
Research on Photoinduction-Based Technology for Trapping Asian Longhorned Beetle (Anoplophora glabripennis (Motschulsky, 1853) (Coleoptera: Cerambycidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
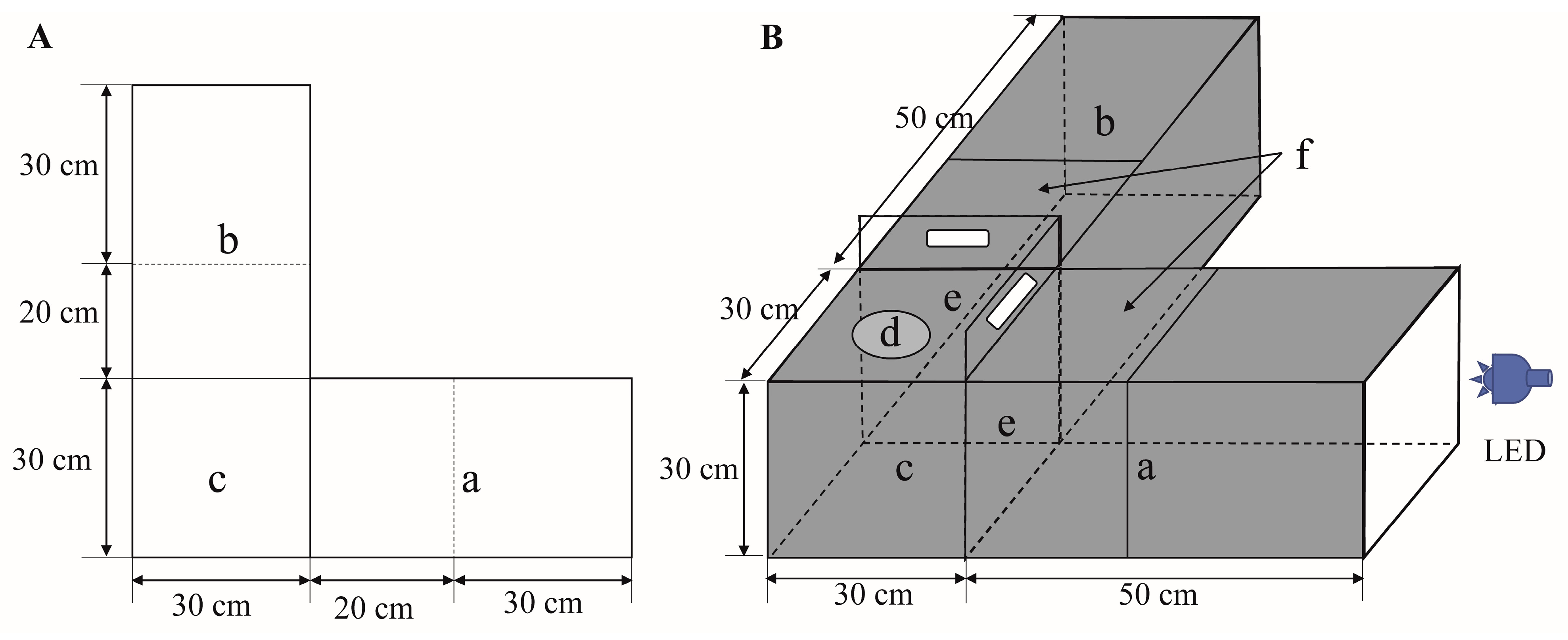
2.2. Experimental Apparatus and Light Sources
2.3. Behavioral Experiments
2.3.1. Effect of Exposure Time on the Phototactic Behavioral Responses
2.3.2. Effect of Diel Rhythm on the Phototactic Behavioral Responses
2.3.3. Monochromatic LED Light Preference
2.3.4. Effect of Light Intensities on the Phototactic Behavioral Responses
2.4. Statistical Analysis
3. Results
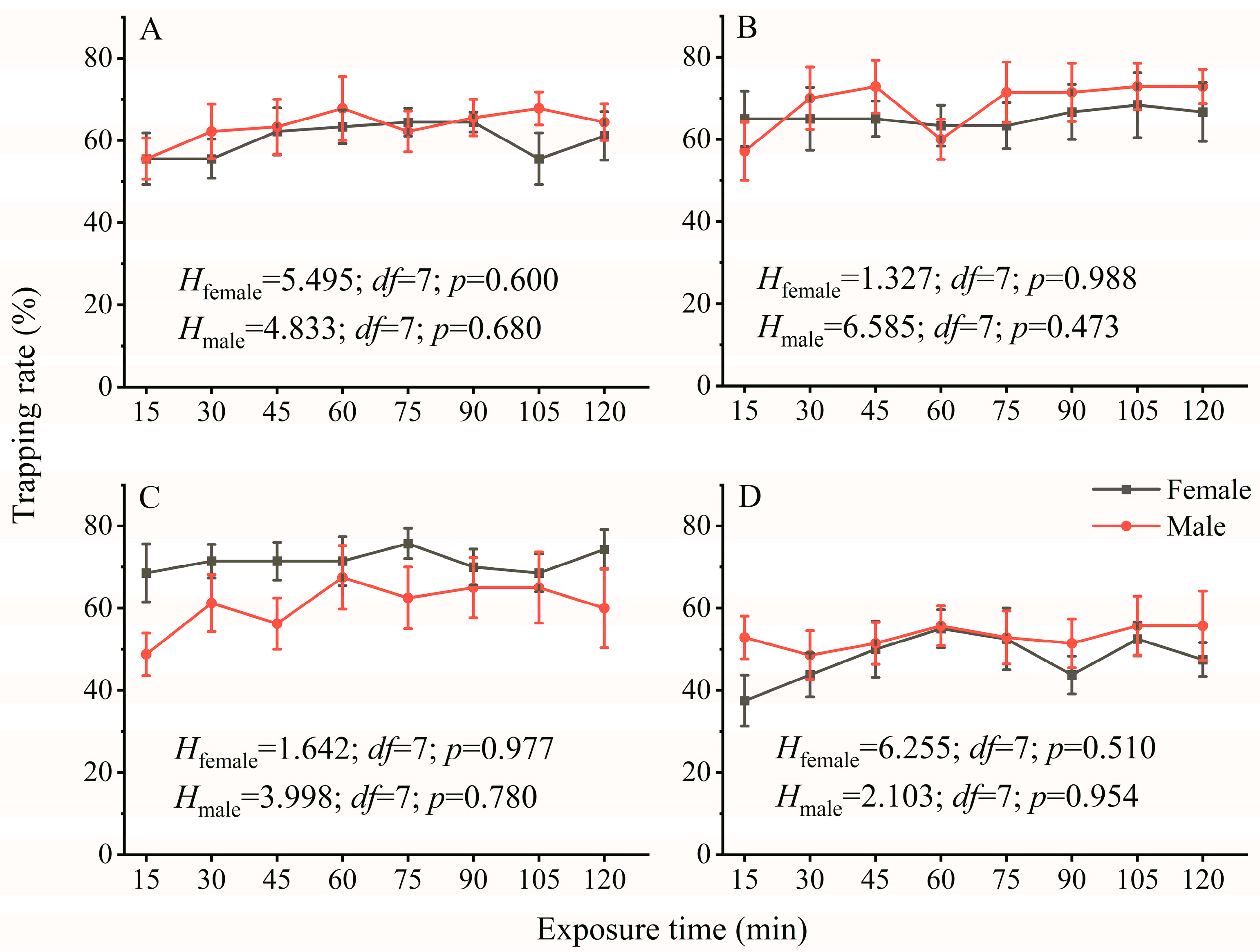
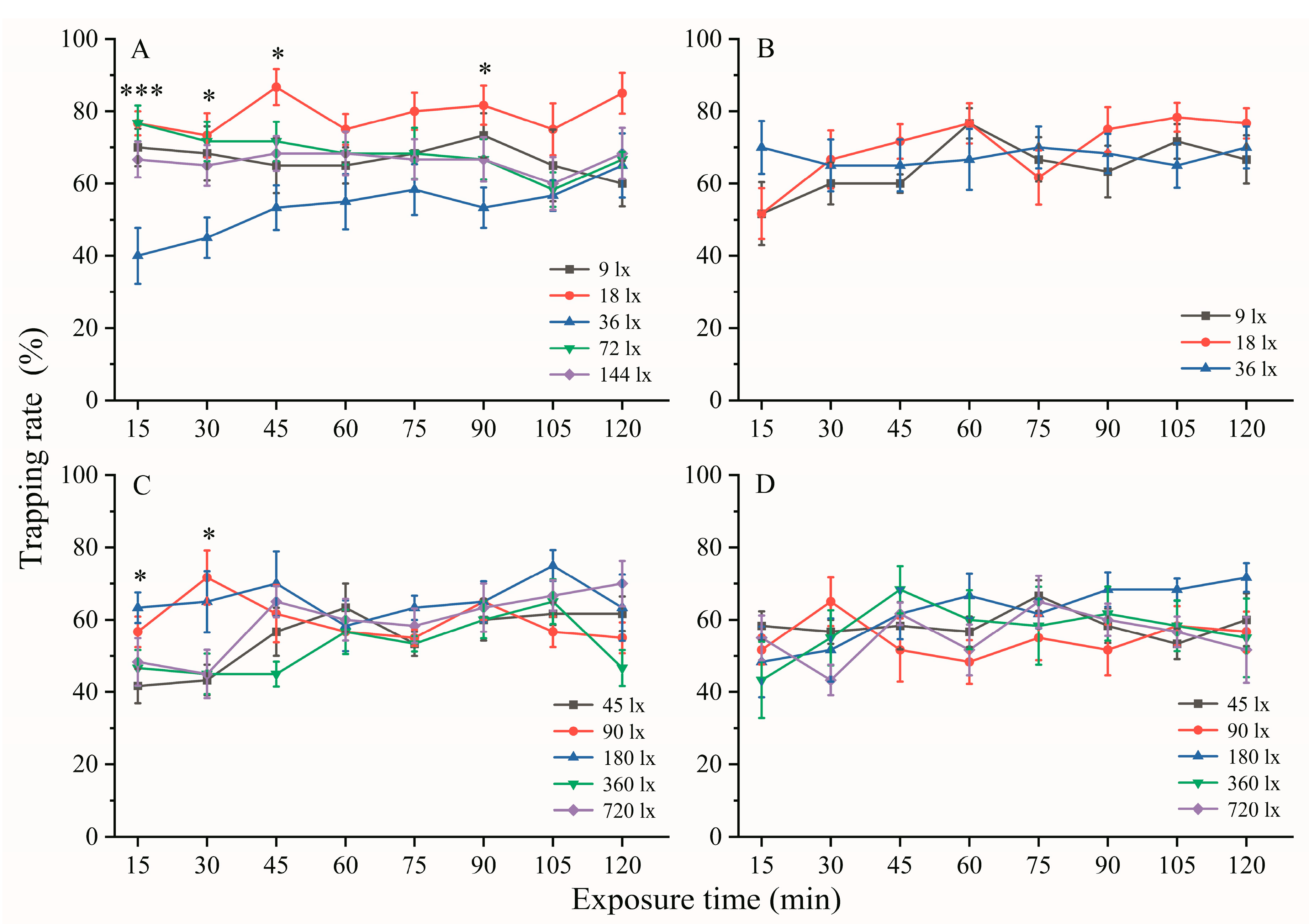
3.1. Effect of Exposure Time on the Phototactic Behavioral Response
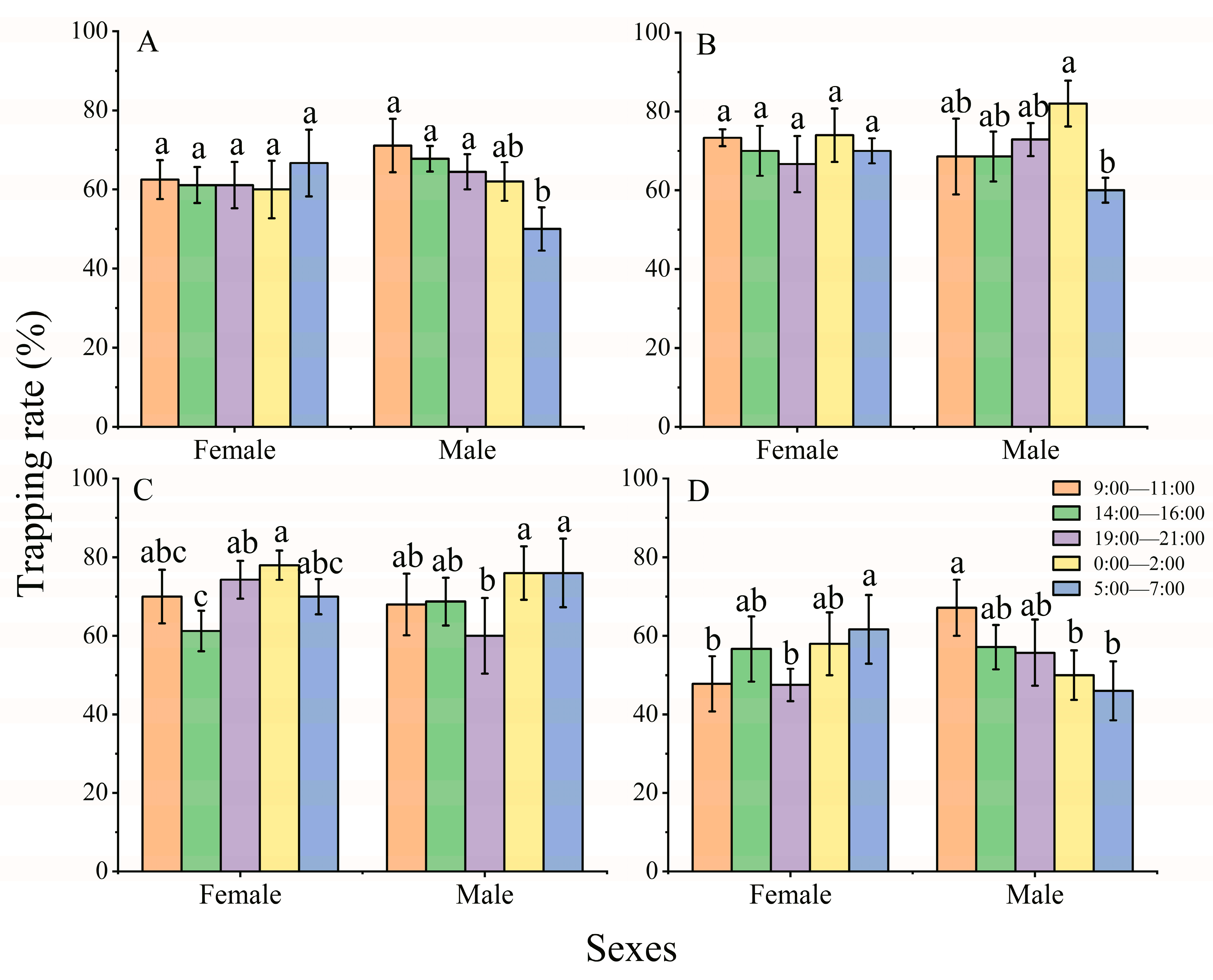
3.2. Effect of Diel Rhythm on the Phototactic Behavioral Response
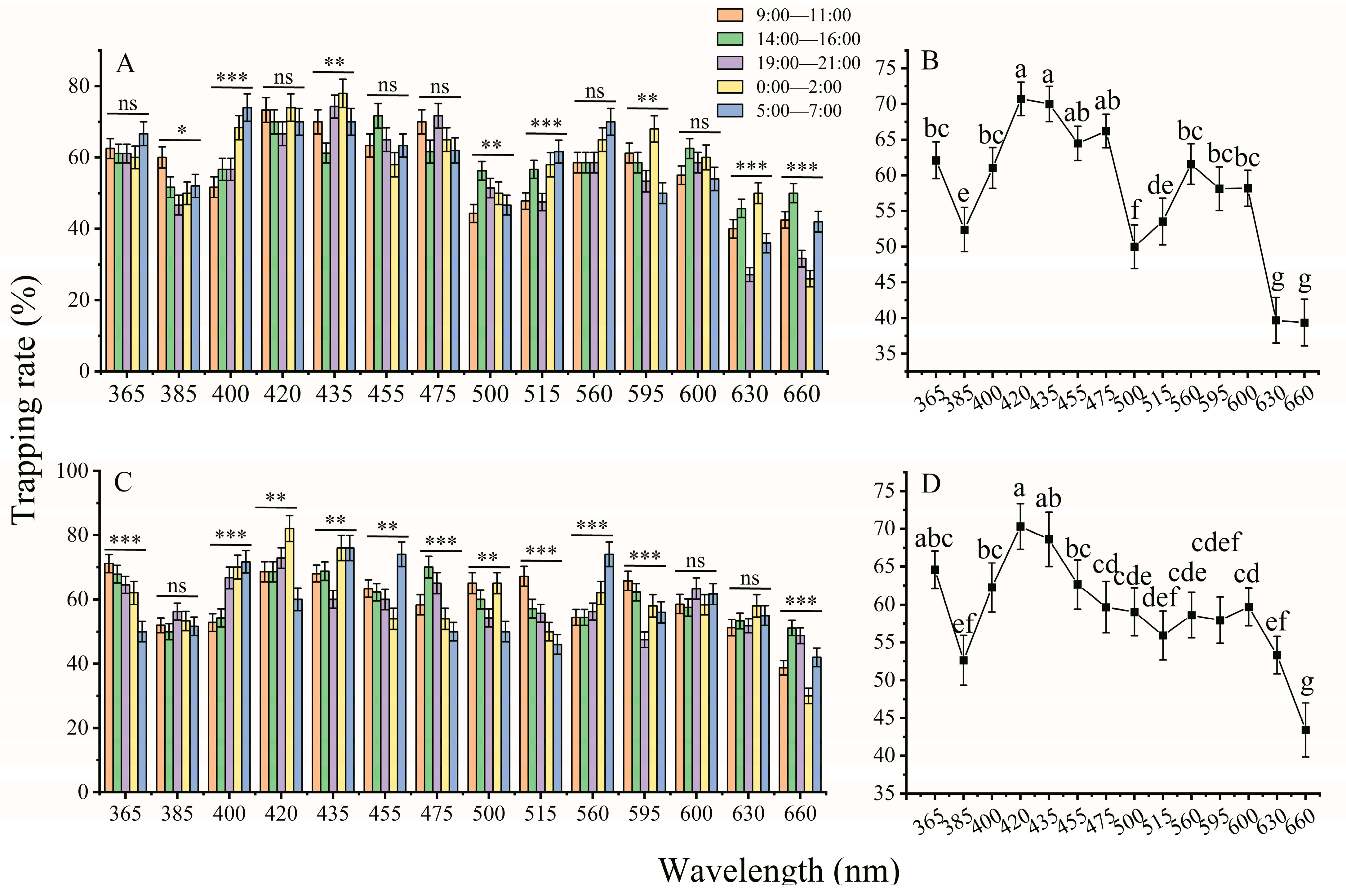
3.3. Monochromatic LED Light Preference
3.4. Effect of Light Intensities on the Phototactic Behavioral Response
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scherm, H.; Sutherst, R.W.; Harrington, R.; Ingram, J.S.I. Global networking for assessment of impacts of global change on plant pests. Environ. Pollut. 2000, 108, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Beketov, M.A.; Kefford, B.J.; Schafer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [PubMed]
- Basset, Y.; Lamarre, G. Toward a world that values insects. Science 2019, 364, 1230–1231. [Google Scholar] [CrossRef]
- Petrovskii, S.; Petrovskaya, N.; Bearup, D. Multiscale approach to pest insect monitoring: Random walks, pattern formation, synchronization, and networks. Phys. Life Rev. 2014, 11, 467–525. [Google Scholar] [CrossRef] [PubMed]
- Nieri, R.; Anfora, G.; Mazzoni, V.; Stacconi, M. Semiochemicals, semiophysicals and their integration for the development of innovative multi-modal systems for agricultural pests’ monitoring and control. Entomol. Gen. 2022, 42, 167–183. [Google Scholar] [CrossRef]
- Branco, S.; Faccoli, M.; Brockerhoff, E.G.; Roux, G.; Jactel, H.; Desneux, N.; Gachet, E.; Mouttet, R.; Streito, J.C.; Branco, M. Preventing invasions of Asian longhorn beetle and Citrus longhorn beetle: Are we on the right track? J. Pest Sci. 2022, 95, 41–66. [Google Scholar] [CrossRef]
- Haack, R.A.; Herard, F.; Sun, J.; Turgeon, J.J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu. Rev. Entomol. 2010, 55, 521–546. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lyu, F.; Bi, Y.; Wang, Z. Reviews on the serious wood-boring pest Anoplophora glabripennis in forestry of China. Forest. Ecol. Sci. 2020, 35, 1–9. [Google Scholar]
- Xu, T.; Teale, S.A. Chemical ecology of the Asian longhorn beetle, Anoplophora glabripennis. J. Chem. Ecol. 2021, 47, 489–503. [Google Scholar] [CrossRef]
- Nehme, M.E.; Keena, M.A.; Zhang, A.; Baker, T.C.; Xu, Z.; Hoover, K. Evaluating the use of male-produced pheromone components and plant volatiles in two trap designs to monitor Anoplophora glabripennis. Environ. Entomol. 2010, 39, 169–176. [Google Scholar] [CrossRef]
- Xu, T.; Hansen, L.; Cha, D.H.; Hao, D.; Zhang, L.; Teale, S.A. Identification of a female-produced pheromone in a destructive invasive species: Asian longhorn beetle, Anoplophora glabripennis. J. Pest Sci. 2020, 93, 1321–1332. [Google Scholar] [CrossRef]
- Marchioro, M.; Faccoli, M. Successful eradication of the Asian longhorn beetle, Anoplophora glabripennis, from North-Eastern Italy: Protocol, techniques and results. Insects 2021, 12, 877. [Google Scholar] [CrossRef] [PubMed]
- Eyre, D.; Barbrook, J. The eradication of Asian longhorned beetle at Paddock Wood, UK. CABI Agric. Biosci. 2021, 2, 1–17. [Google Scholar] [CrossRef]
- Meng, P.S.; Trotter, R.T.; Keena, M.A.; Baker, T.C.; Yan, S.; Schwartzberg, E.G.; Hoover, K. Effects of pheromone and plant volatile release rates and ratios on trapping Anoplophora glabripennis (Coleoptera: Cerambycidae) in China. Environ. Entomol. 2014, 43, 1379–1388. [Google Scholar] [CrossRef]
- Wickham, J.D.; Xu, Z.; Teale, S.A. Evidence for a female-produced, long range pheromone of Anoplophora glabripennis (Coleoptera: Cerambycidae). Insect Sci. 2012, 19, 355–371. [Google Scholar] [CrossRef]
- Lyu, F.; Hai, X.; Wang, Z. Green-colored paperboard enhances the Asian longhorned beetle response to host plant odor cues. J. Pest Sci. 2021, 94, 1345–1355. [Google Scholar] [CrossRef]
- Li, D.; Tokoro, M.; Nacashima, T. Mechanism of mating action of Anoplophora glabripennis (Motsch.). J. Beijing Forest. Univ. 1999, 21, 33–36. [Google Scholar]
- Lyu, F.; Hai, X.; Wang, Z.; Yan, A.; Liu, B.; Bi, Y. Integration of visual and olfactory cues in host plant identification by the Asian Longhorned Beetle, Anoplophora glabripennis (Motschulsky) (Coleoptera: Cerambycidae). PLoS ONE 2015, 10, e0142752. [Google Scholar] [CrossRef]
- Kim, K.; Huang, Q.; Lei, C. Advances in insect phototaxis and application to pest management: A review. Pest Manag. Sci. 2019, 75, 3135–3143. [Google Scholar] [CrossRef]
- Lyu, F.; Hai, X.; Wang, Z.; Yan, A.; Bi, Y.; Liu, B. Diurnal rhythm of four types of adult behaviors of Anoplophora glabripennis. J. Northeast Forest. Univ. 2015, 43, 90–95. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, S.; Shen, Z.; Li, Z.; Zhang, X.; Liu, X.; Zhang, Q.; Liu, X. The expression of three opsin genes and phototactic behavior of Spodoptera exigua (Lepidoptera: Noctuidae): Evidence for visual function of opsin in phototaxis. Insect Biochem. Mol. Biol. 2018, 96, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Van der Kooi, C.J.; Stavenga, D.G.; Arikawa, K.; Belusic, G.; Kelber, A. Evolution of insect color vision: From spectral sensitivity to visual ecology. Annu. Rev. Entomol. 2021, 66, 435–461. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Song, H.; Li, C.; Huang, Q.; Lei, C. Effect of several factors on the phototactic response of the oriental armyworm, Mythimna separata (Lepidoptera: Noctuidae). J. Asia-Pac. Entomol. 2018, 21, 952–957. [Google Scholar] [CrossRef]
- Marchioro, M.; Battisti, A.; Faccoli, M. Light traps in shipping containers: A new tool for the early detection of insect alien species. J. Econ. Entomol. 2020, 113, 1718–1724. [Google Scholar] [CrossRef]
- Werle, C.T.; Bray, A.M.; Oliver, J.B.; Blythe, E.K.; Sampson, B.J. Ambrosia beetle (Coleoptera: Curculionidae: Scolytinae) captures using colored traps in Southeast Tennessee and South Mississippi. J. Entomol. Sci. 2014, 49, 373–382. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, H.S. Phototactic behavioral response of agricultural insects and stored-product insects to light-emitting diodes (LEDs). Appl. Biol. Chem. 2017, 60, 137–144. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Magnani, G.; Rassati, D. Effect of trap color on captures of bark-and wood-Boring Beetles (Coleoptera; Buprestidae and Scolytinae) and associated predators. Insects 2020, 11, 749. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Giannone, F.; Moino, S.; Rassati, D. Exploiting trap color to improve surveys of longhorn beetles. J. Pest Sci. 2021, 94, 871–883. [Google Scholar] [CrossRef]
- Wakakuwa, M.; Stewart, F.; Matsumoto, Y.; Matsunaga, S.; Arikawa, K. Physiological basis of phototaxis to near-infrared light in Nephotettix cincticeps. J. Comp. Physiol. A Neuroethol Sens. Neural. Behav. Physiol. 2014, 200, 527–536. [Google Scholar] [CrossRef]
- Chen, S.P.; Liu, Z.X.; Chen, Y.T.; Wang, Y.; Chen, J.Z.; Fu, S.; Ma, W.F.; Xia, S.; Liu, D.; Wu, T.; et al. CRISPR/Cas9-mediated knockout of LW-opsin reduces the efficiency of phototaxis in the diamondback moth Plutella xylostella. Pest Manag. Sci. 2021, 77, 3519–3528. [Google Scholar] [CrossRef]
- Li, C.; Tian, F.; Lin, T.; Wang, Z.; Liu, J.; Zeng, X. The expression and function of opsin genes related to the phototactic behavior of Asian citrus psyllid. Pest Manag. Sci. 2020, 76, 1578–1587. [Google Scholar] [CrossRef] [PubMed]
- Sander, S.E.; Hall, D.W. Variation in opsin genes correlates with signalling ecology in North American fireflies. Mol. Ecol. 2015, 24, 4679–4696. [Google Scholar] [CrossRef] [PubMed]
- Lord, N.P.; Plimpton, R.L.; Sharkey, C.R.; Suvorov, A.; Lelito, J.P.; Willardson, B.M.; Bybee, S.M. A cure for the blues: Opsin duplication and subfunctionalization for short-wavelength sensitivity in jewel beetles (Coleoptera: Buprestidae). BMC Evol. Biol. 2016, 16, 107. [Google Scholar] [CrossRef]
- Jackowska, M.; Bao, R.; Liu, Z.; McDonald, E.C.; Cook, T.A.; Friedrich, M. Genomic and gene regulatory signatures of cryptozoic adaptation: Loss of blue sensitive photoreceptors through expansion of long wavelength-opsin expression in the red flour beetle Tribolium castaneum. Front. Zool. 2007, 4, 24. [Google Scholar] [CrossRef]
- Sharkey, C.R.; Fujimoto, M.S.; Lord, N.P.; Shin, S.; McKenna, D.D.; Suvorov, A.; Martin, G.J.; Bybee, S.M. Overcoming the loss of blue sensitivity through opsin duplication in the largest animal group, beetles. Sci. Rep. 2017, 7, 8. [Google Scholar] [CrossRef] [PubMed]
- Pawson, S.M.; Watt, M.S.; Brockerhoff, E.G. Using differential responses to light spectra as a monitoring and control tool for Arhopalus ferus (Coleoptera: Cerambycidae) and other exotic wood-boring pests. J. Econ. Entomol. 2009, 102, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.C.; Turnbow, R.H. Cerambycidae new to Andros Island, Bahamas (Coleoptera). Coleopteris. Bull. 2007, 61, 581–588. [Google Scholar] [CrossRef]
- Lingafelter, S.W.; Horner, V.L. The Cerambycidae of North-central Texas. Coleopteris. Bull. 1993, 47, 159–191. [Google Scholar]
- Wang, J.; Che, S.; Qiu, L.; Li, G.; Shao, J.; Zhong, L.; Zhang, G.; Xu, H. Efficacy of emamectin benzoate trunk injection against the Asian long-horned beetle [Anoplophora glabripennis (Coleoptera: Cerambycidae)]. J. Econ. Entomol. 2020, 113, 340–347. [Google Scholar] [CrossRef]
- Wang, B.D.; Gao, R.T.; Mastro, V.C.; Reardon, R.C. Toxicity of four systemic neonicotinoids to adults of Anoplophora glabripennis (Coleoptera: Cerambycidae). J. Econ. Entomol. 2005, 98, 2292–2300. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.Y.; Kenis, M.; Cao, L.M.; Hoelmer, K.A. Exploring the potential for novel associations of generalist parasitoids for biological control of invasive woodboring beetles. BioControl 2021, 66, 97–112. [Google Scholar] [CrossRef]
- Wang, X.; Aparicio, E.M.; Murphy, T.C.; Duan, J.J.; Elkinton, J.S.; Gould, J.R. Assessing the host range of the North American parasitoid Ontsira mellipes: Potential for biological control of Asian longhorned beetle. Biol. Control 2019, 137, 104028. [Google Scholar] [CrossRef]
- Li, F.; Zhang, Y.L.; Wang, X.Y.; Cao, L.M.; Yang, Z.Q.; Gould, J.R.; Duan, J.J. Discovery of parasitoids of Anoplophora glabripennis (Coleoptera: Cerambycidae) and their seasonal abundance in China using sentinel host eggs and larvae. Econ. Entomol. 2020, 113, 1656–1665. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.; Hai, X.; Wang, Z. A review of the host plant location and recognition mechanisms of Asian longhorn beetle. Insects 2023, 14, 292. [Google Scholar] [CrossRef] [PubMed]
- Gorzlancyk, A.M.; Held, D.W.; Kim, D.-J.; Ranger, C.M. Capture of Xylosandrus crassiusculus and other Scolytinae (Coleoptera: Curculionidae) in response to visual and volatile cues. Fla. Entomol. 2013, 96, 1097–1101. [Google Scholar] [CrossRef]
- Green, E.W.; O’Callaghan, E.K.; Hansen, C.N.; Bastianello, S.; Bhutani, S.; Vanin, S.; Armstrong, J.D.; Costa, R.; Kyriacou, C.P. Drosophila circadian rhythms in seminatural environments: Summer afternoon component is not an artifact and requires TrpA1 channels. Proc. Natl. Acad. Sci. USA 2015, 112, 8702–8707. [Google Scholar] [CrossRef] [PubMed]
- Sakai, T.; Ishida, N. Circadian rhythms of female mating activity governed by clock genes in Drosophila. Proc. Natl. Acad. Sci. USA 2001, 98, 9221–9225. [Google Scholar] [CrossRef]
- Bloch, G.; Hazan, E.; Rafaeli, A. Circadian rhythms and endocrine functions in adult insects. J. Insect Physiol. 2013, 59, 56–69. [Google Scholar] [CrossRef]
- Zhao, K.; Ma, B.; Xu, Y.; Stirling, E.; Xu, J. Light exposure mediates circadian rhythms of rhizosphere microbial communities. ISME J. 2021, 15, 2655–2664. [Google Scholar] [CrossRef]
- Macias-Muñoz, A.; Rangel Olguin, A.G.; Briscoe, A.D. Evolution of phototransduction genes in Lepidoptera. Genome Biol. Evol. 2019, 11, 2107–2124. [Google Scholar] [CrossRef]
- Van Grunsven, R.H.A.; Donners, M.; Boekee, K.; Tichelaar, I.; van Geffen, K.G.; Groenendijk, D.; Berendse, F.; Veenendaal, E.M. Spectral composition of light sources and insect phototaxis, with an evaluation of existing spectral response models. J. Insect Conser. 2014, 18, 225–231. [Google Scholar] [CrossRef]
- Luo, C.; Chen, Y. Phototactic behavior of Scleroderma guani (Hymenoptera: Bethylidae)—Parasitoid of Pissodes punctatus (Coleoptera: Curculionidae). J. Insect Behav. 2016, 29, 605–614. [Google Scholar] [CrossRef]
- Yang, Z.; Wang, X.; Zhang, Y. Recent advances in biological control of important native and invasive forest pests in China. Biol. Control 2014, 68, 117–128. [Google Scholar] [CrossRef]
- Wang, Q.; Guo, Z.; Zhang, J.; Chen, Y.; Zhou, J.; Pan, Y.; Liu, X. Phototactic behavioral response of the ectoparasitoid beetle Dastarcus helophoroides (Coleoptera: Bothrideridae): Evidence for attraction by near-infrared light. J. Econ. Entomol. 2021, 114, 1549–1556. [Google Scholar] [CrossRef]
- Warrant, E.; Dacke, M. Vision and visual navigation in nocturnal insects. Annu. Rev. Entomol. 2011, 56, 239–254. [Google Scholar] [CrossRef] [PubMed]
- Wen, C.; Ma, T.; Deng, Y.; Liu, C.; Liang, S.; Wen, J.; Wang, C.; Wen, X. Morphological and optical features of the apposition compound eye of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Micron 2020, 128, 102769. [Google Scholar] [CrossRef] [PubMed]
- Little, C.M.; Chapman, T.W.; Hillier, N.K. Effect of color and contrast of highbush blueberries to host finding behavior by Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2018, 47, 1242–1251. [Google Scholar] [CrossRef] [PubMed]
- Zapponi, L.; Nieri, R.; Zaffaroni-Caorsi, V.; Pugno, N.M.; Mazzoni, V. Vibrational calling signals improve the efficacy of pheromone traps to capture the brown marmorated stink bug. J. Pest Sci. 2022, 96, 587–597. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light Wavelength (nm) | Spectrum Region | Main Wavelength (nm) | Peaking of Wavelength (nm) | Band Width of Peaking (nm) | Luminance (Lux) # |
|---|---|---|---|---|---|
| 365 | Ultraviolet | 472.0 | 365.6 | 12.7 | 24 |
| 385 | Ultraviolet | 388.0 | 385.2 | 12.0 | 180 |
| 400 | Violet | 370.2 | 402.1 | 15.8 | 180 |
| 420 | Violet | 427.1 | 420.9 | 17.6 | 144 |
| 435 | Violet | 441.0 | 435.0 | 17.2 | 36 |
| 455 | Blue | 460.3 | 455.5 | 20.9 | 180 |
| 475 | Blue | 477.2 | 473.9 | 25.4 | 180 |
| 500 | Green | 502.8 | 499.2 | 26.3 | 180 |
| 515 | Green | 522.4 | 515.7 | 32.0 | 180 |
| 560 | Green | 568.1 | 562.1 | 46.9 | 180 |
| 595 | Yellow | 592.6 | 596.3 | 14.6 | 180 |
| 600 | Orange | 595.2 | 599.5 | 15.7 | 180 |
| 630 | Red | 620.3 | 631.6 | 18.6 | 180 |
| 660 | Red | 641.1 | 659.6 | 17.1 | 180 |
| Wavelength (nm) | Sample (F, M) | Statistical Value | p |
|---|---|---|---|
| 365 | 38, 37 | 627.000 | 0.410 |
| 385 | 29, 38 | 524.500 | 0.734 |
| 400 | 29, 31 | 401.500 | 0.470 |
| 420 | 28, 31 | 431.000 | 0.963 |
| 435 | 31, 36 | 520.000 | 0.626 |
| 455 | 29, 34 | 470.000 | 0.746 |
| 475 | 29, 29 | 324.500 | 0.125 |
| 500 | 33, 31 | 364.500 | 0.045 |
| 515 | 37, 32 | 559.500 | 0.691 |
| 560 | 32, 36 | 524.500 | 0.518 |
| 595 | 32, 34 | 538.500 | 0.943 |
| 600 | 33, 33 | 503.500 | 0.590 |
| 630 | 31, 39 | 346.000 | 0.002 |
| 660 | 31, 35 | 490.000 | 0.494 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Hai, X.; Bi, Y.; Zhao, F.; Wang, Z.; Lyu, F. Research on Photoinduction-Based Technology for Trapping Asian Longhorned Beetle (Anoplophora glabripennis (Motschulsky, 1853) (Coleoptera: Cerambycidae). Insects 2023, 14, 465. https://doi.org/10.3390/insects14050465
Jiang X, Hai X, Bi Y, Zhao F, Wang Z, Lyu F. Research on Photoinduction-Based Technology for Trapping Asian Longhorned Beetle (Anoplophora glabripennis (Motschulsky, 1853) (Coleoptera: Cerambycidae). Insects. 2023; 14(5):465. https://doi.org/10.3390/insects14050465
Chicago/Turabian StyleJiang, Xianglan, Xiaoxia Hai, Yongguo Bi, Feng Zhao, Zhigang Wang, and Fei Lyu. 2023. "Research on Photoinduction-Based Technology for Trapping Asian Longhorned Beetle (Anoplophora glabripennis (Motschulsky, 1853) (Coleoptera: Cerambycidae)" Insects 14, no. 5: 465. https://doi.org/10.3390/insects14050465