2. STR Profiling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Damelio, R.; Gardner, R.M. Bloodstain Pattern Analysis: With an Introduction to Crime Scene Reconstruction; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Virkler, K.; Lednev, I.K. Analysis of body fluids for forensic purposes: From laboratory testing to non-destructive rapid confirmatory identification at a crime scene. Forensic Sci. Int. 2009, 188, 1–17. [Google Scholar] [CrossRef]
- Curic, G.; Hercog, R.; Vrselja, Z.; Wagner, J. Identification of person and quantification of human DNA recovered from mosquitoes (Culicidae). Forensic Sci. Int. Genet. 2014, 8, 109–112. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Hansen, I.A.; Raikhel, A.S. Nutritional regulation of vitellogenesis in mosquitoes: Implications for anautogeny. Insect Biochem. Mol. Biol. 2005, 35, 661–675. [Google Scholar] [CrossRef] [PubMed]
- Clements, A.N. The Biology of Mosquitoes. Volume 1: Development, Nutrition and Reproduction; Cambridge University Press: Cambridge, UK, 1992; ISBN 987-184-53242-8. [Google Scholar]
- Lehane, M.J. The Biology of Blood-Sucking in Insects, 2nd ed.; Cambridge University Press: New York, NY, USA, 2005; p. 321. ISBN 978-0-521-83608-1. [Google Scholar]
- Chadee, D.D.; Beier, J.C. Factors influencing the duration of blood-feeding by laboratory-reared and wild Aedes aegypti (Diptera: Culicidae) from Trinidad, West Indies. Ann. Trop. Med. Parasitol. 1997, 91, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Edman, J.; Scott, T.; Costero, A.; Morrison, A.; Harrington, L.; Clark, G. Aedes aegypti (Diptera: Culicidae) movement influenced by availability of oviposition sites. J. Med. Entomol. 1998, 35, 578–583. [Google Scholar] [CrossRef]
- Rabêlo, K.; Albuquerque, C.; Tavares, V.; Santos, S.; Souza, C.; Oliveira, T.; Oliveira, N.; Crovella, S. Trace samples of human blood in mosquitoes as a forensic investigation tool. Genet. Mol. Res. 2015, 14, 14847–14856. [Google Scholar] [CrossRef] [PubMed]
- Trájer, A.J. Which mosquitoes (Diptera: Culicidae) are candidates for DNA extraction in forensic practice? J. Forensic Leg. Med. 2018, 58, 183–191. [Google Scholar] [CrossRef]
- Spitaleri, S.; Romano, C.; Di Luise, E.; Ginestra, E.; Saravo, L. Genotyping of human DNA recovered from mosquitoes found on a crime scene. Int. Congr. Ser. 2006, 1288, 574–576. [Google Scholar] [CrossRef]
- Rabêlo, K.; Albuquerque, C.; Tavares, V.; Santos, S.; Souza, C.; Oliveira, T.; Moura, R.; Brandão, L.; Crovella, S. Detecting multiple DNA human profile from a mosquito blood meal. Genet. Mol. Res. 2016, 15, gmr.15037547. [Google Scholar] [CrossRef]
- Scott, T.W.; Githeko, A.K.; Fleisher, A.; Harrington, L.C.; Yan, G. DNA profiling of human blood in anophelines from lowland and highland sites in western Kenya. Am. J. Trop. Med. Hyg. 2006, 75, 231–237. [Google Scholar] [CrossRef]
- Gray, S.L.; Tiedge, T.M.; Butkus, J.M.; Earp, T.J.; Lindner, S.E.; Roy, R. Determination of human identity from Anopheles stephensi mosquito blood meals using direct amplification and massively parallel sequencing. Forensic Sci. Int. Genet. 2020, 48, 102347. [Google Scholar] [CrossRef]
- Butler, J.M. Forensic DNA Typing: Biology, Technology, and Genetics of STR Markers, 2nd ed.; Elsevier Academic Press: London, UK, 2005; ISBN 0-12-147952-8. [Google Scholar]
- Hiroshige, Y.; Hara, M.; Nagai, A.; Hikitsuchi, T.; Umeda, M.; Kawajiri, Y.; Nakayama, K.; Suzuki, K.; Takada, A.; Ishii, A. A human genotyping trial to estimate the post-feeding time from mosquito blood meals. PLoS ONE 2017, 12, e0179319. [Google Scholar] [CrossRef]
- Kreike, J.; Kampfer, S. Isolation and characterization of human DNA from mosquitoes (Culicidae). Int. J. Leg. Med. 1999, 112, 380–382. [Google Scholar] [CrossRef]
- MacDougall, C. Effect of Blood Meal Size on Mosquito Response to Disturbance While Blood Feeding on a Simulated Host. Master’s Thesis, Department of Biological Sciences, Simon Fraser University, Bumaby, BC, Canada, 2005. [Google Scholar]
- Ibrahim, A.M. Impact of adult age on forensic use of Culex pipiens mosquito (Diptera: Culicidae). Int. J. Mosq. Res. 2015, 2, 28–32. [Google Scholar]
- Swaran, Y.C.; Welch, L. A comparison between direct PCR and extraction to generate DNA profiles from samples retrieved from various substrates. Forensic Sci. Int. Genet. 2012, 6, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Videvall, E.; Strandh, M.; Engelbrecht, A.; Cloete, S.; Cornwallis, C.K. Direct PCR offers a fast and reliable alternative to conventional DNA isolation methods for gut microbiomes. MSystems 2017, 2, e00132-17. [Google Scholar] [CrossRef] [PubMed]
- Van Oorschot, R.A.; Ballantyne, K.N.; Mitchell, R.J. Forensic trace DNA: A review. Investig. Genet. 2010, 1, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.M.; Maingon, R.; Taylor, P.; Hurd, H. The effects of infection with Plasmodium yoelii nigeriensis on the reproductive fitness of the mosquito Anopheles gambiae. Invertebr. Reprod. Dev. 1999, 36, 217–222. [Google Scholar] [CrossRef]
- List, I.S. SOP: Blood Collection in the Mouse, Intracardiac. Va. Tech LACUC Univ. Vet. Anim. Resour. 2017, 1, 1–3. [Google Scholar]
- Boix, F.; Andersen, J.M.; Mørland, J. Pharmacokinetic modeling of subcutaneous heroin and its metabolites in blood and brain of mice. Addict. Biol. 2013, 18, 1–7. [Google Scholar] [CrossRef]
- Baeshen, R.; Ekechukwu, N.E.; Toure, M.; Paton, D.; Coulibaly, M.; Traoré, S.F.; Tripet, F. Differential effects of inbreeding and selection on male reproductive phenotype associated with the colonization and laboratory maintenance of Anopheles gambiae. Malar. J. 2014, 13, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Briegel, H.; Lea, A.O. Influence of the endocrine system on tryptic activity in female Aedes aegypti. J. Insect Physiol. 1979, 25, 227–230. [Google Scholar] [CrossRef]
- Hopwood, J.A.; Ahmed, A.M.; Polwart, A.; Williams, G.T.; Hurd, H. Malaria-induced apoptosis in mosquito ovaries: A mechanism to control vector egg production. J. Exp. Biol. 2001, 204, 2773–2780. [Google Scholar] [CrossRef]
- Houseman, J.G.; Downe, A. Methods of measuring blood meal size and proteinase activity for determining effects of mated state on digestive processes of female Aedes aegypti (L.) (Diptera: Culicidae). Can. Entomol. 1986, 118, 241–248. [Google Scholar] [CrossRef]
- Konishi, E. Size of blood meals of Aedes albopictus and Culex tritaeniorhynchus (Diptera: Culicidae) feeding on an unrestrained dog infected with Dirofilaria immitis (Spirurida: Filariidae). J. Med. Entomol. 1989, 26, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Oostdik, K.; Lenz, K.; Nye, J.; Schelling, K.; Yet, D.; Bruski, S.; Strong, J.; Buchanan, C.; Sutton, J.; Linner, J. Developmental validation of the PowerPlex® Fusion System for analysis of casework and reference samples: A 24-locus multiplex for new database standards. Forensic Sci. Int. Genet. 2014, 12, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Butler, J.M.; Hill, C.R. Biology and genetics of new autosomal STR loci useful for forensic DNA analysis. Forensic Sci. Rev. 2012, 24, 15. [Google Scholar]
- Byrd, J.H.; Tomberlin, J.K. Forensic Entomology: The Utility of Arthropods in Legal Investigations, 3rd ed.; CRC Press: London, UK; New York, NY, USA, 2019; ISBN 978-0-815-35016-3. Available online: https://books.google.com.sa/books?id=HFG6qQeCY-8C&printsec=frontcover#v=onepage&q&f=false (accessed on 10 May 2023).
- Mumcuoglu, K.Y.; Gallili, N.; Reshef, A.; Brauner, P.; Grant, H. Use of human lice in forensic entomology. J. Med. Entomol. 2004, 41, 803–806. [Google Scholar] [CrossRef]
- Schal, C.; Czado, N.; Gamble, R.; Barrett, A.; Weathers, K.; Lodhi, K.M. Isolation, identification, and time course of human DNA typing from bed bugs, Cimex lectularius. Forensic Sci. Int. 2018, 293, 1–6. [Google Scholar] [CrossRef]
- Atoni, E.; Zhao, L.; Karungu, S.; Obanda, V.; Agwanda, B.; Xia, H.; Yuan, Z. The discovery and global distribution of novel mosquito-associated viruses in the last decade (2007–2017). Rev. Med. Virol. 2019, 29, e2079. [Google Scholar] [CrossRef]
- Greenberg, J.A.; DiMenna, M.A.; Hanelt, B.; Hofkin, B.V. Analysis of post-blood meal flight distances in mosquitoes utilizing zoo animal blood meals. J. Vector Ecol. 2012, 37, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Norris, L.C.; Fornadel, C.M.; Hung, W.-C.; Pineda, F.J.; Norris, D.E. Frequency of multiple blood meals taken in a single gonotrophic cycle by Anopheles arabiensis mosquitoes in Macha, Zambia. Am. J. Trop. Med. Hyg. 2010, 83, 33. [Google Scholar] [CrossRef] [PubMed]
- Demeke, T.; Eng, M.; Holigroski, M.; Lee, S.-J. Effect of amount of DNA on Digital PCR assessment of genetically engineered canola and soybean events. Food Anal. Methods 2021, 14, 372–379. [Google Scholar] [CrossRef]
- Budowle, B.; Moretti, T.R.; Niezgoda, S.J.; Brown, B.L. CODIS and PCR-based short tandem repeat loci: Law enforcement tools. In Second European Symposium on Human Identification; Promega Corporation: Madison, WI, USA, 1998; Volume 7388. [Google Scholar]
- Martínez-de la Puente, J.; Ruiz, S.; Soriguer, R.; Figuerola, J. Effect of blood meal digestion and DNA extraction protocol on the success of blood meal source determination in the malaria vector Anopheles atroparvus. Malar. J. 2013, 12, 109. [Google Scholar] [CrossRef] [PubMed]
- Steffen, C.R.; Coble, M.D.; Gettings, K.B.; Vallone, P.M. Corrigendum to ‘US population data for 29 autosomal STR loci. Forensic Sci. Int. Genet. 2017, 31, e36–e40. [Google Scholar] [CrossRef] [PubMed]
- Oshaghi, M.A.; Chavshin, A.R.; Vatandoost, H. Analysis of mosquito bloodmeals using RFLP markers. Exp. Parasitol. 2006, 114, 259–264. [Google Scholar] [CrossRef]
- Ibrahim, Y.; Hussain, S.M.; Alnasser, S.; Almohandes, H.; Sarhandi, I. Patterns and sociodemographic characteristics of substance abuse in Al Qassim, Saudi Arabia: A retrospective study at a psychiatric rehabilitation center. Ann. Saudi Med. 2018, 38, 319–325. [Google Scholar] [CrossRef]
- Brouwer, R. Variations in human body odour as a cause of individual differences of attraction for malaria mosquitoes. Trop. Geogr. Med. 1960, 12, 186–192. [Google Scholar]
- Brady, J.; Costantini, C.; Sagnon, N.; Gibson, G.; Coluzzi, M. The role of body odours in the relative attractiveness of different men to malarial vectors in Burkina Faso. Ann. Trop. Med. Parasitol. 1997, 91, S121–S122. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Hours Post-Feeding | Mosquito Groups (Based on the Source of Blood Meals) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G1 | G2 | G3 | G4 | G5 | G6 | |||||||
| C | CT | C | CT | C | CT | C | CT | C | CT | C | CT | |
| 0 | 1.252 ± 0.001 | 29.91 | 1.305 ± 0.001 | 29.03 | 1.736 ± 0.001 | 28.08 | 1.298 ± 0.0005 | 29.69 | 1.255 ± 0.005 | 29.87 | 1.042 ± 0.004 | 31.36 |
| 2 | 0.661 ± 0.001 | 32.28 | 0.959 ± 0.01 | 31.98 | 0.963 ± 0.002 | 31.96 | 0.574 ± 0.005 | 32.34 | 0.545 ± 0.006 | 32.38 | 0.44 ± 0.011 | 32.51 |
| 4 | 0.656 ± 0.002 | 32.29 | 0.604 ± 0.002 | 32.28 | 0.623 ± 0.006 | 32.28 | 0.229 ± 0.001 | 33.77 | 0.211 ± 0.0005 | 33.79 | 0.192 ± 0.0005 | 33.85 |
| 6 | 0.486 ± 0.006 | 32.31 | 0.288 ± 0.005 | 33.46 | 0.457 ± 0.002 | 32.49 | 0.218 ± 0.001 | 33.76 | 0.19 ± 0.005 | 33.83 | 0.092 ± 0.001 | 35.03 |
| 8 | 0.217 ± 0.001 | 33.78 | 0.278 ± 0.001 | 33.49 | 0.309 ± 0.001 | 33.43 | 0.197 ± 0.002 | 33.84 | 0.161 ± 0.005 | 33.92 | 0.085 ± 0.003 | 35.24 |
| 10 | 0.145 ± 0.005 | 33.98 | 0.189 ± 0.005 | 33.85 | 0.211 ± 0.006 | 33.79 | 0.134 ± 0.003 | 33.99 | 0.111 ± 0.006 | 34.02 | 0.063 ± 0.001 | 35.12 |
| 12 | 0.129 ± 0.0005 | 33.10 | 0.147 ± 0.001 | 33.99 | 0.154 ± 0.005 | 33.97 | 0.098 ± 0.0005 | 35.08 | 0.077 ± 0.001 | 35.11 | 0.056 ± 0.001 | 35.13 |
| 24 | 0.094 ± 0.001 | 35.10 | 0.112 ± 0.001 | 34.02 | 0.118 ± 0.001 | 33.87 | 0.052 ± 0.001 | 35.15 | 0.039 ± 0.0005 | 35.25 | 0.019 ± 0.0005 | 35.45 |
| 36 | 0.05 ± 0.005 | 35.19 | 0.094 ± 0.002 | 35.11 | 0.091 ± 0.0005 | 35.14 | 0.086 ± 0.0005 | 35.14 | 0.003 ± 0.0005 | 38.88 | 0.001 ± 0.00 | 44.18 |
| 48 | 0.004 ± 0.0005 | 38.86 | 0.007 ± 0.001 | 38.84 | 0.008 ± 0.0005 | 38.83 | 0.007 ± 0.0005 | 38.84 | 0.002 ± 0.00 | 39.16 | 0.001 ± 0.00 | 44.21 |
| Control | 144.4 * ± 0.34 | 22.1 | 241.8 * ± 1.03 | 21.8 | 288.2 * ± 0.69 | 21.3 | 237.99 * ± 1.14 | 21.86 | 0.001 * ± 0.00 | 44.21 | 145.1 * ± 0.6 | 22.14 |
| STR Loci | Hours Post-Blood Feeding | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 2 h | 4 h | 6 h | 8 h | 10 h | 12 h | 24 h | 36 h | 48 h | Control | |
| Amelogenin | XY | XY | XY | XY | XY | XY | XY | N/A | N/A | N/A | XY |
| D3S1358 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | N/A | N/A | 15,16 |
| D1S1656 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | N/A | N/A | 15,16.3 |
| D2S441 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | 10,11 |
| D10S1248 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | N/A | N/A | N/A | 14,15 |
| D13S317 | 11,13 | 11,13 | 11,13 | 11,13 | 11,13 | 11,13 | 11,13 | N/A | N/A | N/A | 11,13 |
| Penta E | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | N/A | N/A | N/A | 16,18 |
| D16S539 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | N/A | N/A | 10,12 |
| D18S51 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | N/A | 13,13 |
| D2S1338 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | N/A | N/A | N/A | 17,18 |
| CSF1PO | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | N/A | 10,11 |
| Penta D | 9,15 | 9,15 | 9,15 | 9,15 | 9,15 | 9,15 | 9,15 | N/A | N/A | N/A | 9,15 |
| TH01 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | N/A | N/A | 7,9.3 |
| vWA | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | N/A | N/A | 18,19 |
| D21S11 | 29,30 | 29,30 | 29,30 | 29,30 | 29,30 | 29,30 | 29,30 | N/A | N/A | N/A | 29,30 |
| D7S820 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | N/A | 10,11 |
| D5S818 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | N/A | N/A | N/A | 12,13 |
| TPOX | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | N/A | N/A | N/A | 11,11 |
| DYS391 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | N/A | N/A | 11,11 |
| D8S1179 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | N/A | 12,12 |
| D12S391 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | N/A | N/A | 20,25 |
| D19S433 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | N/A | N/A | 13,13 |
| FGA | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | N/A | 21,26 | N/A | 21,26 |
| D22S1045 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | N/A | N/A | N/A | 15,16 |
| STR Loci | Hours Post-Blood Feeding | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 24 | 36 | 48 | Control | |
| Amelogenin | XX | XX | XX | XX | XX | XX | XX | XX | XX | XX | XX |
| D3S1358 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 |
| D1S1656 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 |
| D2S441 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 |
| D10S1248 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 |
| D13S317 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 |
| Penta E | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 |
| D16S539 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 |
| D18S51 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 |
| D2S1338 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 |
| CSF1PO | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | 10,11 |
| Penta D | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 |
| TH01 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 |
| vWA | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 |
| D21S11 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 |
| D7S820 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 |
| D5S818 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 |
| TPOX | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 |
| D8S1179 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 |
| D12S391 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 |
| D19S433 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 |
| FGA | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 |
| D22S1045 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 |
| STR Loci | Hours Post-Blood Feeding | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 2 h | 4 h | 6 h | 8 h | 10 h | 12 h | 24 h | 36 h | 48 h | Control | |
| Amelogenin | XX,Y | XX,Y | XXY | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | N/A | XX,Y |
| D3S1358 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | N/A | 15,16,17 |
| D1S1656 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3, 16.3 | N/A | 15,15.3,16.3 |
| D2S441 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | N/A | 10,11,14 |
| D10S1248 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | N/A | 13,14,15 |
| D13S317 | 11,12,13,14 | 11,12,13,14 | 11,12,13,14 | 11,12,13,14 | 11,12,13,14 | 11,12,13,14 | 11,12,13,14 | 11,12,13,14 | 11,12,13,14 | N/A | 11,12,13,14 |
| Penta E | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | N/A | 16,18 |
| D16S539 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | N/A | 9,10,12 |
| D18S51 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | N/A | 13,16 |
| D2S1338 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | N/A | 17,18 |
| CSF1PO | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | 10,11 |
| Penta D | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | N/A | 9,15,16 |
| TH01 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 9,15,16 | N/A | 6,7,9.3 |
| vWA | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | N/A | N/A | 18,19 |
| D21S11 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 18,19 | N/A | 29,30,32.2 |
| D7S820 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | 10,11 |
| D5S818 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | N/A | 12,13,14 |
| TPOX | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | N/A | 9,11 |
| DYS391 | 7,11 | 7,11 | 7,11 | 7,11 | 7,11 | 7,11 | 7,11 | 7,11 | 7, N/A | N/A | 7,11 |
| D8S1179 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | N/A | 12,14,15 |
| D12S391 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | N/A | 20,23,25 |
| D19S433 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | N/A | 12,13 |
| FGA | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | N/A | 21,23,26 |
| D22S1045 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | N/A | 15,16 |
| STR Loci | Hours Post-Blood Feeding | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 h | 2 h | 4 h | 6 h | 8 h | 10 h | 12 h | 24 h | 36 h | 48 h | Control | |
| Amelogenin | XY | XY | XY | XY | XY | XY | XY | XY | N/A | N/A | XY |
| D3S1358 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | N/A | N/A | 15,16 |
| D1S1656 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | 15,16.3 | N/A | N/A | 15,16.3 |
| D2S441 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | 10,11 |
| D10S1248 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | N/A | N/A | 14,15 |
| D13S317 | 11,13 | 11,13 | 11,13 | 11,13 | 11,13 | 11,13 | 11,13 | 11,13 | N/A | N/A | 11,13 |
| Penta E | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | N/A | N/A | 16,18 |
| D16S539 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | 10,12 | N/A | N/A | 10,12 |
| D18S51 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 |
| D2S1338 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | N/A | N/A | 17,18 |
| CSF1PO | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | 10,11 |
| Penta D | 9,15 | 9,15 | 9,15 | 9,15 | 9,15 | 9,15 | 9,15 | 9,15 | N/A | N/A | 9,15 |
| TH01 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | 7,9.3 | N/A | N/A | 7,9.3 |
| vWA | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | N/A | N/A | 18,19 |
| D21S11 | 29,30 | 29,30 | 29,30 | 29,30 | 29,30 | 29,30 | 29,30 | 29,30 | N/A | N/A | 29,30 |
| D7S820 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | 10,11 |
| D5S818 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | N/A | N/A | 12,13 |
| TPOX | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | N/A | N/A | 11,11 |
| DYS391 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | N/A | N/A | 11,11 |
| D8S1179 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 | 12,12 |
| D12S391 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 | 20,25 |
| D19S433 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 | 13,13 |
| FGA | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 | 21,26 |
| D22S1045 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | N/A | N/A | 16,16 |
| STR Loci | Hours Post-Blood Feeding | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 24 | 36 | 48 | Control | |
| Amelogenin | XX | XX | XX | XX | XX | XX | XX | XX | N/A | N/A | XX |
| D3S1358 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | 17,17 | N/A | N/A | 17,17 |
| D1S1656 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | 15.3,16.3 | N/A | N/A | 15.3,16.3 |
| D2S441 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | 10,14 | N/A | N/A | 10,14 |
| D10S1248 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | 13,14 | N/A | N/A | 13,14 |
| D13S317 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | N/A | N/A | 12,14 |
| Penta E | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | N/A | N/A | N/A | 16,18 |
| D16S539 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | 9,10 | N/A | N/A | 9,10 |
| D18S51 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | N/A | N/A | 13,16 |
| D2S1338 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | N/A | N/A | 17,18 |
| CSF1PO | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | 10,11 |
| Penta D | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9,16 | 9, N/A | N/A | N/A | 9,16 |
| TH01 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | 6,7 | N/A | N/A | 6,7 |
| vWA | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | N/A | N/A | 18,19 |
| D21S11 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | 30,32.2 | N/A | N/A | 30,32.2 |
| D7S820 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | 11,11 | N/A | N/A | 11,11 |
| D5S818 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | 12,14 | N/A | N/A | 12,14 |
| TPOX | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | N/A | N/A | 9,11 |
| D8S1179 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | 14,15 | N/A | N/A | 14,15 |
| D12S391 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | 23,23 | N/A | N/A | 23,23 |
| D19S433 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12, N/A | N/A | 12,13 |
| FGA | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 21,23 | 23, N/A | N/A | 21,23 |
| D22S1045 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | N/A | N/A | N/A | 15,16 |
| STR Loci | Hours Post-Blood Feeding | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 0 | 2 | 4 | 6 | 8 | 10 | 12 | 24 | 36 | 48 | Control | |
| Amelogenin | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y | XX,Y |
| D3S1358 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | 15,16,17 | N/A | 15,16,17 |
| D1S1656 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15.3,16.3 | 15,15,16.3 | N/A | 15,15.3,16.3 |
| D2S441 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | 10,11,14 | N/A | 10,11,14 |
| D10S1248 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | 13,14,15 | N/A | 13,14,15 |
| D13S317 | 11,12,13,14 | 11,12,13, 14 | 11,12,13, 14 | 11,12,13, 14 | 11,12,13, 14 | 11,12,13, 14 | 11,12,13, 14 | 11,12,13, 14 | 11,12,13,14 | N/A | 11,12,13, 14 |
| Penta E | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | 16,18 | N/A | N/A | 16,18 |
| D16S539 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | 9,10,12 | N/A | N/A | 9,10,12 |
| D18S51 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | 13,16 | N/A | N/A | 13,16 |
| D2S1338 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | 17,18 | N/A | N/A | 17,18 |
| CSF1PO | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | 10,11 |
| Penta D | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15,16 | 9,15, N/A | N/A | N/A | 9,15,16 |
| TH01 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | 6,7,9.3 | N/A | N/A | 6,7,9.3 |
| vWA | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | 18,19 | N/A | N/A | 18,19 |
| D21S11 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | 29,30,32.2 | N/A | N/A | 29,30,32.2 |
| D7S820 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | 10,11 | N/A | N/A | 10,11 |
| D5S818 | 11,12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | 12,13,14 | N/A | N/A | 12,13,14 |
| TPOX | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | 9,11 | N/A | N/A | 9,11 |
| DYS391 | 7,11 | 7,11 | 7,11 | 7,11 | 7,11 | 7,11 | 7,11 | 7, N/A | N/A | N/A | 7,11 |
| D8S1179 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | 12,14,15 | N/A | N/A | 12,14,15 |
| D12S391 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | 20,23,25 | N/A | N/A | 20,23,25 |
| D19S433 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | 12,13 | N/A | N/A | 12,13 |
| FGA | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | 21,23,26 | N/A | N/A | 21,23,26 |
| D22S1045 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | 15,16 | N/A | N/A | 15,16 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, A.M.; Alotaibi, A.M.; Al-Qahtani, W.S.; Tripet, F.; Amer, S.A. Forensic DNA Analysis of Mixed Mosquito Blood Meals: STR Profiling for Human Identification. Insects 2023, 14, 467. https://doi.org/10.3390/insects14050467
Ahmed AM, Alotaibi AM, Al-Qahtani WS, Tripet F, Amer SA. Forensic DNA Analysis of Mixed Mosquito Blood Meals: STR Profiling for Human Identification. Insects. 2023; 14(5):467. https://doi.org/10.3390/insects14050467
Chicago/Turabian StyleAhmed, Ashraf Mohamed, Amani Mohammed Alotaibi, Wedad Saeed Al-Qahtani, Frederic Tripet, and Sayed Amin Amer. 2023. "Forensic DNA Analysis of Mixed Mosquito Blood Meals: STR Profiling for Human Identification" Insects 14, no. 5: 467. https://doi.org/10.3390/insects14050467