1. Introduction
The brown stink bug,
Euschistus heros (Hemiptera: Pentatomidae), is one of the key pests in soybean,
Glycine max (L.) Merr., grown in Brazil. As a piercing-sucking insect, it is able to feed on both the plant and the grains, which can cause indirect damage to the plants and direct damage to yields [
1]. It is a neotropical pentatomid and due to the increase in global temperature in the past decades,
E. heros attacks soybean crops across Brazil [
2].
The majority of producers still exclusively use chemical control for managing
E. heros [
3]. Although chemical control is one of the pillars of Integrated Pest Management (IPM), it should be used as the last management tactic, always based on pest monitoring, and following the recommended dose and rotating mode of action. Unfortunately, the traditional culture of the producers and the lack of different compounds for the management of pentatomids has resulted in overuse of chemical pesticides and widespread insecticide resistance [
4]. In addition, over reliance on chemical pesticides has caused a decrease in natural enemies, environmental damage, and an increase in production costs [
5,
6].
The use of biological control has been increasing for pest management in crops as a way to reduce the use of pesticides and increase the sustainability of the agroecosystem, while preventing economic damage [
7]. The egg parasitoid
Telenomus podisi (Hymenoptera: Scelionidae) has been shown to be effective in managing populations of
E. heros, the preferred host [
8]. By parasitizing the egg stage, it has advantages in preventing the establishment and feeding damage of the pest [
9]. Currently, around 60,000 hectares of soybean in Brazil are managed for
E. heros with the release of
T. podisi, with potential for expansion in the coming years [
3,
10].
For the release of
T. podisi to occur effectively, a sufficient amount of parasitoids must be produced in specialized biofactories. However, the production of
T. podisi requires substantial labor since the shelf life of natural enemies is low. An additional challenge is that, in most cases, the parasitoids are used at specific times of the year, depending on the planting cycle of each crop [
11,
12]. Large-scale insect rearing aims to reduce costs and increase production without reducing parasitoid quality [
13,
14]. Currently, techniques using artificial diets [
15,
16,
17] and storage of host eggs at low temperatures [
11,
18,
19] help in reducing rearing costs. Parasitoids also require hosts eggs with good quality and sufficient nutritional value [
20]. Consequently, the egg aspect and content of nutrients in mass rearing are essential for parasitism, providing enough parasitoids for field release.
The development of research in quality control of
T. podisi is still new in Brazil, as the commercialization of this parasitoid was only registered in 2019 [
21]. The reproductive rates of parasitoids are the basis for evaluating the control potential of a species as a biological control agent applied with inundative releases in the field [
22]. Biological parameters, including sex ratio and viability, can also affect the success of parasitism in the field [
23]. In addition, climate conditions strongly influence insect physiological development, migration, and dispersal [
24]. Therefore, studies to optimize rearing of
T. podisi based on biological attributes and rearing temperature are essential to understand the main requirements of this natural enemy, improving mass production with satisfactory quality to manage pests in the field [
23].
The objectives of this study were to evaluate the parasitism capacity, thermal requirements, and biological characteristics of T. podisi that emerged from E. heros eggs. Eggs were obtained from adult stink bugs fed with different diets and used fresh or stored eggs that were frozen in liquid nitrogen. All conditions were tested at different temperatures (15 ± 2, 18 ± 2, 21 ± 2, 24 ± 2, 27 ± 2, 30 ± 2, and 33 ± 2 °C). Our work provides new information for T. podisi rearing to assist in the success of biological control programs against E. heros in Brazil and elsewhere.
4. Discussion
The interaction between parasitoid and host depends on many factors, with temperature and egg quality being among the most important parameters that influence parasitism and survival [
23,
24,
31,
32]. Temperature directly influences life process in small insects [
33]. Our parasitism capacity results show an inverse relationship between temperature and survival period. As observed in most insects, the increase in temperature results in a decreased lifespan. This can be explained by increases in the insect’s metabolic rate at high temperatures and decreased metabolism at low temperatures [
34,
35]. Similar effects of high temperatures on survivorship and parasitism capacity were found for several other parasitoids species [
34,
36,
37,
38,
39,
40].
In the cases of applied inundative biological control programs, the parasitism capacity is more relevant than the period of time females remain alive in the field [
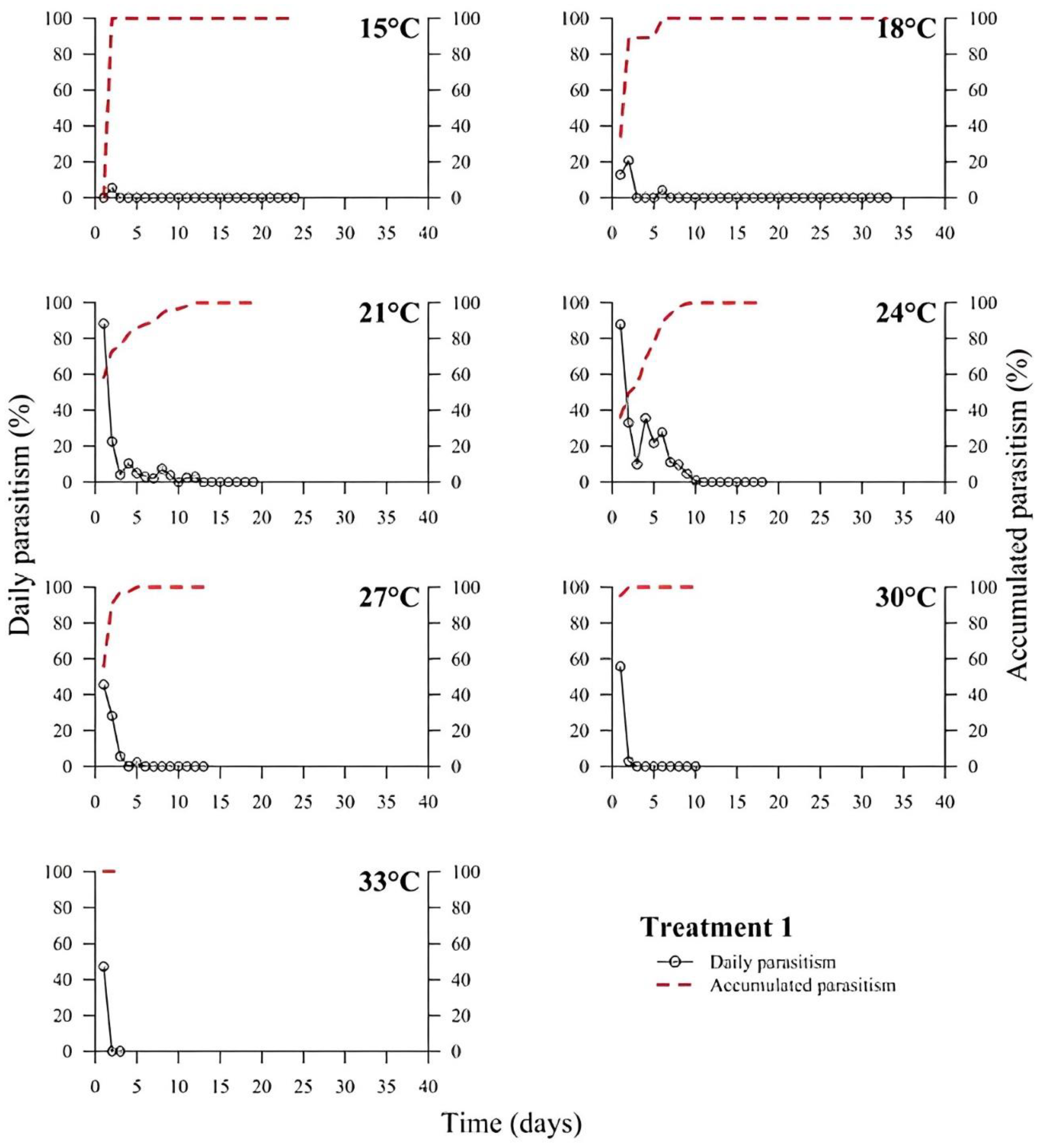
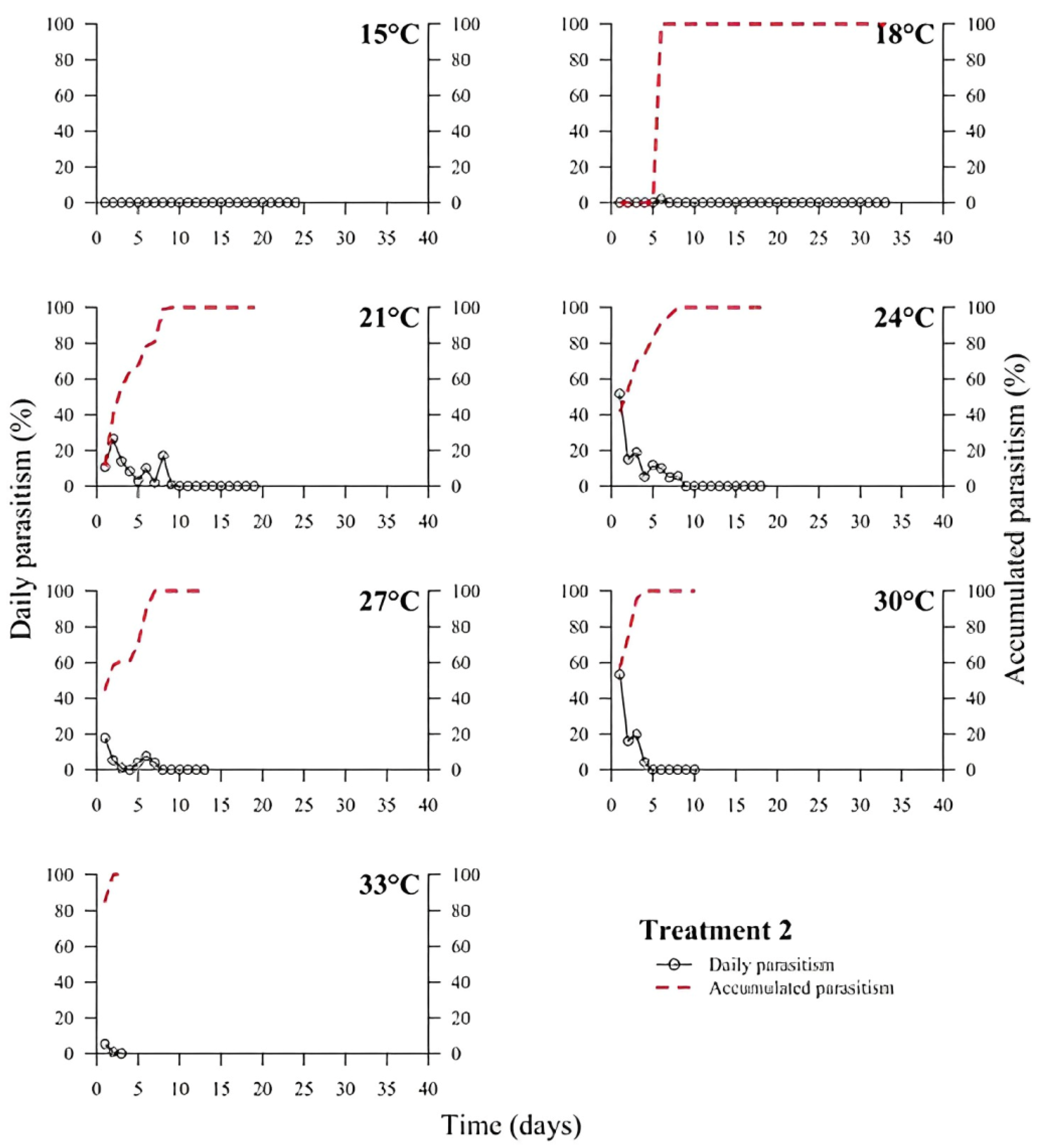
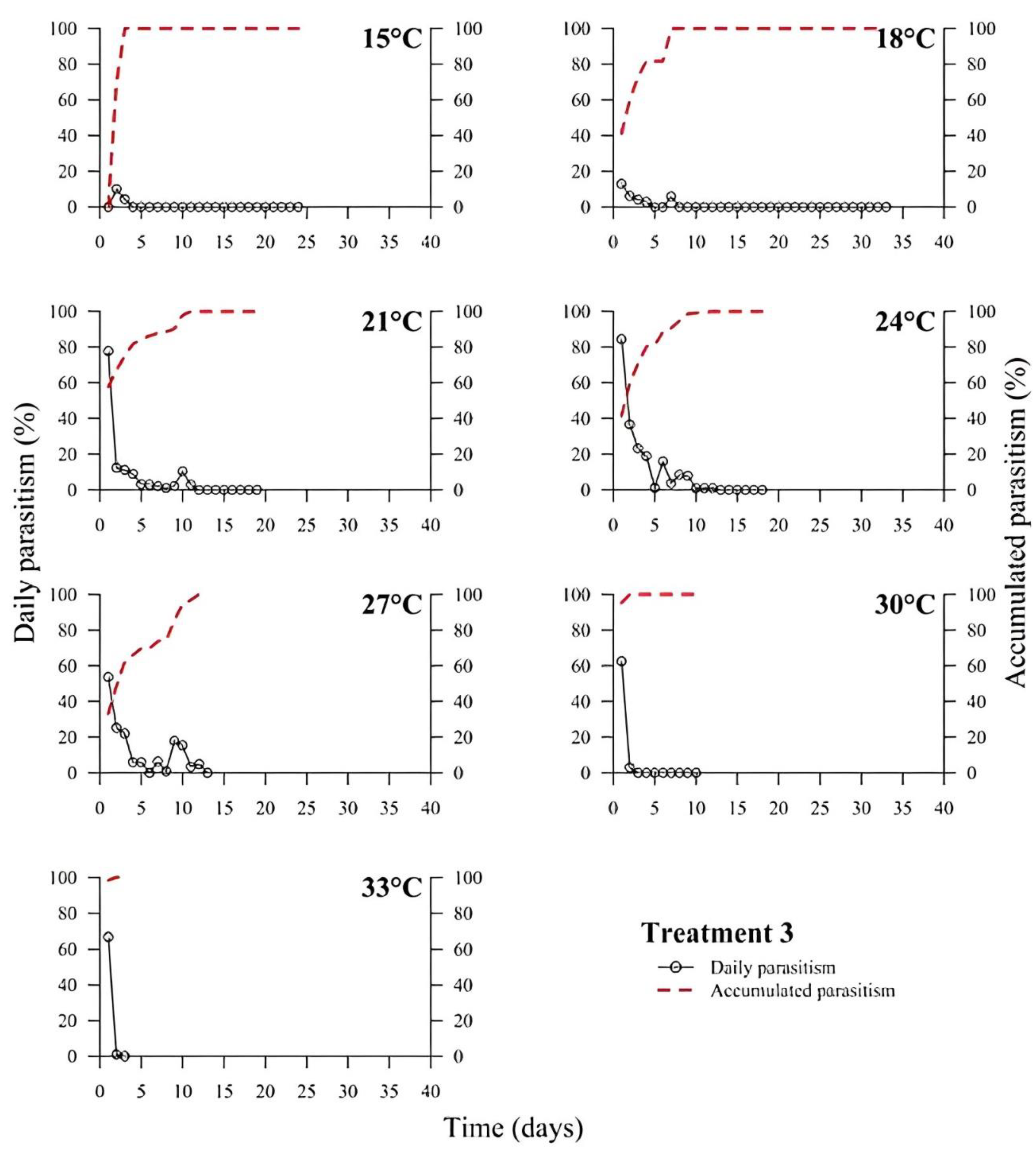
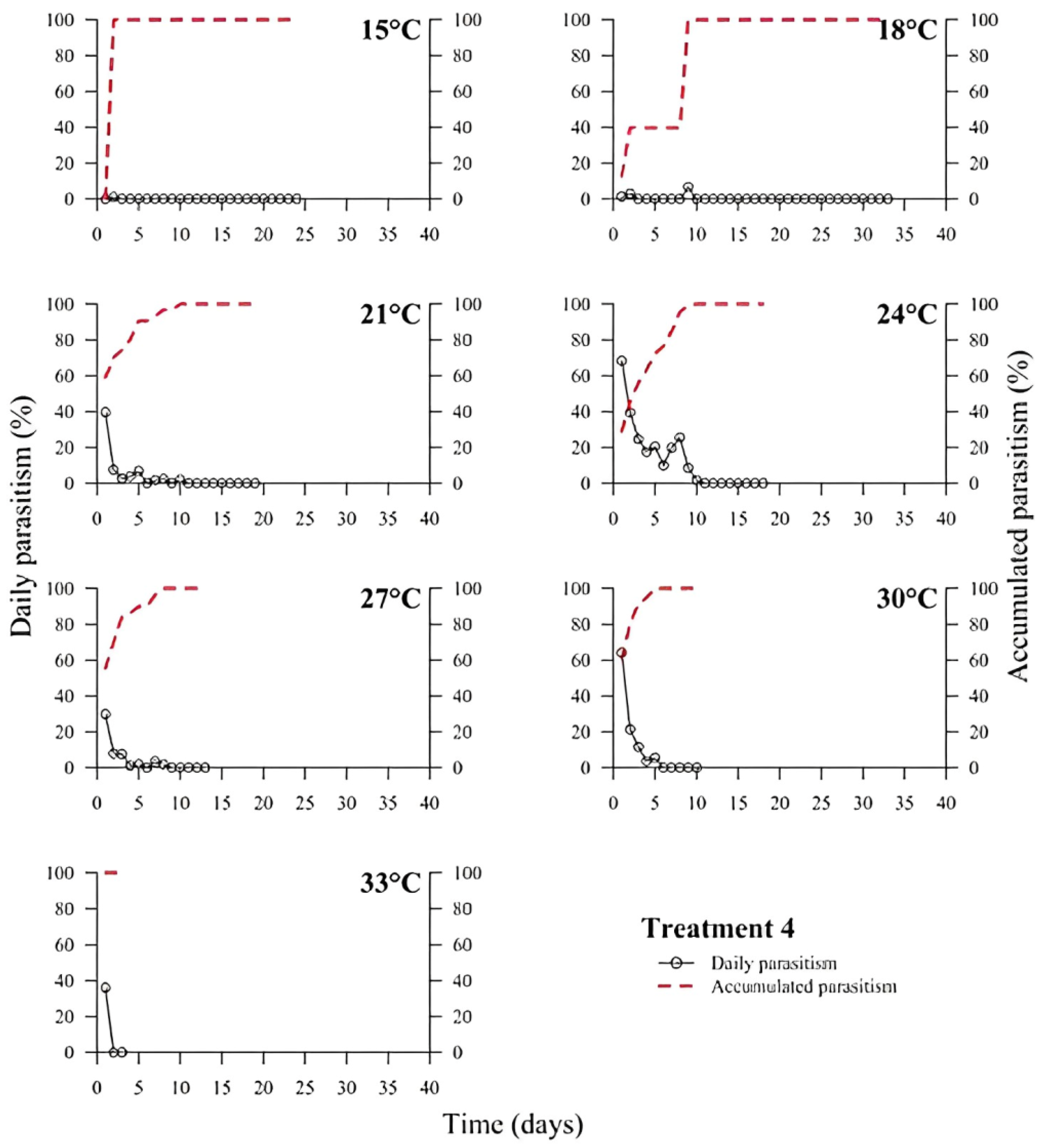
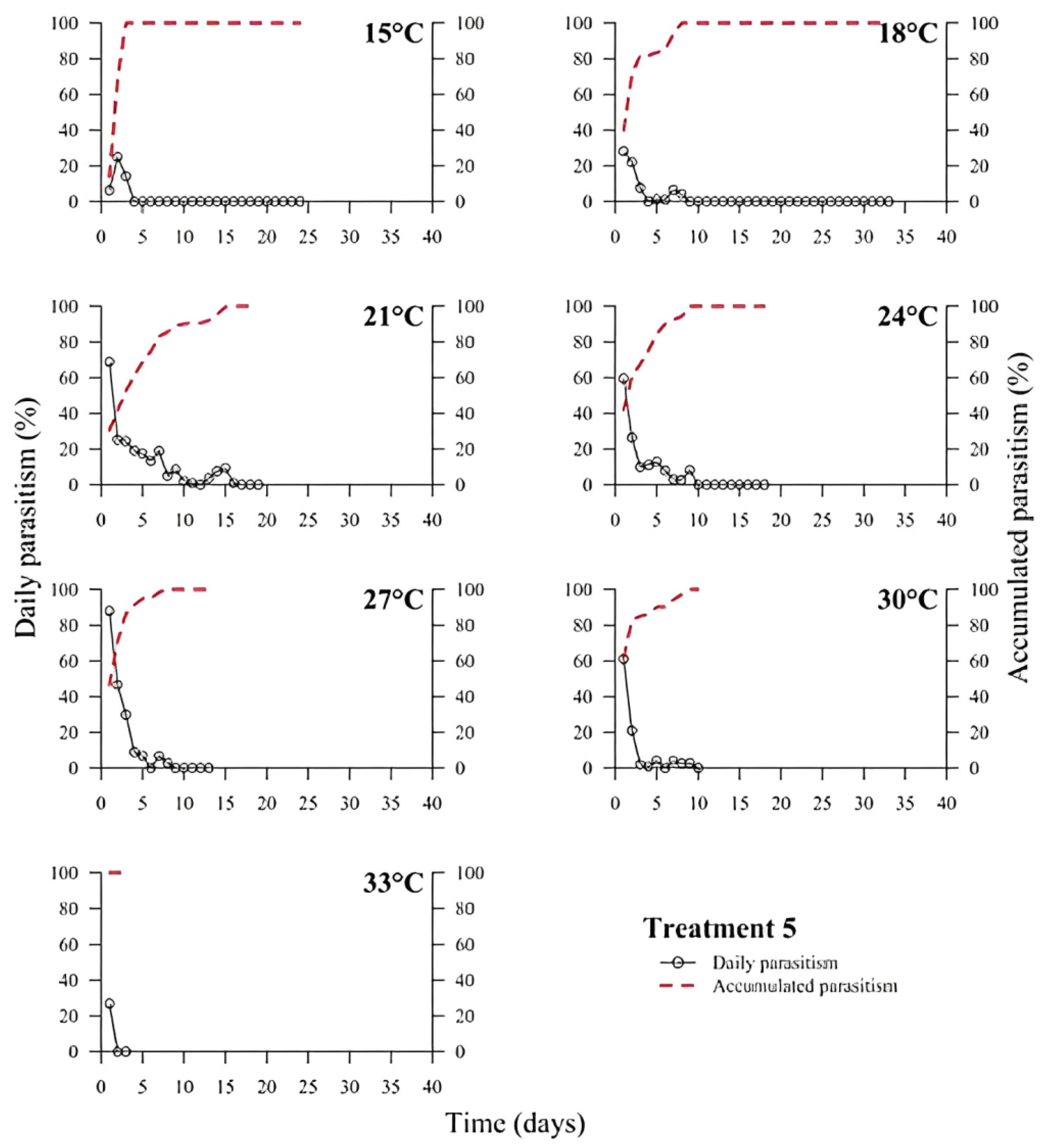
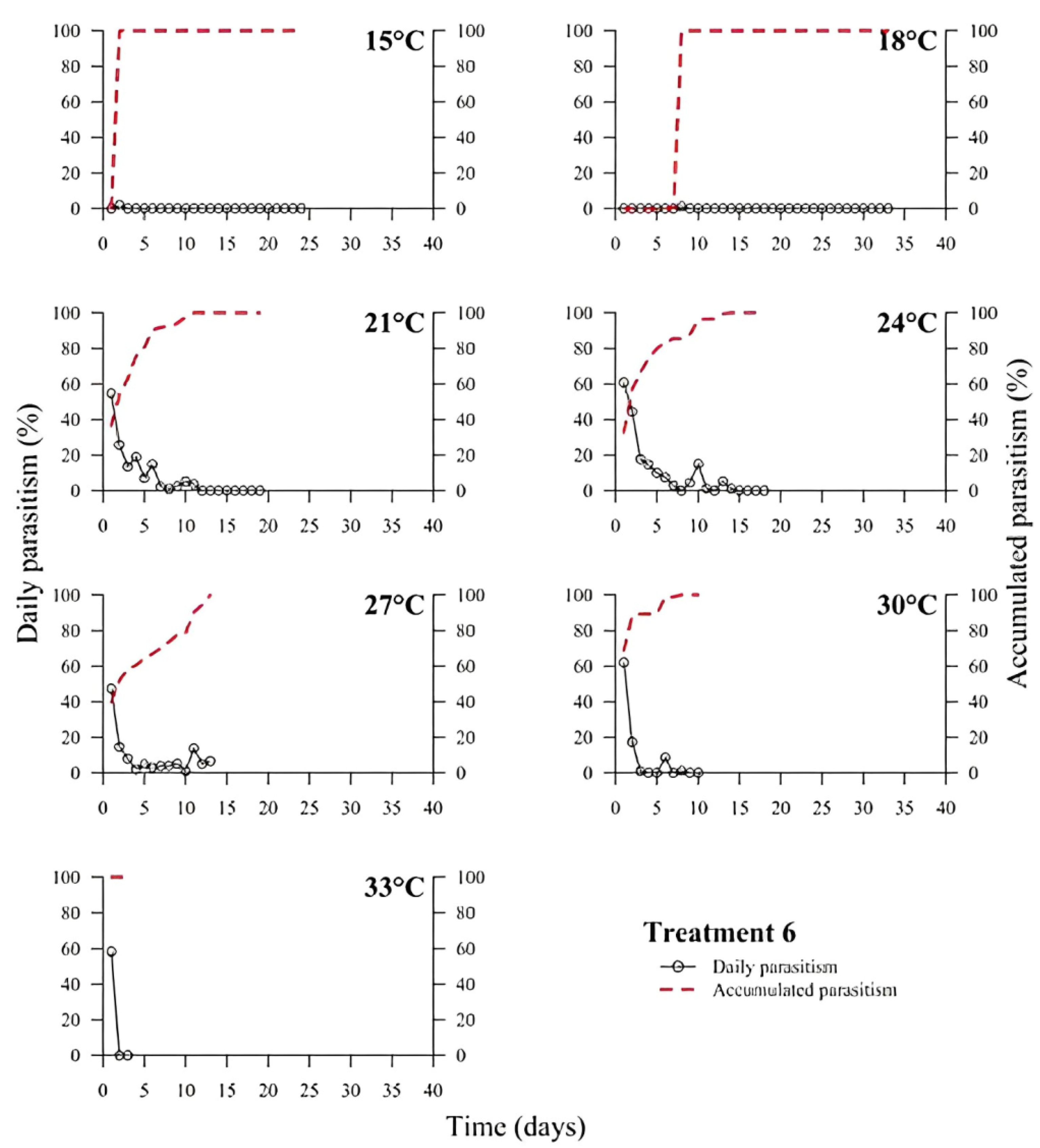
39]. In all treatments, the highest rate of parasitism occurred in the first days (
Figure 1,
Figure 2,
Figure 3,
Figure 4,
Figure 5 and
Figure 6). This can be explained by the ability of
T. podisi to store mature eggs in the ovaries or oviducts, completing oogenesis even before the adult emerges from the host egg [
41].
According to the International Organization for Biological Control (IOBC) guidelines, ≥80% is accepted as a good parameter of parasitism for the genus
Trichogramma [
42]. Based on that, the goal of 80% of parasitism was not achieved in most treatments, except for T5 at 27 °C. Species, such as
T. podisi, are unable to perform lipogenesis and depend entirely on lipid storage in the larval stage for fecundity and lifespan in the adult stage [
33,
39,
43,
44]. In addition, since they cannot synthesize lipids in the adult stage, parasitoids are more vulnerable to temperature increases and pesticides than most pests, including the host
E. heros [
39,
43,
45]. Therefore, nutritional aspects are essential for the success in parasitism, and parasitoids tend to choose sources that provide greater energy return [
20,
46].
Our results showed that artificial diets had better parasitism rates at 15, 18, and 27 °C than treatments with natural diets. These data emphasize the importance of studies on the influence of eggs from hosts reared with artificial diets in the production of parasitoids. Possibly, eggs from artificial diets had superior nutritional quality, resulting in greater parasitism under stressful conditions (thermal extremes), which has not been reported in other studies with artificial diets that examined the biology of
T. podisi [
16,
47]. Future host preference tests and field bioassays should be carried out, in addition to the analysis of egg nutrition, influenced by diet and storage.
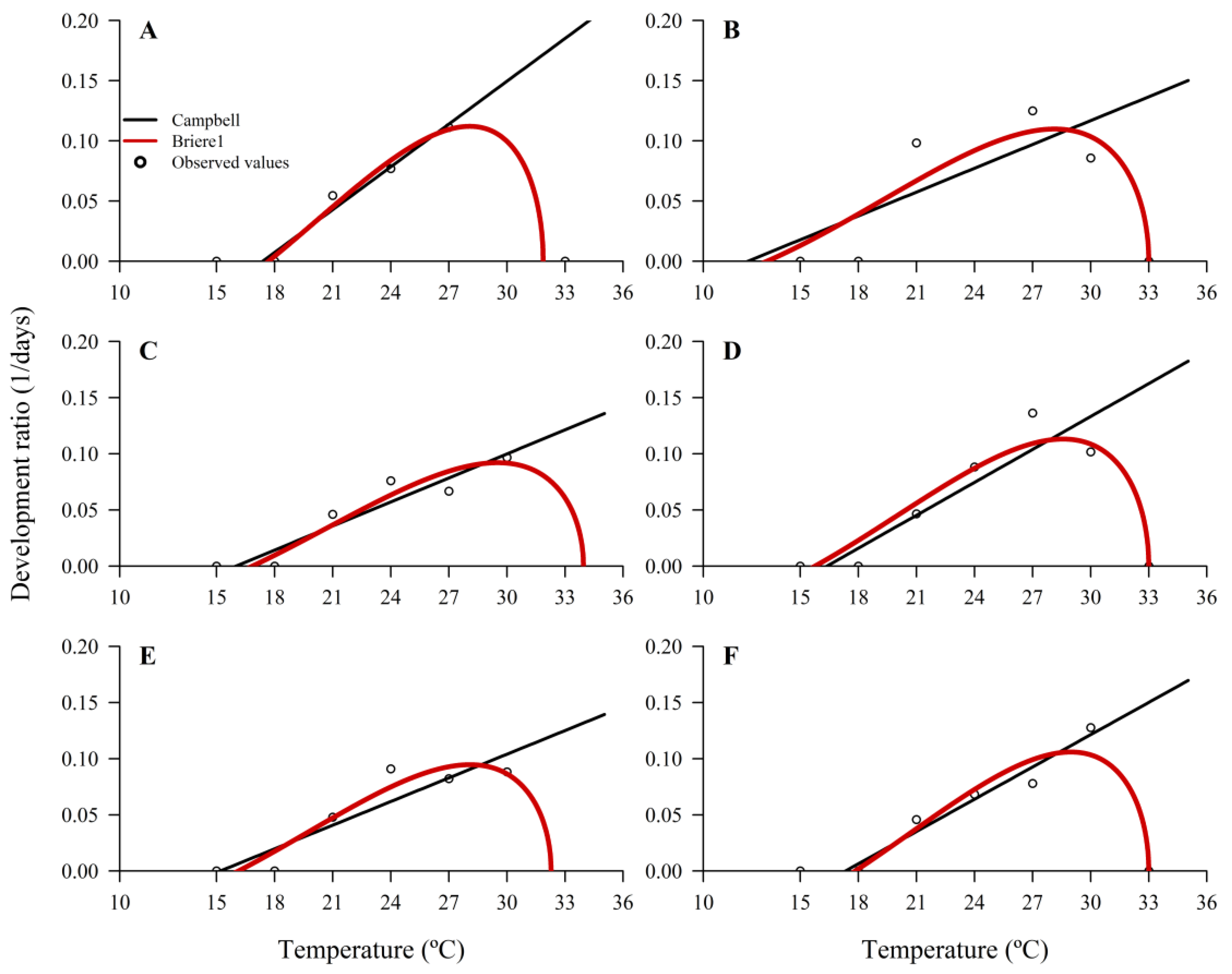
Rearing for biological control release also depends on providing parasitoids when host eggs are deposited. The efficiency and development of
T. podisi are temperature-dependent, allowing for some storage of adults, and the ability to increase or decrease development [
24,
32]. From thermal requirement data, given by the thermal constant
K (degree-days), upper and lower temperature threshold (
Tmax and
Tmin), and optimal temperature (
Topt), it is possible to estimate the influence of temperature on the parasitoid’s lifespan [
48,
49], making it possible to optimize the mass production of natural enemies. The hosts (
E. heros) and the parasitoids used in this bioassay originate from colonies kept in the laboratory for years, in which the hosts are reared under the same conditions as T1 (natural diet and fresh eggs). Possibly, since they are adapted to this type of diet, they needed less thermal accumulation to complete development and future studies should examine the effects of temperature on successive generations.
The host–parasitoid interaction can be influenced by associative learning over generations, where parasitoids adapt and prefer certain types of hosts due to pre-emergence conditioning established during their larval period [
50,
51,
52].
E. heros, the preferred host for
T. podisi, has a
K of 327 degree-days [
35], which is significantly higher than all treatments. This is a good indication that, in favorable situations in the field, the parasitoid will complete its cycle faster than the host, further contributing to pest management [
34,
53].
Values of
K,
Tmin,
Tmax, and
Topt vary according to the type of host used, species of parasitoid, and adjusted model for the analyses, potentially relating our results to those of other studies [
22,
34,
35,
54,
55,
56,
57]. Since T2 presented
Tmin of 16 °C and was composed of a natural diet (little variety of ingredients) and cryopreserved eggs, the nutritional aspect of the eggs from this diet was potentially inferior compared to the other treatments, influencing
Tmin.
Quality control parameters are essential for mass rearing of insects. This is especially challenging for biological control insects that must be produced as needed to match the crop and target pest. The ability to maintain laboratory colonies and quickly produce the desired number of individuals requires the creation of artificial diet, ways to store host eggs for long periods, and the ability to accelerate or slow down development. These manipulations must not impact the overall quality of the reared insects [
7,
23,
39]. In Europe, there are quality control guidelines for more than 30 species of natural enemies according to the IOBC, but the majority are species not produced in Brazil [
42]. However, these guidelines serve as a model for national quality control tests [
58,
59].
Based on the analysis of biotic factors, including adult size and life history, across rearing temperatures, we can assess the quality of
T. podisi for release in the field [
60], since the success of this parasitoid release depends on these quality parameters [
23,
39]. Even though we found significant factorial interaction between the diets and the temperatures tested, there are no expressly significant differences between body length based on the diet and cryopreservation of eggs offered to
T. podisi. Body size may be relevant in reducing energy costs [
61,
62]. Larger insects may reduce movements to search for hosts, are likely to forage in a more heterogeneous environment, interacting with more species, and exhibit greater preferential prey selection than smaller individuals [
61,
62,
63,
64]. As only females are responsible for parasitism in the field and differences were found only in body length of females at 21 °C, these data can be useful to improve the success of the parasitoid in biological control programs at this temperature.
Temperature was important in the rearing of
T. podisi, since survival was prolonged at lower temperatures. Low temperatures can reduce the insect’s metabolic rate, affecting the viability of parasitized eggs, consequently preventing insect emergence [
34,
57,
65]. The loss of turgidity of the eggs as the temperature increases can prevent the insertion of the parasitoid ovipositor into the egg’s chorion, reducing successful parasitism [
39,
66].
Considering the release of parasitoids in the field, these results may indicate an important factor in applied biological control programs due to increases in average global temperature, mainly in regions with high thermal amplitude (low temperatures at night and high during the day). Thermal extremes can be harmful to the survival, development, dispersion, and longevity of parasitoids, compromising the efficiency of parasitoid release and management of target pests [
24]. With the increase in global temperature, our results of the upper temperature threshold are relevant for mass releases of
T. podisi in regions with high temperatures, considering that
Topt ranged between 28 and 29 °C. Parasitism rates of 60% were found when
T. podisi was reared at 27 and 30 °C, demonstrating that
T. podisi is capable of parasitizing more than half of pentatomid eggs soon after release at higher field temperatures [
39,
43]. Increases in average global temperature tend to negatively affect the distribution of parasitoids more than of hosts, due to lower
Topt and
Tmax [
54,
67,
68]. In an ideal scenario, delaying the development of parasitoids when field temperatures border on the upper threshold can help in the successful application of biological control. Our results can be useful for biological programs in areas with thermal ranges from 21 to 30 °C, indicating that
T. podisi is adapted to these conditions, although field experiments must confirm our results.
The nutritional composition of the egg is also fundamental for the choice and development of the parasitoid [
69]. Since AD2 is rich in different types of grains (sunflower seed and wheat germ) compared to the AD1 and the natural diet (
Table 1), the
E. heros eggs obtained from AD2 likely had a higher nutritional value than the other diets, allowing for the development of parasitoids at high temperatures (33 °C). Future nutritional analysis of diets and eggs should be performed to confirm this assumption. In addition, the production cost of each diet should be analyzed to conclude whether the shift from natural to artificial diet is beneficial.
The artificial diets, especially AD2 with fresh eggs (T5), showed better results than the natural diet, contradicting other studies in which artificial diets did not influence the evaluated parameters of
T. podisi [
16,
47]. Viability and sex ratio had better results with fresh eggs than cryopreserved eggs, possibly due to small changes in the physical-nutritional characteristics caused by low temperatures, causing females to prefer fresh eggs [
20]. In order to increase fitness, foraging theory predicts that parasitoids, when searching for host eggs, evaluate and explore the energy content before parasitizing the egg [
20,
46,
70,
71]. Several studies have been carried out on the use of cryopreservation in mass rearing of egg parasitoids [
11,
72,
73,
74,
75]. Although the use of liquid nitrogen to store host eggs is beneficial to maintain a stock throughout the year, our results demonstrated that cold storage affected the biological performance of
T. podisi.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}