
Machine Learning Modeling of Aedes albopictus Habitat Suitability in the 21st Century

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Datasets for Supervised Learning
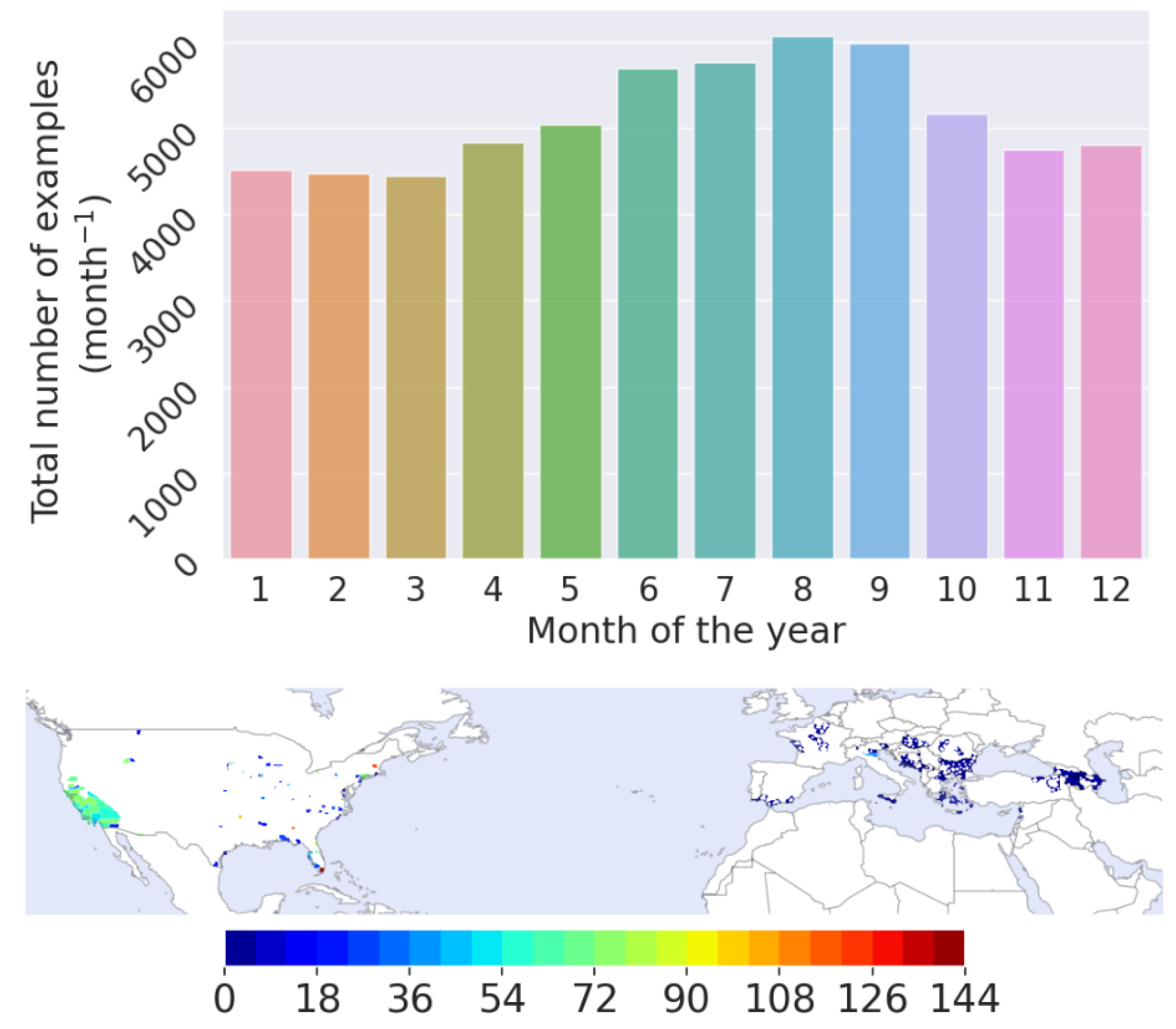
2.1.1. Vector Presence/Absence Dataset
- The Vectorbase PopBio (MapVEu tool) database was extracted for Ae. albopictus, for a period spanning from 2003 to 2021 [35]. The database was queried for Ae. albopictus in taxonomy and the “abundance” data type. The data request to the database included zero-counts.
- Ae. albopictus surveillance data from the Emilia-Romagna region in Italy for the years 2008–2012 [36]. These include bi-weekly surveillance data from ovitraps placed throughout the region.
- Surveillance data from Hungary (2017–2019), Slovenia (2016), and Serbia (2018) which were kindly provided by Prof. Dušan Petrić (University of Novi Sad), Dr. Kornélia Kurucz (University of Pécs), Dr. Katja Kalan (University of Primorska), and Dr. Ognyan Mikov (National Centre of Infectious and Parasitic Diseases, Bulgaria) [37].
- Data provided for the project Aedes challenge 2019 and 2020 from the Centre of Disease Control (CDC), accessed on 10 October 2021 (https://predict.cdc.gov), for Ae. albopictus. These data are provided in administrative units [38].
2.1.2. Feature Dataset
- Forested. Created by adding the primf (primary vegetation - potential forest land) and secdf (secondary vegetation - potential forest land) classes for each grid box/month.
- Non-forested. Created by adding the primn (primary vegetation - potential non-forest land) and secdn (secondary vegetation - potential non-forest land) classes.
- Crops. Created by adding the crops related classes; c3ann (C3 annual crops), c4ann (C4 annual crops), c3per (C3 perennial crops), c4per (C4 perennial crops), and c3nfx (C3 nitrogen-fixed crops).
- Graze land. Created by adding the pastr (managed pasture) and range (range land) classes.
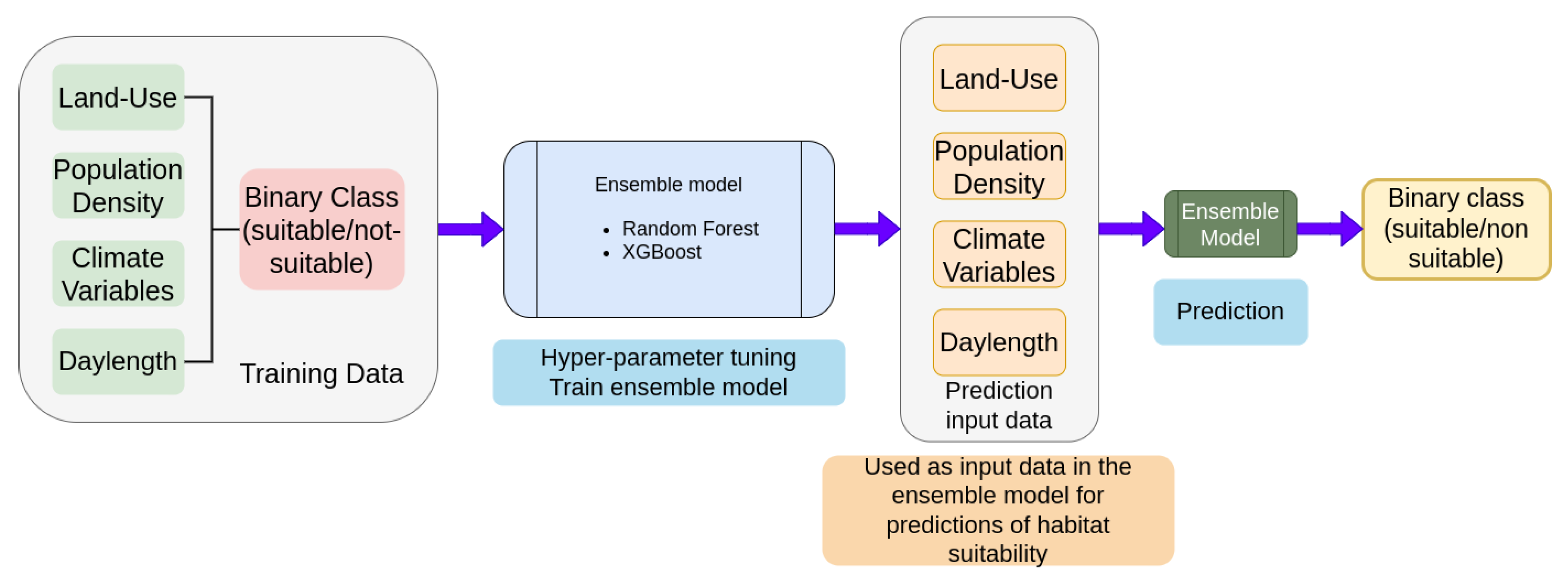
2.2. Machine Learning
2.3. Population at Risk
2.4. Maps
3. Results and Discussion
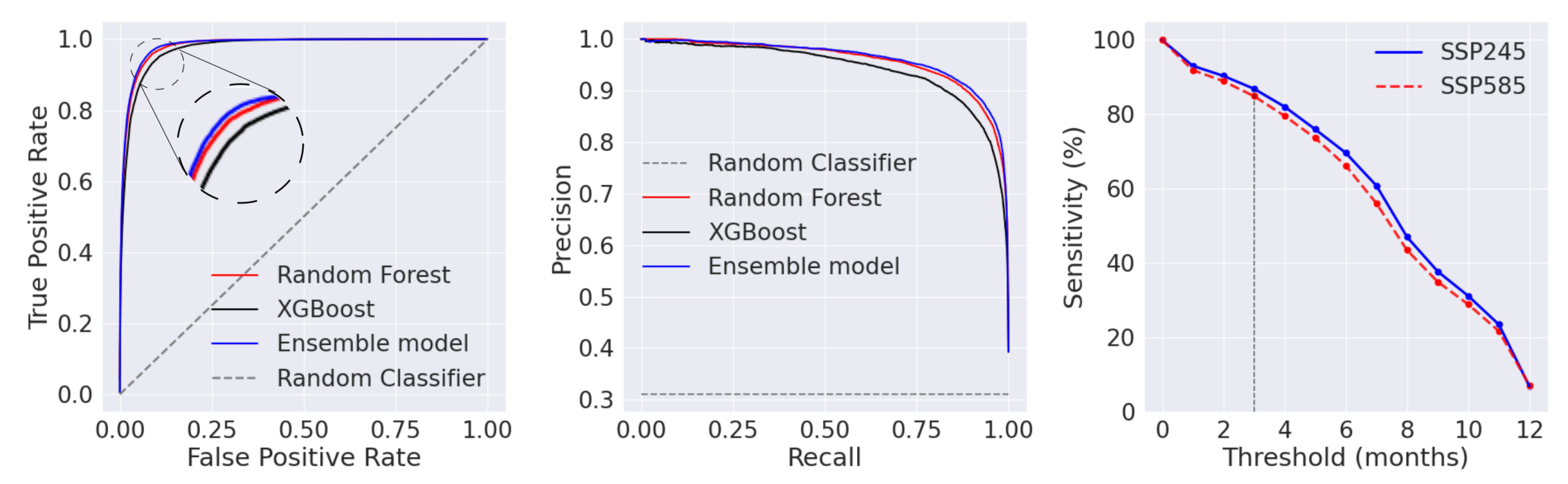
3.1. Machine Learning Model
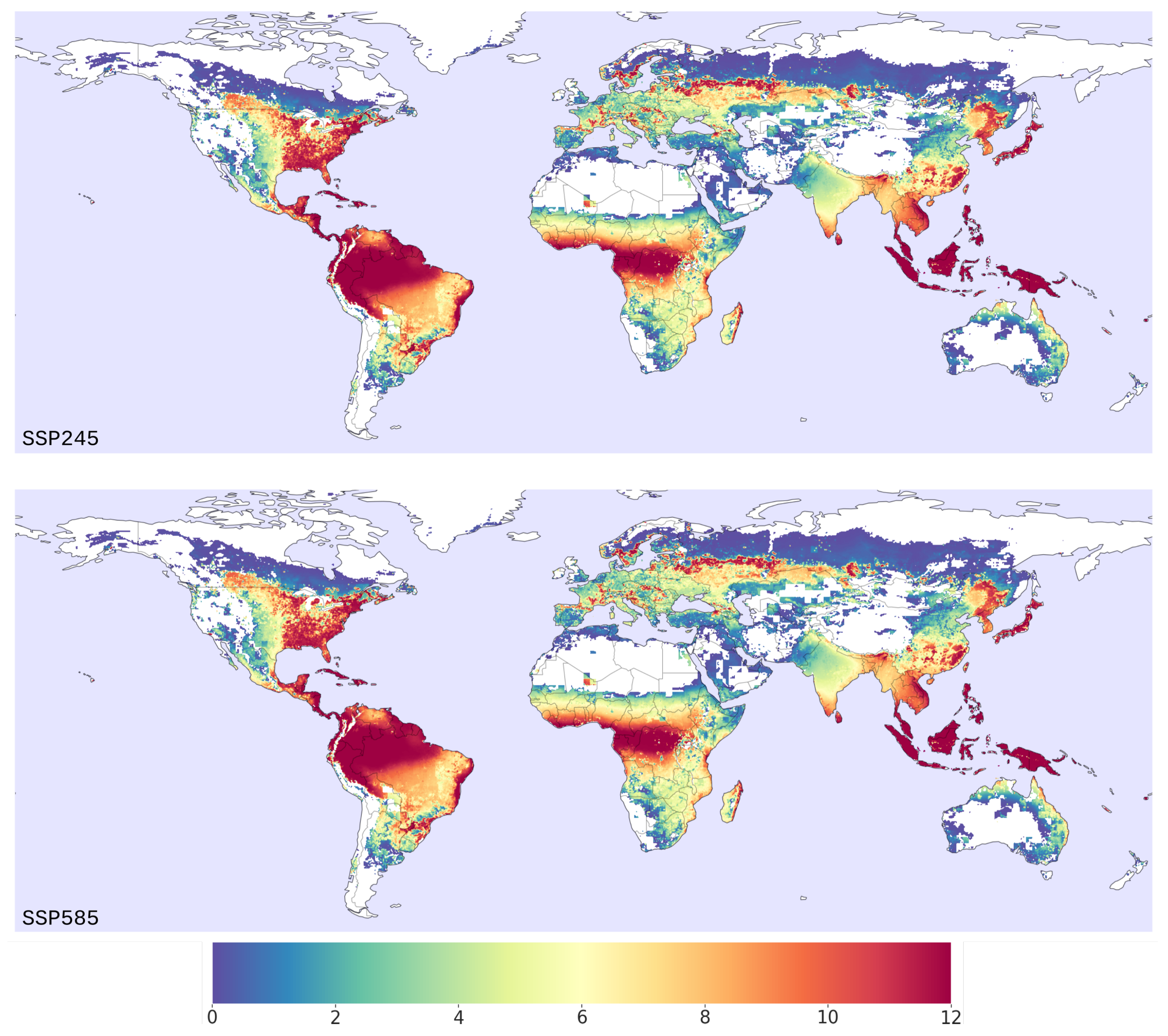
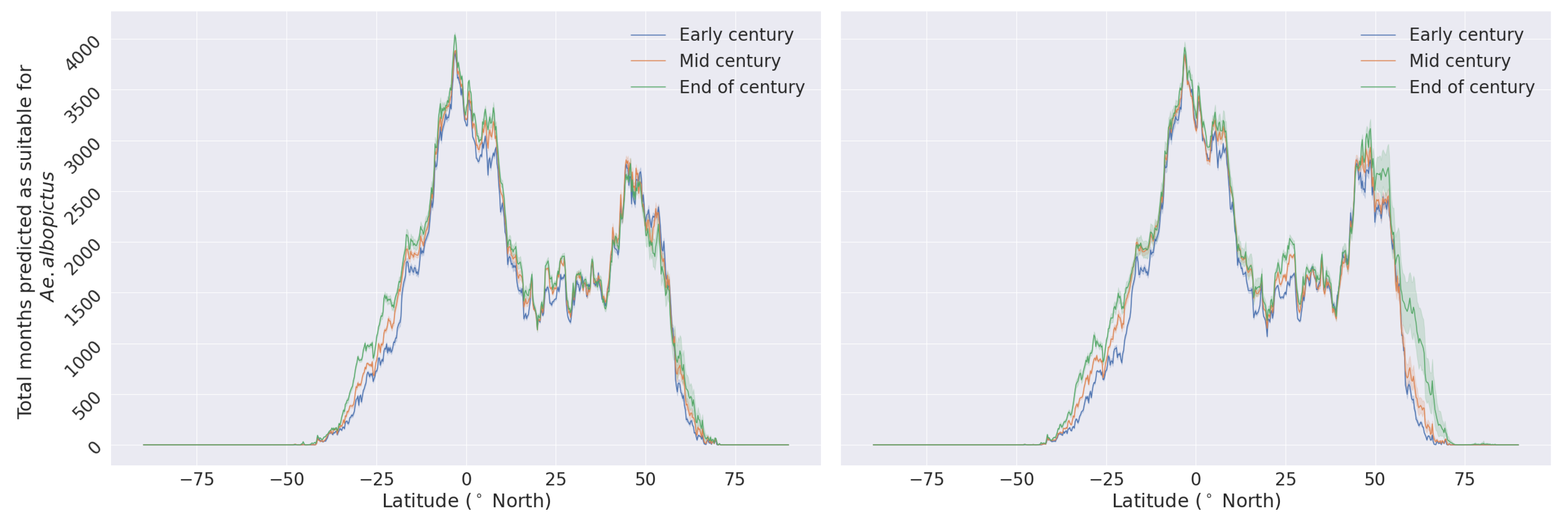
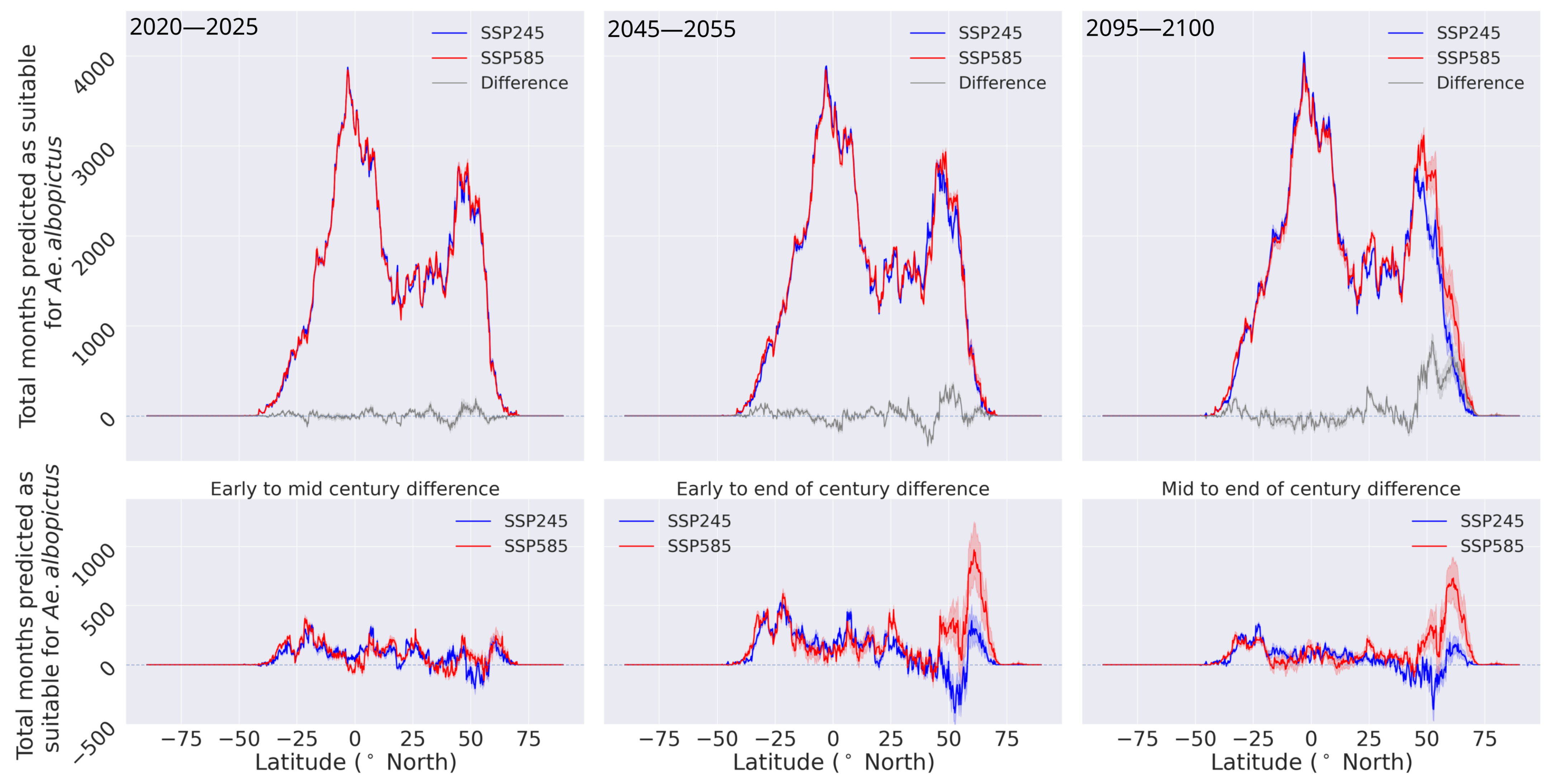
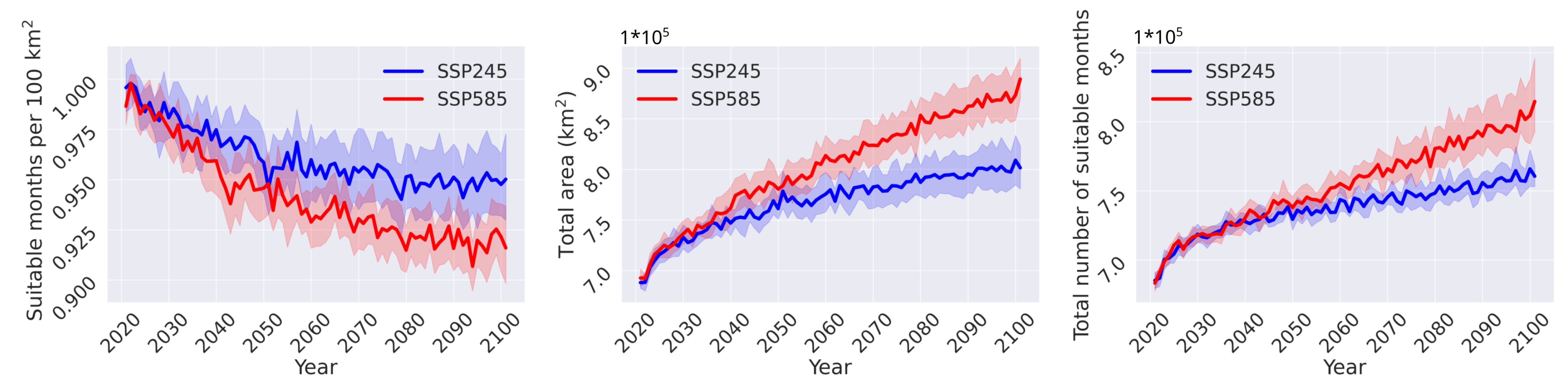
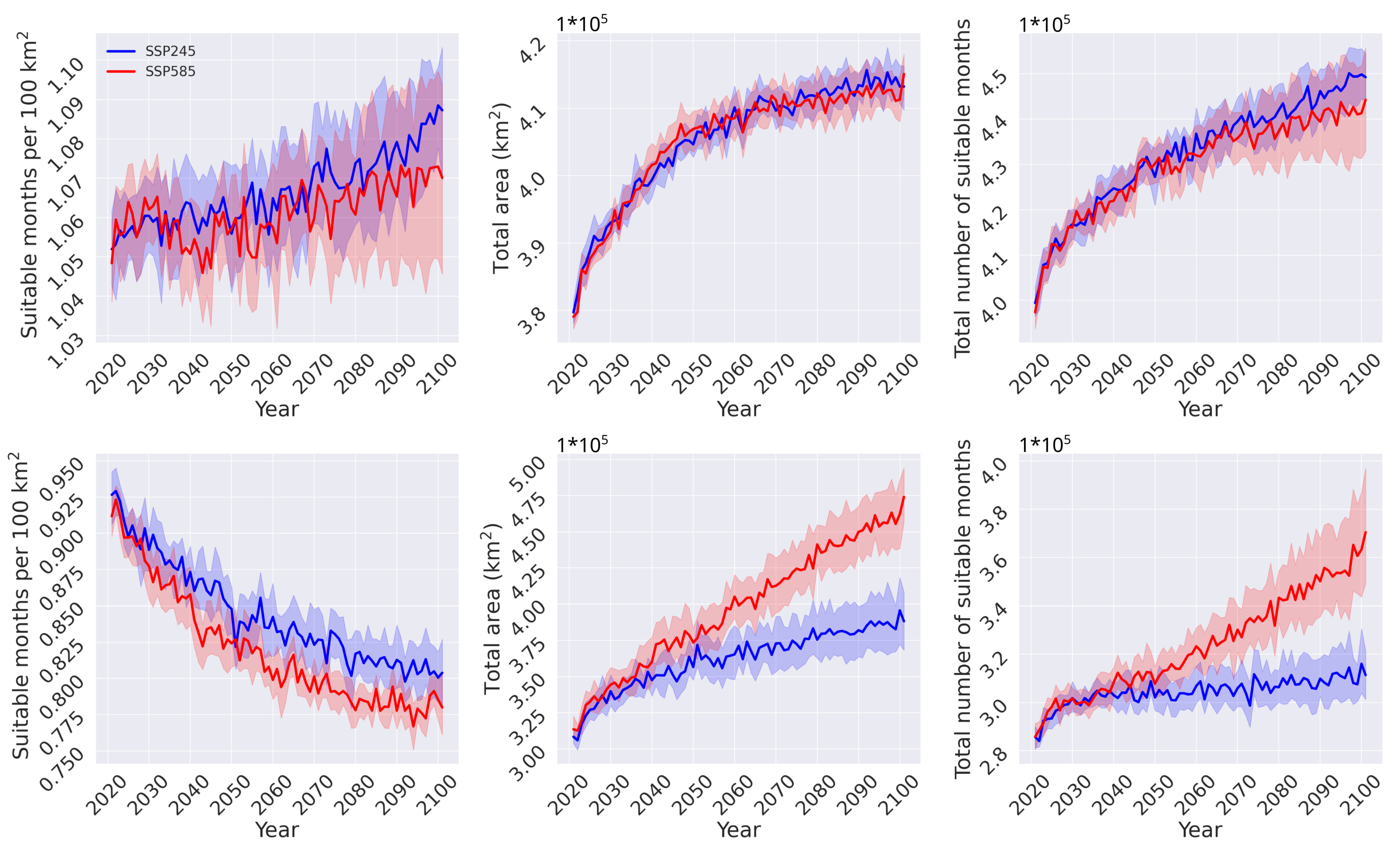
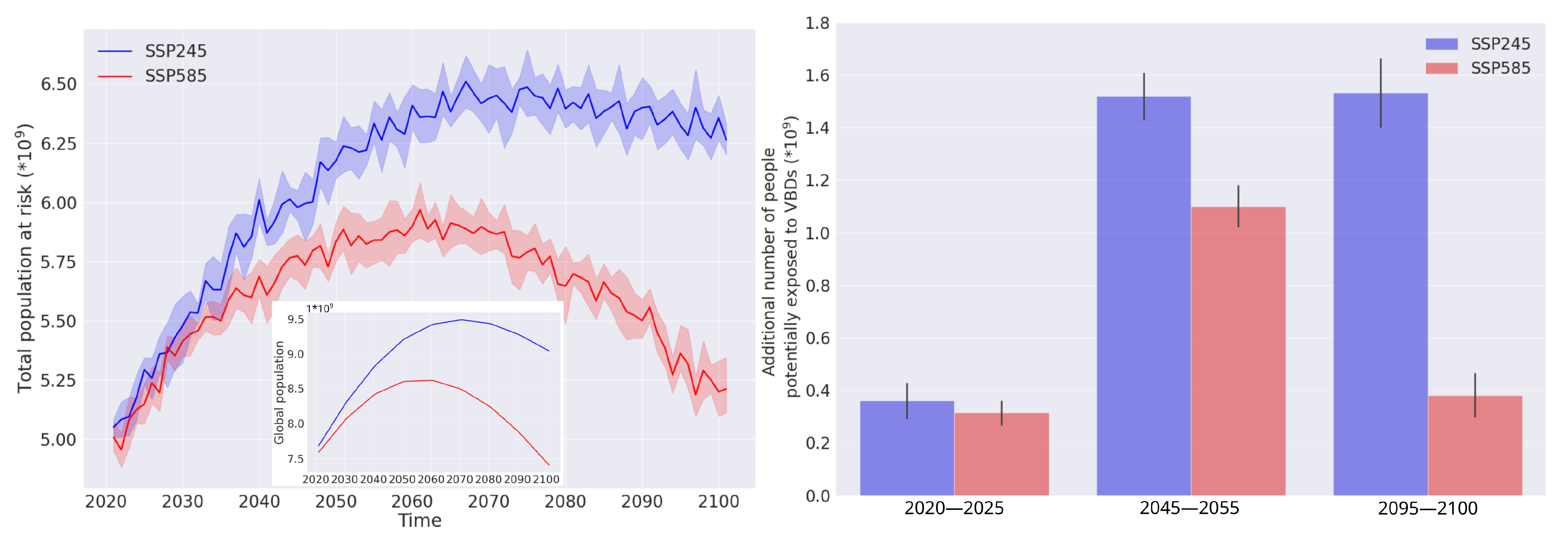
3.2. Habitat Suitability under Climate Change
4. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CMIP6 | Coupled model intercomparison project 6 |
| RCP | Representative concentration scenario |
| SSP | Shared socioeconomic pathway |
| ENM | Environmental niche model |
| BRT | Boost regression trees |
| SVM | Support vector machine |
| LUH2 | Land use harmonization 2 |
| NEX | Nasa earth exchange |
| GDDP | Global daily downscaled projections |
| ECDC | European centre for disease control |
| XGBoost | Extreme gradient boost |
| ROC | Receiver operating characteristic |
| AUC | Area under curve |
| ML | Machine learning |
| LSTM | Long short-term memory |
References
- Weaver, S.C.; Charlier, C.; Vasilakis, N.; Lecuit, M. Zika, Chikungunya, and Other Emerging Vector-Borne Viral Diseases. Annu. Rev. Med. 2018, 69, 395–408. [Google Scholar] [CrossRef] [PubMed]
- Messina, J.P.; Kraemer, M.U.; Brady, O.J.; Pigott, D.M.; Shearer, F.M.; Weiss, D.J.; Golding, N.; Ruktanonchai, C.W.; Gething, P.W.; Cohn, E.; et al. Mapping global environmental suitability for Zika virus. eLife 2016, 5, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Paixão, E.S.; Teixeira, M.G.; Rodrigues, L.C. Zika, chikungunya and dengue: The causes and threats of new and reemerging arboviral diseases. BMJ Glob. Health 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Delatte, H.; Dehecq, J.S.; Thiria, J.; Domerg, C.; Paupy, C.; Fontenille, D. Geographic distribution and developmental sites of Aedes albopictus (Diptera: Culicidae) during a Chikungunya epidemic event. Vector-Borne Zoonotic Dis. 2008, 8, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. Albopictus. eLife 2015, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Waldock, J.; Chandra, N.L.; Lelieveld, J.; Proestos, Y.; Michael, E.; Christophides, G.; Parham, P.E. The role of environmental variables on Aedes albopictus biology and chikungunya epidemiology. Pathog. Glob. Health 2013, 107, 224–241. [Google Scholar] [CrossRef] [PubMed]
- Ibáñez-Justicia, A.; Alcaraz-Hernández, J.D.; Van Lammeren, R.; Koenraadt, C.J.; Bergsma, A.; Delucchi, L.; Rizzoli, A.; Takken, W. Habitat suitability modeling to assess the introductions of Aedes albopictus (Diptera: Culicidae) in The Netherlands. Parasites Vectors 2020, 13, 217. [Google Scholar] [CrossRef]
- Reynolds, A.J.; Poelchau, M.F.; Rahman, Z.; Armbruster, P.A.; Denlinger, D.L. Transcript profiling reveals mechanisms for lipid conservation during diapause in the mosquito, Aedes albopictus. J. Insect Physiol. 2012, 58, 966–973. [Google Scholar] [CrossRef]
- Tatem, A.J.; Rogers, D.J.; Hay, S.I. Global Transport Networks and Infectious Disease Spread. Adv. Parasitol. 2006, 62, 293–343. [Google Scholar] [CrossRef] [PubMed]
- Roth, G.A.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, regional, and national age-sex-specific mortality for 282 causes of death in 195 countries and territories, 1980–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1736–1788. [Google Scholar] [CrossRef]
- Ryan, S.J.; Carlson, C.J.; Mordecai, E.A.; Johnson, L.R. Global expansion and redistribution of Aedes-borne virus transmission risk with climate change. PLoS Neglected Trop. Dis. 2018, 13, e0007213. [Google Scholar] [CrossRef] [PubMed]
- Egbendewe-Mondzozo, A.; Musumba, M.; McCarl, B.A.; Wu, X. Climate change and vector-borne diseases: An economic impact analysis of malaria in Africa. Int. J. Environ. Res. Public Health 2011, 8, 913–930. [Google Scholar] [CrossRef] [PubMed]
- Tatem, A.J.; Hay, S.I.; Rogers, D.J. Global traffic and disease vector dispersal. Proc. Natl. Acad. Sci. USA 2006, 103, 6242–6247. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Duan, Q.; Shen, C.; Wu, Y.; Xing, C. Global surface air temperatures in CMIP6: Historical performance and future changes. Environ. Res. Lett. 2020, 15, 104056. [Google Scholar] [CrossRef]
- Li, J.; Huo, R.; Chen, H.; Zhao, Y.; Zhao, T. Comparative Assessment and Future Prediction Using CMIP6 and CMIP5 for Annual Precipitation and Extreme Precipitation Simulation. Front. Earth Sci. 2021, 9, 430. [Google Scholar] [CrossRef]
- Reiter, P. Climate change and mosquito-borne disease. Environ. Health Perspect. 2001, 109, 141–161. [Google Scholar] [CrossRef] [PubMed]
- Proestos, Y.; Christophides, G.K.; Ergüler, K.; Tanarhte, M.; Waldock, J.; Lelieveld, J. Present and future projections of habitat suitability of the Asian tiger mosquito, a vector of viral pathogens, from global climate simulation. Philos. Trans. R. Soc. Biol. Sci. 2015, 370, 20130554. [Google Scholar] [CrossRef]
- Afrane, Y.A.; Githeko, A.K.; Yan, G. The ecology of Anopheles mosquitoes under climate change: Case studies from the effects of deforestation in East African highlands. Ann. N. Y. Acad. Sci. 2012, 1249, 204–210. [Google Scholar] [CrossRef]
- Liu, B.; Gao, X.; Zheng, K.; Ma, J.; Jiao, Z.; Xiao, J.; Wang, H. The potential distribution and dynamics of important vectors Culex pipiens pallens and Culex pipiens quinquefasciatus in China under climate change scenarios: An ecological niche modeling approach. Pest Manag. Sci. 2020, 76, 3096–3107. [Google Scholar] [CrossRef]
- Domingos, P. A few useful things to know about machine learning. Commun. ACM 2012, 55, 78–87. [Google Scholar] [CrossRef]
- Feng, X.; Park, D.S.; Walker, C.; Peterson, A.T.; Merow, C.; Papeş, M. A checklist for maximizing reproducibility of ecological niche models. Nat. Ecol. Evol. 2019, 3, 1382–1395. [Google Scholar] [CrossRef]
- Benedict, M.Q.; Levine, R.S.; Hawley, W.A.; Lounibos, L.P. Spread of the tiger: Global risk of invasion by the mosquito Aedes albopictus. Vector-Borne Zoonotic Dis. 2007, 7, 76–85. [Google Scholar] [CrossRef]
- Jia, P.; Lu, L.; Chen, X.; Chen, J.; Guo, L.; Yu, X.; Liu, Q. A climate-driven mechanistic population model of Aedes albopictus with diapause. Parasites Vectors 2016, 9, 175. [Google Scholar] [CrossRef]
- Kamal, M.; Kenawy, M.A.; Rady, M.H.; Khaled, A.S.; Samy, A.M. Mapping the global potential distributions of two arboviral vectors Aedes aegypti and Ae. albopictus under changing climate. PLoS ONE 2018, 13, e0210122. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M.; Elith, J.; Graham, C.H.; Lehmann, A.; Leathwick, J.; Ferrier, S. Sample selection bias and presence-only distribution models: Implications for background and pseudo-absence data. Ecol. Appl. 2009, 19, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Friedman, J.H. Stochastic gradient boosting. Comput. Stat. Data Anal. 2002, 38, 367–378. [Google Scholar] [CrossRef]
- Riahi, K.; Grübler, A.; Nakicenovic, N. Scenarios of long-term socio-economic and environmental development under climate stabilization. Technol. Forecast. Soc. Chang. 2007, 74, 887–935. [Google Scholar] [CrossRef]
- Wise, M.; Calvin, K.; Thomson, A.; Clarke, L.; Bond-Lamberty, B.; Sands, R.; Smith, S.J.; Janetos, A.; Edmonds, J. Implications of limiting CO2 concentrations for land use and energy. Science 2009, 324, 1183–1186. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Riahi, K. The role of non-CO2 greenhouse gases in climate change mitigation: Long-term scenarios for the 21st century. Energy J. 2006, 27, 177–200. [Google Scholar] [CrossRef]
- Smith, S.J.; Wigley, T. Multi-Gas Forcing Stabilization with Minicam. Energy J. 2006, 3, 373–392. [Google Scholar] [CrossRef]
- Clarke, L.E.; Wise, M.A.; Placet, M.; Izaurralde, R.C.; Lurz, J.P.; Kim, S.H.; Smith, S.J.; Thomson, A.M. Climate Change Mitigation: An Analysis of Advanced Technology Scenarios; Technical Report; Pacific Northwest National Lab.(PNNL): Richland, WA, USA, 2006.
- Wang, W.; Thrasher, B.; Michaelis, A.; Nemani, R.; Lee, T. The NASA Earth Exchange Global Daily Downscaled Projections. In Proceedings of the EGU General Assembly 2021, online, 19–30 April 2021. [Google Scholar] [CrossRef]
- Giraldo-Calderón, G.I.; Emrich, S.J.; MacCallum, R.M.; Maslen, G.; Emrich, S.; Collins, F.; Dialynas, E.; Topalis, P.; Ho, N.; Gesing, S.; et al. VectorBase: An updated Bioinformatics Resource for invertebrate vectors and other organisms related with human diseases. Nucleic Acids Res. 2015, 43, D707–D713. [Google Scholar] [CrossRef] [PubMed]
- Carrieri, M.; Albieri, A.; Angelini, P.; Baldacchini, F.; Venturelli, C.; Zeo, S.M.; Bellini, R. Surveillance of the chikungunya vector Aedes albopictus (Skuse) in Emilia-Romagna (northern Italy): Organizational and technical aspects of a large scale monitoring system. J. Vector Ecol. 2011, 36, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Kalan, K.; Ivovic, V.; Glasnovic, P.; Buzan, E. Presence and Potential Distribution of Aedes albopictus and Aedes japonicus japonicus (Diptera: Culicidae) in Slovenia. J. Med. Entomol. 2017, 54, 1510–1518. [Google Scholar] [CrossRef] [PubMed]
- CDC. Aedes Challenge. Available online: https://predict.cdc.gov/post/5c4f6d687620e103b6dcd015 (accessed on 10 December 2021).
- Hurtt, G.C.; Chini, L.; Sahajpal, R.; Frolking, S.; Bodirsky, B.L.; Calvin, K.; Doelman, J.C.; Fisk, J.; Fujimori, S.; Goldewijk, K.K.; et al. Harmonization of Global Land-Use Change and Management for the Period 850–2100 (LUH2) for CMIP6. Geosci. Model Dev. Discuss. 2020, 13, 5425–5464. [Google Scholar] [CrossRef]
- Fujimori, S.; Hasegawa, T.; Ito, A.; Takahashi, K.; Masui, T. Data descriptor: Gridded emissions and land use data for 2005–2100 under diverse socioeconomic and climate mitigation scenarios. Sci. Data 2018, 5, 180210. [Google Scholar] [CrossRef]
- Stephan Hoyer, A.K.; Brevdo, E. Xarray. 2014. Available online: https://github.com/pydata/xarray (accessed on 10 December 2021).
- Jones, B.; O’Neill, B.C. Spatially explicit global population scenarios consistent with the Shared Socioeconomic Pathways. Environ. Res. Lett. 2016, 11, 084003. [Google Scholar] [CrossRef]
- Brock, T.D. Calculating solar radiation for ecological studies. Ecol. Model. 1981, 14, 1–19. [Google Scholar] [CrossRef]
- Forsythe, W.C.; Rykiel, E.J.; Stahl, R.S.; Wu, H.i.; Schoolfield, R.M. A model comparison for daylength as a function of latitude and day of year. Ecol. Model. 1995, 80, 87–95. [Google Scholar] [CrossRef]
- Ziehn, T.; Chamberlain, M.A.; Law, R.M.; Lenton, A.; Bodman, R.W.; Dix, M.; Stevens, L.; Wang, Y.P.; Srbinovsky, J. The Australian Earth System Model: ACCESS-ESM1.5. J. South. Hemisph. Earth Syst. Sci. 2020, 70, 193–214. [Google Scholar] [CrossRef]
- Döscher, R.; Acosta, M.; Alessandri, A.; Anthoni, P.; Arsouze, T.; Bergman, T.; Bernardello, R.; Boussetta, S.; Caron, L.-P.; Carver, G.; et al. The EC-Earth3 Earth System Model for the Climate Model Intercomparison Project 6. Geosci. Model Dev. Discuss. 2022, 15, 2973–3020. [Google Scholar] [CrossRef]
- Adcroft, A.; Anderson, W.; Balaji, V.; Blanton, C.; Bushuk, M.; Dufour, C.O.; Dunne, J.P.; Griffies, S.M.; Hallberg, R.; Harrison, M.J.; et al. The GFDL Global Ocean and Sea Ice Model OM4.0: Model Description and Simulation Features. J. Adv. Model. Earth Syst. 2019, 11, 3167–3211. [Google Scholar] [CrossRef]
- Li, L.; Yu, Y.; Tang, Y.; Lin, P.; Xie, J.; Song, M.; Dong, L.; Zhou, T.; Liu, L.; Wang, L.; et al. The Flexible Global Ocean-Atmosphere-Land System Model Grid-Point Version 3 (FGOALS-g3): Description and Evaluation. J. Adv. Model. Earth Syst. 2020, 12, e2019MS002012. [Google Scholar] [CrossRef]
- Volodin, E.M.; Mortikov, E.V.; Kostrykin, S.V.; Galin, V.Y.; Lykossov, V.N.; Gritsun, A.S.; Diansky, N.A.; Gusev, A.V.; Iakovlev, N.G.; Shestakova, A.A.; et al. Simulation of the modern climate using the INM-CM48 climate model. Russ. J. Numer. Anal. Math. Model. 2018, 33, 367–374. [Google Scholar] [CrossRef]
- Volodin, E.M.; Gritsun, A.S. Simulation of Possible Future Climate Changes in the 21st Century in the INM-CM5 Climate Model. Izv.—Atmos. Ocean. Phys. 2020, 56, 218–228. [Google Scholar] [CrossRef]
- Tatebe, H.; Ogura, T.; Nitta, T.; Komuro, Y.; Ogochi, K.; Takemura, T.; Sudo, K.; Sekiguchi, M.; Abe, M.; Saito, F.; et al. Description and basic evaluation of simulated mean state, internal variability, and climate sensitivity in MIROC6. Geosci. Model Dev. 2019, 12, 2727–2765. [Google Scholar] [CrossRef]
- Yukimoto, S.; Kawai, H.; Koshiro, T.; Oshima, N.; Yoshida, K.; Urakawa, S.; Tsujino, H.; Deushi, M.; Tanaka, T.; Hosaka, M.; et al. The meteorological research institute Earth system model version 2.0, MRI-ESM2.0: Description and basic evaluation of the physical component. J. Meteorol. Soc. Jpn. 2019, 97, 931–965. [Google Scholar] [CrossRef]
- Seland, Ø..; Bentsen, M.; Seland Graff, L.; Olivié, D.; Toniazzo, T.; Gjermundsen, A.; Debernard, J.B.; Gupta, A.K.; He, Y.; Kirkevåg, A.; et al. The Norwegian Earth System Model, NorESM2—Evaluation of theCMIP6 DECK and historical simulations. Geosci. Model Dev. Discuss. 2020, 1–68. [Google Scholar]
- Pedregosa, F.; Varoquaux, G.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine Learning in Python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Buitinck, L.; Louppe, G.; Blondel, M.; Pedregosa, F.; Mueller, A.; Grisel, O.; Niculae, V.; Prettenhofer, P.; Gramfort, A.; Grobler, J.; et al. API design for machine learning software: Experiences from the scikit-learn project. In Proceedings of the ECML PKDD Workshop: Languages for Data Mining and Machine Learning, 1 September 2013; pp. 108–122. [Google Scholar]
- Tharwat, A. Classification assessment methods. Appl. Comput. Inform. 2018, 17, 168–192. [Google Scholar] [CrossRef]
- Caminade, C.; Medlock, J.M.; Ducheyne, E.; McIntyre, K.M.; Leach, S.; Baylis, M.; Morse, A.P. Suitability of European climate for the Asian tiger mosquito Aedes albopictus: Recent trends and future scenarios. J. R. Soc. Interface 2012, 9, 2708–2717. [Google Scholar] [CrossRef] [PubMed]
- Petric, M.; Ducheyne, E.; Gossner, C.M.; Marsboom, C.; Nicolas, G.; Venail, R.; Hendrickx, G.; Schaffner, F. Seasonality and timing of peak abundance of aedes albopictus in europe: Implications to public and animal health. Geospat. Health 2021, 16. [Google Scholar] [CrossRef]
- Eritja, R.; Palmer, J.R.; Roiz, D.; Sanpera-Calbet, I.; Bartumeus, F. Direct Evidence of Adult Aedes albopictus Dispersal by Car. Sci. Rep. 2017, 7, 14399. [Google Scholar] [CrossRef] [PubMed]
- Office, M. Cartopy: A Cartographic Python Library with a Matplotlib Interface; Exeter: Devon, UK, 2010. [Google Scholar]
- Sammut, C.; Webb, G.I. (Eds.) Area Under Curve. In Encyclopedia of Machine Learning; Springer: Boston, MA, USA, 2010; p. 40. [Google Scholar] [CrossRef]
- Erguler, K.; Smith-Unna, S.E.; Waldock, J.; Proestos, Y.; Christophides, G.K.; Lelieveld, J.; Parham, P.E. Large-scale modeling of the environmentally-driven population dynamics of temperate aedes albopictus (Skuse). PLoS ONE 2016, 11, e0149282. [Google Scholar] [CrossRef]
- Erguler, K.; Chandra, N.L.; Proestos, Y.; Lelieveld, J.; Christophides, G.K.; Parham, P.E. A large-scale stochastic spatiotemporal model for Aedes albopictus-borne chikungunya epidemiology. PLoS ONE 2017, 12, e0174293. [Google Scholar] [CrossRef]
- Johnson, T.L.; Haque, U.; Monaghan, A.J.; Eisen, L.; Hahn, M.B.; Hayden, M.H.; Savage, H.M.; McAllister, J.; Mutebi, J.P.; Eisen, R.J. Modeling the Environmental Suitability for Aedes (Stegomyia) aegypti and Aedes (Stegomyia) albopictus (Diptera: Culicidae) in the Contiguous United States. J. Med. Entomol. 2017, 54, 1605–1614. [Google Scholar] [CrossRef]
- Cunze, S.; Kochmann, J.; Koch, L.K.; Klimpel, S. Aedes albopictus and its environmental limits in Europe. PLoS ONE 2016, 11, e0162116. [Google Scholar] [CrossRef] [PubMed]
- Tjaden, N.B.; Suk, J.E.; Fischer, D.; Thomas, S.M.; Beierkuhnlein, C.; Semenza, J.C. modeling the effects of global climate change on Chikungunya transmission in the 21 st century. Sci. Rep. 2017, 7, 3813. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Fu, J.; Jiang, D.; Hao, M.; Lin, G. Mapping the spatial distribution of Aedes aegypti and Aedes albopictus. Acta Tropica 2018, 178, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Früh, L.; Kampen, H.; Kerkow, A.; Schaub, G.A.; Walther, D.; Wieland, R. modeling the potential distribution of an invasive mosquito species: Comparative evaluation of four machine learning methods and their combinations. Ecol. Model. 2018, 388, 136–144. [Google Scholar] [CrossRef]
- Cui, G.; Zhong, S.; Zheng, T.; Li, Z.; Zhang, X.; Li, C.; Hemming-Schroeder, E.; Zhou, G.; Li, Y. Aedes albopictus life table: Environment, food, and age dependence survivorship and reproduction in a tropical area. Parasites Vectors 2021, 14, 568. [Google Scholar] [CrossRef] [PubMed]
- Xia, D.; Guo, X.; Hu, T.; Li, L.; Teng, P.Y.; Yin, Q.Q.; Luo, L.; Xie, T.; Wei, Y.H.; Yang, Q.; et al. Photoperiodic diapause in a subtropical population of Aedes albopictus in Guangzhou, China: Optimized field-laboratory-based study and statistical models for comprehensive characterization. Infect. Dis. Poverty 2018, 7, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Paupy, C.; Delatte, H.; Bagny, L.; Corbel, V.; Fontenille, D. Aedes albopictus, an arbovirus vector: From the darkness to the light. Microbes Infect. 2009, 11, 1177–1185. [Google Scholar] [CrossRef]
- Valerio, L.; Marini, F.; Bongiorno, G.; Facchinelli, L.; Pombi, M.; Caputo, B.; Maroli, M.; Della Torre, A. Host-feeding patterns of aedes albopictus (Diptera: Culicidae) in urban and rural contexts within Rome province, Italy. Vector-Borne Zoonotic Dis. 2010, 10, 291–294. [Google Scholar] [CrossRef]
- Pereira dos Santos, T.; Roiz, D.; Santos de Abreu, F.V.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Santos Neves, M.S.A.; Simard, F.; Lourenço-de Oliveira, R.; Paupy, C. Potential of Aedes albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerg. Microbes Infect. 2018, 7, 1–8. [Google Scholar] [CrossRef]
- Kraemer, M.U.; Reiner, R.C.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Armbruster, P.A. Photoperiodic Diapause and the Establishment of Aedes albopictus (Diptera: Culicidae) in North America. J. Med. Entomol. 2016, 53, 1013–1023. [Google Scholar] [CrossRef]
- Tebaldi, C.; Debeire, K.; Eyring, V.; Fischer, E.; Fyfe, J.; Friedlingstein, P.; Knutti, R.; Lowe, J.; O’Neill, B.; Sanderson, B.; et al. Climate model projections from the Scenario Model Intercomparison Project (ScenarioMIP) of CMIP6. Earth Syst. Dyn. 2021, 12, 253–293. [Google Scholar] [CrossRef]
- Messina, J.P.; Brady, O.J.; Golding, N.; Kraemer, M.U.; Wint, G.R.; Ray, S.E.; Pigott, D.M.; Shearer, F.M.; Johnson, K.; Earl, L.; et al. The current and future global distribution and population at risk of dengue. Nat. Microbiol. 2019, 4, 1508–1515. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Long Name | Ref. |
|---|---|---|
| ACCESS-ESM1-5 | Australian Community Climate and Earth System Simulator (ACCESS) | [45] |
| EC-Earth3 | EC-Earth European Consortium | [46] |
| GFDL-CM4 | Geophysical Fluid Dynamics Laboratory (GFDL) | [47] |
| FGOALS-g3 | Flexible Global Ocean-Atmosphere-Land System Model Grid Point Version 3 | [48] |
| INM-CM4-8 | Institute of Numerical Mathematics (INM) | [49] |
| INM-CM5-0 | Institute of Numerical Mathematics (INM) | [50] |
| MIROC6 | Model for Interdisciplinary Research on Climate | [51] |
| MRI-ESM2-0 | Meteorological Research Institute Earth System Model Version 2.0 | [52] |
| NorESM2-MM | Norwegian Earth System Model | [53] |
| Name | Long Name | Units |
|---|---|---|
| tas | Average temperature | C |
| tasmin | Minimum temperature | C |
| tasmax | Maximum temperature | C |
| tp | Total precipitation | mm |
| hurs | Relative humidity | % |
| pop_density | Population density | per sq. km |
| daylength | Day length | hours |
| urban | Urban land use | Fraction coverage |
| crops | Crops related land use | Fraction coverage |
| forested | Potential forest land use | Fraction coverage |
| non-forested | Potential non-forest land use | Fraction coverage |
| graze-land | Grazing land use | Fraction coverage |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgiades, P.; Proestos, Y.; Lelieveld, J.; Erguler, K. Machine Learning Modeling of Aedes albopictus Habitat Suitability in the 21st Century. Insects 2023, 14, 447. https://doi.org/10.3390/insects14050447
Georgiades P, Proestos Y, Lelieveld J, Erguler K. Machine Learning Modeling of Aedes albopictus Habitat Suitability in the 21st Century. Insects. 2023; 14(5):447. https://doi.org/10.3390/insects14050447
Chicago/Turabian StyleGeorgiades, Pantelis, Yiannis Proestos, Jos Lelieveld, and Kamil Erguler. 2023. "Machine Learning Modeling of Aedes albopictus Habitat Suitability in the 21st Century" Insects 14, no. 5: 447. https://doi.org/10.3390/insects14050447