
Is Integrated Management of Gymnandrosoma aurantianum Possible with Trichogramma atopovirilia and Novel Products Used in Citrus Orchards in Brazil?
 ,
,  ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Insecticides
2.3. Evaluation of Lethal and Sublethal Effects on Adults of T. atopovirilia
2.4. Evaluation of Lethal and Sublethal Effects on Pupae of T. atopovirilia
2.5. Residual Activity (Persistence) of Insecticides to T. atopovirilia
2.6. Statistical Analysis
3. Results
3.1. Evaluation of Lethal and Sublethal Effects on Adults of T. atopovirilia
3.2. Evaluation of Lethal and Sublethal Effects on Pupae of T. atopovirilia
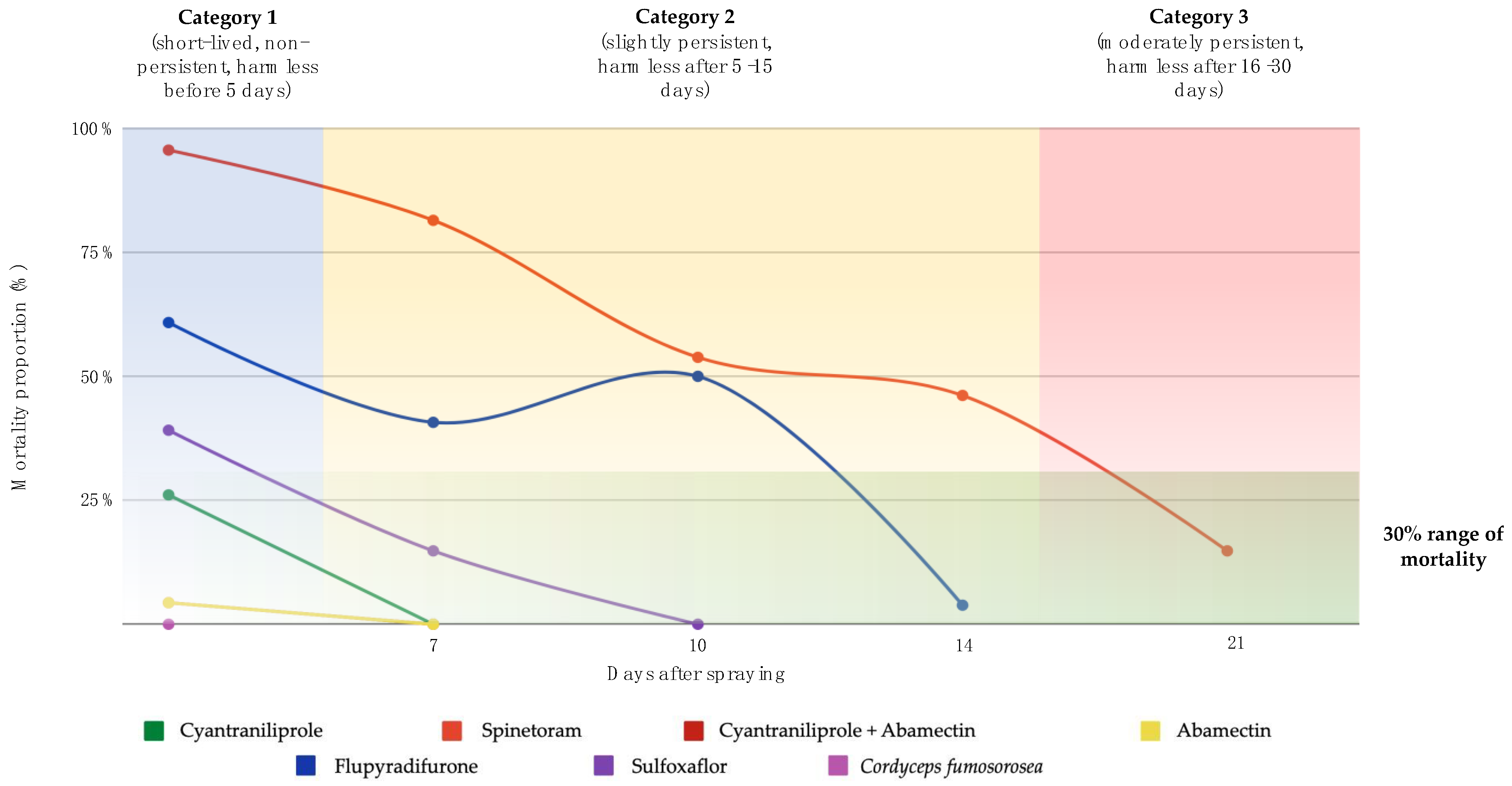
3.3. Residual Effects (Persistence) of Insecticides on T. atopovirilia
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slone, J.D.; Burrack, H.J. Integrated Pest Management Practices Reduce Insecticide Applications, Preserve Beneficial Insects, and Decrease Pesticide Residues in Flue-Cured Tobacco Production. J. Econ. Entomol. 2016, 109, 2397–2404. [Google Scholar] [CrossRef]
- Rakes, M.; Pasini, R.A.; Morais, M.C.; Araújo, M.B.; de Bastos Pazini, J.; Seidel, E.J.; Bernardi, D.; Grützmacher, A.D. Pesticide Selectivity to the Parasitoid Trichogramma pretiosum: A Pattern 10-Year Database and Its Implications for Integrated Pest Management. Ecotoxicol. Environ. Saf. 2021, 208, 111504. [Google Scholar] [CrossRef] [PubMed]
- Bueno, A.d.F.; Carvalho, G.A.; dos Santos, A.C.; Sosa-Gómez, D.R.; da Silva, D.M. Pesticide Selectivity to Natural Enemies: Challenges and Constraints for Research and Field Recommendation. Ciência Rural. 2017, 47, e20160829. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Coelho, A. Insect Rearing Techniques for Biological Control Programs, a Component of Sustainable Agriculture in Brazil. Insects 2022, 13, 105. [Google Scholar] [CrossRef] [PubMed]
- Bull, D.L.; House, V.S. Effects of Different Insecticides on Parasitism of Host Eggs by Trichogramma pretiosum Riley. Southw. Entomol. 1983, 1, 46–53. [Google Scholar]
- Jacobs, R.J.; Kouskolekas, C.A.; Gross Júnior, H.R. Responses of Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) to Residues of Permethin and Endosulfan. Environ. Entomol. 1984, 2, 355–358. [Google Scholar] [CrossRef]
- Brunner, J.F.; Dunley, J.E.; Doerr, M.D.; Beers, E.H. Effect of Pesticides on Colpoclypeus florus (Hymenoptera: Eulophidae) and Trichogramma platneri (Hymenoptera: Trichogrammatidae), Parasitoids of Leafrollers in Washington. J. Econ. Entomol. 2001, 94, 1075–1084. [Google Scholar] [CrossRef]
- Parra, J.R.P.; Bento, J.M.S.; Garcia, M.S.; Yamamoto, P.T.; Vilela, E.F.; Leal, W.S. Development of a Control Alternative for the Citrus Fruit Borer, Ecdytolopha aurantiana (Lepidoptera, Tortricidae): From Basic Research to the Grower. Rev. Bras. Entomol. 2004, 48, 561–567. [Google Scholar] [CrossRef]
- Bento, J.M.S.; Parra, J.R.P.; De Miranda, S.H.G.; Adami, A.C.O.; Vilela, E.F.; Leal, W.S. How Much Is a Pheromone Worth? F1000Research 2016, 5, 1763. [Google Scholar] [CrossRef]
- Bento, J.M.S. Manual de Bicho-Furão: Medidas Essenciais de Controle; Bento, J.M., Parra, J.R.P., Yamamoto, P.T., Eds.; Fundecitrus: Araraquara, Brazil, 2019. [Google Scholar]
- Molina, R.M.S.; Fronza, V.; Parra, J.R.P. Seleção de Trichogramma spp., Para o Controle de Ecdytolopha aurantiana, Com Base Na Biologia e Exigências Térmicas. Rev. Bras. Entomol. 2005, 49, 152–158. [Google Scholar] [CrossRef]
- Molina, R.M.S. Bioecologia de Duas Espécies de Trichogramma Para o Controle de Ecdytolopha aurantiana (Lima, 1927) (Lepidoptera: Tortricidae) Em Citros. Dissertação (Mestrado em Entomologia). Master’s Thesis, ESALQ/USP, Piracicaba, Brazil, 2003. [Google Scholar]
- Gómez-Torres, M.L. Controle Biológico de Ecdytolopha aurantiana (Lima, 1927) (Lepidoptera: Tortricidae) Com Trichogramma atopovirilia Oatman & Platner, 1983. Dissertação (Mestrado em Entomologia). Master’s Thesis, ESALQ/USP, Piracicaba, Brazil, 2005. [Google Scholar]
- Garcia, M.S. Bioecologia e Potencial de Controle Biológico de Eclytolopha aurantianum (LIMA, 1927) (Lepidoptera: Tortricidae), o Bicho-Furão-Dos-Citros, Através de Trichogramma pretiosum Rley, 1879. Tese (Doutorado em Entomologia). Ph.D. Thesis, ESALQ/USP, Piracicaba, Brazil, 1998. [Google Scholar]
- Greene, G.L.; Leppla, N.C. DWA Velvet Bean Caterpillar: A Rearing Procedure and Artificial Medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- Garcia, M.S.; Parra, J.R.P. Comparação de Dietas Artificiais, Com Fontes Protéicas Variáveis, Para Criação de Ecdytolopha aurantiana (Lima) (Lepidoptera: Tortricidae). An. Soc. Entomol. Bras. 1999, 28, 219–232. [Google Scholar] [CrossRef]
- Camarozano, C.T.; Coelho, A.; da Silva, R.B.Q.; Parra, J.R.P. Can Trichogramma atopovirilia Oatman & Platner Replaces Trichogramma galloi Zucchi for Diatraea saccharalis (Fabricius) Control? Sci. Agric. 2021, 79. [Google Scholar] [CrossRef]
- Asrar, M.; Ali, S.; Mushtaq, N.; Zia, K.; Bibi, R.; Akhtar, I.; Hussain, D.; Li, Y.; Niaz, Y.; Alwahibi, M.S.; et al. Toxicity of Neem Seed Extract and Different Insecticides on Trichogramma chilonis (Hymenoptera: Trichogrammatidae). J. King Saud. Univ. Sci. 2022, 34, 102135. [Google Scholar] [CrossRef]
- Cônsoli, L.F.; Parra, J.R.; Vinson, S.B. Estimating Parasitoid Immature Mortality by Comparing Oviposition and Pupal Development of Trichogramma galloi Zucchi and T. pretiosum Riley on Natural and Factitious Hosts. Rev. Bras. Biol. 2000, 60, 381–387. [Google Scholar] [CrossRef]
- Bowen, W.R.; Stern, V.M. Effect of Temperature on the Production of Males and Sexual Mosaics in a Uniparental Race of Trichogramma semifumatum (Hymenoptera: Trichogrammatidae). Ann. Entomol. Soc. Am. 1996, 59, 823–834. [Google Scholar] [CrossRef]
- Hassan, S.A. Métodos Padronizados Para Testes de Seletividade Com Ênfase Em Trichogramma. In Trichogramma e o Controle Biológico Aplicado; Parra, J.R.P., Zucchi, R.A., Eds.; Fealq: Piracicaba, Brazil, 1997; pp. 207–233. [Google Scholar]
- Fagundes Matioli, T.; Zanuzo Zanardi, O.; Takao Yamamoto, P. Impacts of Seven Insecticides on Cotesia flavipes (Cameron) (Hymenoptera: Braconidae). Ecotoxicology 2019, 28, 1210–1219. [Google Scholar] [CrossRef]
- Ramírez-Cerón, D.; Rodríguez-Leyva, E.; Lomeli-Flores, J.R.; Soto-Rojas, L.; Ramírez-Alarcón, S.; Segura-Miranda, A. Toxicity and Residual Activity of Insecticides against Diadegma insulare, a Parasitoid of the Diamondback Moth. Insects 2022, 13, 514. [Google Scholar] [CrossRef]
- Hassan, S.A.; Bigler, F.; Bogenschütz, H.; Boller, E.; Brun, J.; Calis, J.N.M.; Coremans-Pelseneer, J.; Duso, C.; Grove, A.; Heimbach, U.; et al. Results of the Sixth Joint Pesticide Testing Programme of the IOBC/WPRS-Working Group «pesticides and Beneficial Organisms». Entomophaga 1994, 39, 107–119. [Google Scholar] [CrossRef]
- Takahashi, T.A. Seletividade de Inseticidas a Trichogramma atopovirilia e Trichogramma pretiosum (Hymenoptera: Trichogrammatidae) em Ovos de Dois Hospedeiros Naturais. Ph.D. Thesis, Universidade Federal do Paraná, Curitiba, Brazil, 2016. [Google Scholar]
- Khan, M.A. Lethal and Parasitism Effects of Selected Novel Pesticides on Adult Trichogramma chilonis (Hymenoptera: Trichogrammatidae). J. Plant Dis. Prot. 2020, 127, 81–90. [Google Scholar] [CrossRef]
- Sr, J.L. Insect Physiology and Biochemistry, 4th ed.; CRC Press: Boca Raton, FL, USA, 2022. [Google Scholar]
- Williams, L.; Price, L.D. A Space-Efficient Contact Toxicity Bioassay for Minute Hymenoptera, Used to Test the Effects of Novel and Conventional Insecticides on the Egg Parasitoids Anaphes Iole and Trichogramma pretiosum. BioControl 2004, 49, 163–185. [Google Scholar] [CrossRef]
- Nidagundi, K.S.; Mallapur, C.P. Contact Toxicity of Insecticides and Biopesticides to Trichogramma chilonis Ishii. (Hymenoptera: Trichogrammatidae) under Laboratory Condition. Res. Sq. 2021, 1–16. [Google Scholar] [CrossRef]
- Tabebordbar, F.; Shishehbor, P.; Ziaee, M.; Sohrabi, F. Lethal and Sublethal Effects of Two New Insecticides Spirotetramat and Flupyradifurone in Comparison to Conventional Insecticide Deltamethrin on Trichogramma evanescens (Hymenoptera: Trichogrammatidae). J. Asia Pac. Entomol. 2020, 23, 1114–1119. [Google Scholar] [CrossRef]
- Iost Filho, F.H.; Paiva, A.C.R.; Yamamoto, P.T. Flupyradifurone Is Selective to the Egg’s Parasitoid Trichogramma pretiosum. In Proceedings of the ESA 2017 Annual Meeting, Denver, CO, USA, 5–8 November 2017. [Google Scholar]
- Costa, M.A.; Farias, E.S.; Passos, L.C.; Carvalho, V.C.; Carvalho, G.A. Side Effects of Insecticides Applied to Cotton on Adult Trichogramma pretiosum by Three Exposure Routes. Pest. Manag. Sci. 2022, 78, 1895–1902. [Google Scholar] [CrossRef]
- Harwood, G.P.; Prayugo, V.; Dolezal, A.G. Butenolide Insecticide Flupyradifurone Affects Honey Bee Worker Antiviral Immunity and Survival. Front. Insect Sci. 2022, 2, 907555. [Google Scholar] [CrossRef]
- Pratissoli, D.; Milanez, A.M.; Celestino, F.N.; Barbosa, W.F. Trichogramma atopovirilia Oatman & Platner (Hymenoptera: Trichogrammatidae) Em Condições de Laboratório. Rev. Ceres 2011, 58, 661–664. [Google Scholar]
- Soares, M.A.; Leão, G.; Leite, D.; Zanuncio, J.C.; Mendes De Sá, V.G.; Soares Ferreira, C.; Rocha, S.L.; Pires, M.; Serrão, J.E. Quality Control of Trichogramma atopovirilia and Trichogramma pretiosum (Hym.: Trichogrammatidae) Adults Reared under Laboratory Conditions. Arch. Biol. Technol. 2012, 55, 305–311. [Google Scholar] [CrossRef]
- Delpuech, J.M.; Meyet, J. Reduction in the Sex Ratio of the Progeny of a Parasitoid Wasp (Trichogramma brassicae) Surviving the Insecticide Chlorpyrifos. Arch. Environ. Contam. Toxicol. 2003, 45, 203–208. [Google Scholar] [CrossRef]
- van Emden, H.F.; Sponagl, B.; Wagner, E.; Baker, T.; Ganguly, S.; Douloumpaka, S. Hopkins’ ‘Host Selection Principle’, Another Nail in Its Coffin. Phv. Entomol. 1996, 21, 325–328. [Google Scholar] [CrossRef]
- Salgado, V.L.; Sparks, T.C. The Spinosyns: Chemistry, Biochemistry, Mode of Action, and Resistance. In Comprehensive Molecular Insect Science; Gilbert, L.I., Latrou, K., Gill, S.S., Eds.; Elsevier: Oxford, UK, 2005; pp. 137–173. [Google Scholar]


{kind=link}
| Active Ingredient (a.i.) | Trade Name, Formulation | Concentration (g a.i. L−1) | c.a.i. 1 | Toxicology Group | Mode of Action |
|---|---|---|---|---|---|
| Cyantraniliprole + abamectin | Minecto Pro ®, SC | 60 + 18 | 0.40 | Diamide +Ivermectin | Ryanodine receptor modulators. Act on the nervous system. Glutamate-gated chloride channel allosteric modulators. Acts on the nervous system and muscle system. |
| Abamectin | Batent ®, EC | 18 | 0.60 | Ivermectin | Glutamate-gated chloride channel allosteric modulators. Acts on the nervous system and muscle system. |
| Spinetoram | Delegate ®, WG | 250 | 0.25 | Spinosyns | Nicotinic acetylcholine receptor allosteric modulators. Acts on the central nervous system. |
| Sulfoxaflor | Verter ®, SC | 240 | 0.60 | Sulfoxamine | Competitive modulators of nicotinic acetylcholine receptors. Act on the nervous system. |
| Cyantraniliprole | Benevia ®, OD | 100 | 0.75 | Diamide | Ryanodine receptor modulators. Act on the nervous system. |
| Flupyradifurone | Sivanto ®, SL | 200 | 0.80 | Butenolide | Competitive modulators of nicotinic acetylcholine receptors. Act on the nervous system. |
| Cordyceps fumosorosea * | Challenger ®, SC | 85 | 0.10 | Entomopathogen fungi | Attachment of hyphae or spores. Penetration and internal colonization in the insect body |
| Treatment | Adult Mortality (%) | 24 h Parasitism (G. aurantianum Treated Eggs) 1 | Parasitism Reduction (%) | IOBC/WPRS Classification 2 | Lifespan Parasitism 1 | Longevity (Days) 1 |
|---|---|---|---|---|---|---|
| Control | 3.33 ± 3.33 a | 8.66 ± 1.15 a | -- | -- | 56.00 ± 8.33 a | 8.03 ± 0.93 a |
| Cyantraniliprole + abamectin | 10.00 ± 5.57 a | 7.60 ± 0.85 a | 45.53 | 2 | 30.50 ± 4.60 abc | 6.36 ± 0.92 bc |
| Abamectin | 10.00 ± 5.57 a | 3.71 ± 0.66 b | 53.81 | 2 | 25.86 ± 5.15 bc | 4.26 ± 0.57 c |
| Spinetoram | 46.67 ± 9.24 b | 3.79 ± 0.49 b | 77.31 | 2 | 12.70 ± 6.39 c | 2.93 ± 0.72 d |
| Sulfoxaflor | 10.00 ± 4.63 a | 5.26 ± 1.02 ab | 0 | 1 | 60.16 ± 7.86 a | 8.47 ± 1.09 a |
| Cyantraniliprole | 3.33 ± 3.33 a | 9.36 ± 1.27 a | 0 | 1 | 62.60 ± 7.11 a | 9.20 ±1.00 a |
| Flupyradifurone | 0.00 ± 0.00 a | 6.33 ± 1.00 ab | 27.62 | 1 | 40.53 ± 4.70 ab | 7.20 ± 0.71 ab |
| Cordyceps fumosorosea | 10.00 ± 5.57 a | 6.50 ± 0.79 ab | 3.57 | 1 | 54.00 ± 8.28 ab | 8.50 ± 0.75 a |
| χ2 | 36.56 | 26.99 | 49.91 | 54.62 | ||
| p | <0.001 | <0.001 | <0.001 | <0.001 |
| Treatment | Emergence (%) 2 | Sex Ratio 2 | Longevity (Days) 1 | Parasitism (72 h) 1 | F2 Emergence (%) 2,3 |
|---|---|---|---|---|---|
| Control | 42.42 ± 7.00 ab | 0.70 ± 0.04 a | 5.41 ± 0.28 a | 35.81 ± 5.49 ns | 95.22 ± 0.03 a |
| Cyantraniliprole + Abamectin | 46.16 ± 6.37 ab | 0.28 ± 0.03 b | 3.20 ± 0.48 b | 21.00 ± 8.15 | 86.37 ± 0.09 ab |
| Abamectin | 33.10 ± 10.59 b | 0.57 ± 0.13 a | 3.44 ± 0.24 b | 38.55 ± 6.07 | 94.09 ± 0.02 ab |
| Spinetoram | 12.03 ± 4.86 c | 0.73± 0.14 a | 3.84 ± 0.42 ab | 27.50 ± 7.44 | 82.12 ± 0.16 b |
| Sulfoxaflor | 43.76 ± 7.65 ab | 0.63 ± 0.05 a | 2.50 ± 0.22 b | 27.33 ± 7.44 | 85.46 ± 0.03 b |
| Cyantraniliprole | 28.82 ± 5.79 bc | 0.21 ± 0.05 b | 3.28 ± 0.47 b | 36.85 ± 6.89 | 95.47 ± 0.01 ab |
| Flupyradifurone | 45.17 ± 7.65 ab | 0.68 ± 0.06 a | 3.60 ± 0.24 ab | 46.80 ± 8.15 | 91.56 ± 0.02 ab |
| Cordyceps fumosorosea | 46.16 ± 6.37 a | 0.63 ± 0.10 a | 4.25 ± 0.52 ab | 36.33 ± 5.51 | 94.58 ± 0.01 a |
| χ2 | 27.17 | 99.89 | 24.85 | 9.57 | 25.83 |
| p | <0.001 | <0.001 | <0.001 | 0.2146 | <0.001 |
| Treatment | Emergence (%) 1 | Parasitism Reduction (%) 2 | IOBC/WPRS Classification 3 |
|---|---|---|---|
| Control | 76.50 ± 2.67 ab | -- | -- |
| Cyantraniliprole + abamectin | 74.81 ± 2.78 b | 2.75 | 1 |
| Abamectin | 77.82 ± 2.62 ab | 0.00 | 1 |
| Spinetoram | 5.42 ± 1.36 c | 92.92 | 3 |
| Sulfoxaflor | 80.22 ± 3.15 ab | 0.00 | 1 |
| Cyantraniliprole | 74.37 ± 4.10 b | 3.21 | 1 |
| Flupyradifurone | 84.28 ± 1.95 a | 0.00 | 1 |
| Cordyceps fumosorosea | 74.57 ± 2.14 ab | 2.20 | 1 |
| χ2 | 79.75 | - | - |
| p | <0.0001 | - | - |
| Treatment | Day 3 | Day 7 | Day 10 | Day 14 | Day 21 | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| P 1 | E (%) 2 | P 1 | E (%) 2 | P 1 | E (%) 2 | P 1 | E (%) 2 | P 1 | E (%) 2 | |
| Control | 22.36 ± 4.11 ab | 97.20 ± 1.14 a | 58.84 ± 3.44 a | 98.65 ± 0.42 a | 14.45 ± 3.88 b | 99.93 ± 0.06 a | 18.23 ± 3.25 ab | 100 ± 0.00 a | 12.36 ± 1.85 | 100 ± 0.00 a |
| Cyantraniliprole + abamectin | 24.55 ± 4.07 a | 93.28 ± 2.91 a | ||||||||
| Abamectin | 9.16 ± 3.74 b | 91.92 ± 5.52 a | ||||||||
| Spinetoram | * | * | * | * | * | * | 23.00 ± 5.04 a | 95.45 ± 3.24 a | 7.81 ± 2.19 ns | 84.68 ± 4.57 b |
| Sulfoxaflor | 14.67 ± 6.68 ab | 88.88 ± 11.11 b | 48.17 ± 4.85 ab | 93.42 ± 4.25 b | ||||||
| Cyantraniliprole | 26.71 ± 5.68 a | 94.86 ± 5.68 a | ||||||||
| Flupyradifurone | 33.56 ± 7.72 a | 98.36 ± 0.86 a | 35.56 ± 5.61 b | 96.32 ± 0.75 b | 35.56 ± 5.61 a | 96.33 ± 0.75 b | 9.96 ± 2.72 b | 99.27 ± 0.72 a | ||
| Cordyceps fumosorosea | 31.71 ± 5.29 a | 98.09 ± 0.57 a | ||||||||
| χ2 | 21.46 | 69.55 | 10.72 | 18.52 | 10.14 | 18.65 | 8.39 | 3.89 | 1.98 | 20.53 |
| p | 0.0015 | 0.008 | 0.004 | <0.001 | 0.0014 | <0.001 | 0.0151 | 0.35 | 0.15 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cantori, L.V.; Iost Filho, F.H.; Pazini, J.d.B.; Diniz, A.J.F.; Yamamoto, P.T.; Parra, J.R.P. Is Integrated Management of Gymnandrosoma aurantianum Possible with Trichogramma atopovirilia and Novel Products Used in Citrus Orchards in Brazil? Insects 2023, 14, 419. https://doi.org/10.3390/insects14050419
Cantori LV, Iost Filho FH, Pazini JdB, Diniz AJF, Yamamoto PT, Parra JRP. Is Integrated Management of Gymnandrosoma aurantianum Possible with Trichogramma atopovirilia and Novel Products Used in Citrus Orchards in Brazil? Insects. 2023; 14(5):419. https://doi.org/10.3390/insects14050419
Chicago/Turabian StyleCantori, Lucas Vinicius, Fernando Henrique Iost Filho, Juliano de Bastos Pazini, Alexandre José Ferreira Diniz, Pedro Takao Yamamoto, and José Roberto Postali Parra. 2023. "Is Integrated Management of Gymnandrosoma aurantianum Possible with Trichogramma atopovirilia and Novel Products Used in Citrus Orchards in Brazil?" Insects 14, no. 5: 419. https://doi.org/10.3390/insects14050419