

A Review of the Host Plant Location and Recognition Mechanisms of Asian Longhorn Beetle
Abstract
:Simple Summary
Abstract
1. Introduction
2. Host Plant List and Host Kairomones
2.1. Host Plant Lists
2.2. Host Plant Kairomones
3. Interaction of ALBs, Microbes and Host Plants
3.1. Odorant-Binding Proteins of ALB to Recognize Host Plants
3.2. Collaboration with Microbes to Degrade Host Plant Tissue
4. Practical Applications of Host Plants and Host Kairomones
4.1. Mixed Forest
4.2. Trapping Technique
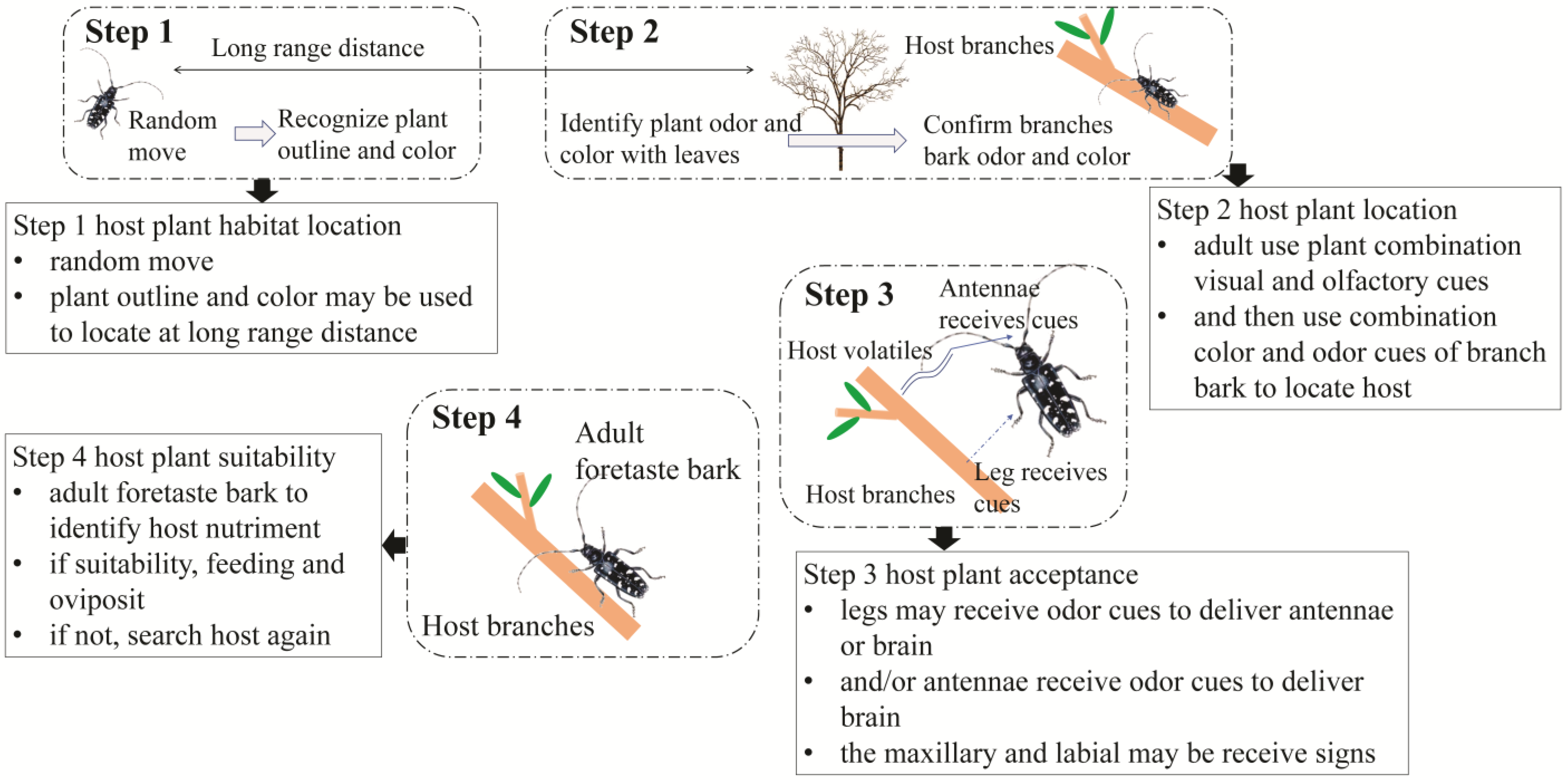
5. Host Plant Location and Recognition Behavior Hypothesis
6. Further Research Directions on Interaction of ALBs and Host Plants
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Elisabeth, C.B.; Erik, G.B. Ecosystem services from urban forests: The case of Oslomarka, Norway. Ecosyst. Serv. 2021, 51, 101358. [Google Scholar] [CrossRef]
- Allison, J.D.; Redak, R.A. The impact of trap type and design features on survey and detection of bark and woodboring beetles and their associates: A review and meta-analysis. Annu. Rev. Entomol. 2017, 62, 127–146. [Google Scholar] [CrossRef]
- Branco, S.; Faccoli, M.; Brockerhoff, E.G.; Roux, G.; Jactel, H.; Desneux, N.; Gachet, E.; Mouttet, R.; Streito, J.C.; Branco, M. Preventing invasions of Asian longhorn beetle and citrus longhorn beetle: Are we on the right track? J. Pest. Sci. 2022, 95, 41–66. [Google Scholar] [CrossRef]
- Pedlar, J.H.; Mckenney, D.W.; Denys, Y.; Hope, E.S. Potential economic impacts of the Asian Longhorned Beetle (Coleoptera: Cerambycidae) in eastern Canada. J. Econ. Entomol. 2019, 113, 839–850. [Google Scholar] [CrossRef] [PubMed]
- Haack, R.A.; Herard, F.; Sun, J.; Turgeon, J.J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu. Rev. Entomol. 2010, 55, 521–546. [Google Scholar] [CrossRef] [Green Version]
- Van der Gaag, D.J.; Loomans, A.J.M. Host plants of Anoplophora glabripennis, a review. EPPO Bull. 2014, 44, 518–528. [Google Scholar] [CrossRef]
- Brabbs, T.; Collins, D.; Herard, F.; Maspero, M.; Eyre, D. Prospects for the use of biological control agents against Anoplophora in Europe. Pest Manag. Sci. 2015, 71, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.J.; Hajek, A.E. Thermoregulatory behavior and fungal infection of Anoplophora glabripennis (Coleoptera: Cerambycidae). Environ. Entomol. 2014, 43, 384–392. [Google Scholar] [CrossRef]
- Clifton, E.H.; Jaronski, S.T.; Hajek, A.E. Virulence of commercialized fungal entomopathogens against Asian Longhorned Beetle (Coleoptera: Cerambycidae). J. Insect Sci. 2020, 20, 1. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.Y.; Kenis, M.; Cao, L.M.; Hoelmer, K.A. Exploring the potential for novel associations of generalist parasitoids for biological control of invasive woodboring beetles. BioControl 2021, 66, 97–112. [Google Scholar] [CrossRef]
- Kaitlin, R.; Golec, J.R.; Duan, J.J. Host selection and potential non-target risk of Dastarcus helophoroides, a larval parasitoid of the Asian longhorned beetle, Anoplophora glabripennis. Biol. Control 2018, 123, 120–126. [Google Scholar] [CrossRef]
- Beketov, M.A.; Kefford, B.J.; Schafer, R.B.; Liess, M. Pesticides reduce regional biodiversity of stream invertebrates. Proc. Natl. Acad. Sci. USA 2013, 110, 11039–11043. [Google Scholar] [CrossRef] [Green Version]
- Basset, Y.; Lamarre, G. Toward a world that values insects. Science 2019, 364, 1230–1231. [Google Scholar] [CrossRef] [Green Version]
- Golec, J.R.; Li, F.; Cao, L.; Wang, X.; Duan, J.J. Mortality factors of Anoplophora glabripennis (Coleoptera: Cerambycidae) infesting Salix and Populus in central, northwest, and northeast China. Biol. Control 2018, 126, 198–208. [Google Scholar] [CrossRef]
- Hanks, L.M. Influence of the larval host plant on reproductive strategies of Cerambycid beetles. Annu. Rev. Entomol. 1999, 44, 483–505. [Google Scholar] [CrossRef] [Green Version]
- Morewood, W.D.; Hoover, K.; Neiner, P.R.; Mcneil, J.R.; Sellmer, J.C. Host tree resistance against the polyphagous wood-boring beetle Anoplophora glabripennis. Entomol. Exp. Appl. 2004, 110, 79–86. [Google Scholar] [CrossRef]
- Morewood, W.D.; Neiner, P.R.; Mcneil, J.R.; Sellmer, J.C.; Hoover, K. Oviposition preference and larval performance of Anoplophora glabripennis (Coleoptera: Cerambycidae) in four eastern north American hardwood tree species. Environ. Entomol. 2003, 32, 1028–1034. [Google Scholar] [CrossRef]
- Faccoli, M.; Favaro, R. Host preference and host colonization of the Asian long-horned beetle, Anoplophora glabripennis (Coleoptera Cerambycidae), in Southern Europe. Bull Entomol. Res. 2016, 106, 359–367. [Google Scholar] [CrossRef]
- Mitchell, R.F.; Hall, L.P.; Reagel, P.F.; McKenna, D.D.; Baker, T.C.; Hildebrand, J.G. Odorant receptors and antennal lobe morphology offer a new approach to understanding olfaction in the Asian longhorned beetle. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 2017, 203, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyu, F.; Hai, X.; Wang, Z. Green-colored paperboard enhances the Asian longhorned beetle response to host plant odor cues. J. Pest Sci. 2021, 94, 1345–1355. [Google Scholar] [CrossRef]
- Wang, Z.G.; Huang, D.Z.; Yan, J.J. The sequence choice of the main tree species to Anoplophora glabripennis in northern part of China. J. Agric. Univ. Hebei 2009, 32, 62–68. [Google Scholar] [CrossRef]
- Straw, N.A.; Fielding, N.J.; Tilbury, C.; Williams, D.T.; Inward, D. Host plant selection and resource utilisation by Asian longhorn beetle Anoplophora glabripennis (Coleoptera: Cerambycidae) in southern England. Forestry 2015, 88, 84–95. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.Z.; Yang, X.Y.; Wei, J.N.; Lang, X.R. An Investigation on the resistance of major forestation species to Anoplophora glabripennis and A. nobilis. J. Northwest For. Coll. 1997, 12, 42–46. [Google Scholar]
- Chen, B.Y.; Liao, Y.; Zhu, L. The occurrence and prevention and control techniques discussion of Anoplophora glabropennis on Camphor trees in nursery. China Plant Prot. 2008, 28, 30–31. [Google Scholar]
- Zhang, F.; Jin, Y.; Chen, H.; Wu, X. The selectivity mechanism of Anoplphora glabripennis on four different species of maples. Acta Ecol. Sin. 2006, 26, 870–877. [Google Scholar]
- Meng, P.S.; Hoover, K.; Keena, M.A. Asian Longhorned Beetle (Coleoptera: Cerambycidae), an introduced pest of Maple and other hardwood trees in North America and Europe. J. Integr. Pest Manag. 2015, 6, 84–88. [Google Scholar] [CrossRef]
- Wang, Z.; Yuan, K.Y.; Wang, X.M. Investigation of damages and annual life history of Anoplophora glabripennis in Baotou in the Inner Mongolia autonomous region. J. Inn. Mong. Agric. Univ. 2016, 37, 87–95. [Google Scholar]
- An, Y.L. The occurrence of Asian longhorned beetle and its analysis in USA. China Commod. Insp. 1999, 2, 9–10. [Google Scholar]
- Wang, F.; Zhou, J.; Yang, X. Study on the relationship between the behavior of selecting host of Anoplophora nobilis and host resistance on this beetle in mixed forest. Sci. Silvae Sin. 2000, 36, 58–65. [Google Scholar]
- Lim, J.; Jung, S.Y.; Lim, J.S.; Jang, J.; Kim, K.M.; Lee, Y.M.; Lee, B.W. A review of host plants of Cerambycidae (Coleoptera: Chrysomeloidea) with new host records for fourteen Cerambycids, including the Asian longhorn beetle (Anoplophora glabripennis Motschulsky), in Korea. Korean J. Appl. Entomol. 2014, 53, 111–133. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z. The Occurrence Regulation of Anoplophora glabripennis in the Harbin. Master’s Thesis, University of Heilongjiang, Haerbing, China, 2010. [Google Scholar]
- Jiang, W.N.; Liu, J.F.; Dong, X.W. Distribution and damage characteristics of Anoplophora glabripennis in Heilongjiang Province. Prot. For. Sci. Technol. 2012, 4, 71–77. [Google Scholar]
- Yang, X.Y.; Yan, X.H.; Zhou, J.X. Resistance of populus spp. to Anoplophora glabripennis. J. Northwest For. Coll. 1991, 6, 30–38. [Google Scholar]
- Hu, J.; Angeli, S.; Schuetz, S.; Luo, Y.; Hajek, A.E. Ecology and management of exotic and endemic Asian longhorned beetle Anoplophora glabripennis. Agric. For. Entomol. 2009, 11, 359–375. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, D.; Wang, Z.; Yan, Y.; Yin, J. Effects of secondary metabolites of poplar trees on the carboxylesterase and glutathione-S-transferase in Anoplophora glabripennis. Sci. Silvae Sin. 2001, 37, 123–128. [Google Scholar]
- Mu, Z.; Yin, R.; Qi, C.; Zhang, R.; Yu, J. A research for the occurrence of Anoplophora glabripennis Motschulsky in the poplar intensive management plantation. Sci. Silvae Sin. 1999, 35, 148–152. [Google Scholar]
- Tang, H.; Han, Y.; Ru, J.; Qu, M.; Wang, X. Evaluation of control effects on different measures against Anoplophora glabripennis by life table method. J. Northwest For. Univ. 2020, 35, 101–107. [Google Scholar]
- Wang, X.; Yuan, C.; Wang, J.; Fan, D.; Chu, X.; Jie, F.; An, B.; Ning, L. Study on the mechanism of inhibiting the hatching of eggs of Anoplophora glabropennis by I-69 Populus. In Proceedings of the Annual Meeting of Entomological Society of China, Yichang, China; 2000; pp. 884–885. [Google Scholar]
- Wang, Z.; Yan, J.; Liu, Y.; Tang, X.; Chen, Y. Investigation of Anoplophora glabripennis in Southern Tibet. J. Northeast For. Univ. 2003, 31, 70–71. [Google Scholar]
- Zhao, X.; Zhou, Z.; Guo, C.; Li, H.; Guo, S. Relationship between tannin content of willows poplar and damage of Anoplophora glabripennis. For. Investig. Des. 2011, 157, 73–75. [Google Scholar]
- Zhu, N.; Zhang, D.Y.; Shen, H.X.; Hu, Q.; Fan, J.T. Oviposition preferences of Anoplophora glabripennis on the three host plants and composition analysis of host plant volatiles. J. Zhejiang Agric. For. Univ. 2017, 34, 1059–1064. [Google Scholar]
- Ma, W.; Zhou, Y.; Liu, Y.; Meng, Z.; Zhang, A.; Yan, S. Trapping efficacy of traps baited with four types of attractants for population evaluation of Anoplophora glabripennis. J. Northeast For. Univ. 2021, 49, 120–130. [Google Scholar] [CrossRef]
- Yan, X.F.; Liu, Y.H. Host plant effects on weight, longevity, fecundity and hatching rate of Anoplophora glabripennis. Chin. Agric. Sci. Bull. 2012, 28, 52–56. [Google Scholar]
- Fan, L.; Yan, S.; Sun, Z.; Meng, Z. EAG and behavioral responses of Asian longhorn beetle Anoplophora glabripennis (Coleoptera: Cerambycidae) to plant volatiles. Chin. J. Ecol. 2013, 32, 142–148. [Google Scholar] [CrossRef]
- Bahatiguli, M. Investigation on the control of Asian longhorn beetle in Xinjiang by cutting tree crown. Prot. For. Sci. Technol. 2016, 154, 100–123. [Google Scholar] [CrossRef]
- Gao, R.; Wang, B.; Mastro, V.; Li, Y.; Wang, Y.; Yang, X. Infestation of Betula platyphylla by Anoplophora glabripennis and its control using insecticides. Sci. Silvae Sin. 2009, 45, 163–166. [Google Scholar]
- Qin, K. Study on the Bionomics and Control of Technology of Anoplophora glabripennis in Harbin. Master’s thesis, Northeast Forestry University, Harbin, China, 2009. [Google Scholar]
- Du, H.; Wang, P.; Xu, H.; Zhang, W.; Wang, Z. EAG responses of Asian longhorn beetle Anoplophora glabripennis (Coleoptera: Cerambycidae) to volatiles of hickory. J. Zhejiang Agric. Univ. 2016, 33, 166–171. [Google Scholar]
- Qiao, H.L.; Luo, Y.Q.; Feng, X.; Sun, J.H.; Han, X.W. The resistance of the main host-tree species to Anoplophora glabripennis in Xinjiang. Chin. Bull. Entomol. 2007, 44, 660–664. [Google Scholar]
- Turgeon, J.J.; Chuck, J.; Smith, M.T.; Mary, O.; Scarr, T.A.; Ben, G. Records of unsuccessful attack by Anoplophora glabripennis (Coleoptera: Cerambycidae) on broadleaf trees of questionable suitability in Canada. Can. Entomol. 2016, 148, 569–578. [Google Scholar] [CrossRef]
- Gu, Q.; Jia, R.; Li, H.; Guo, S.; Xu, T.; Lu, P.; Qiao, H. Host adaptability of Anoplophora glabripennis based on metabolomics and insect intestinal bacteria. J. Plant Prot. 2022, 49, 1750–1763. [Google Scholar] [CrossRef]
- Bancroft, J.S.; Smith, M.T.; Tropp, C.J. Rapid test of the suitability of host-trees and the effects of larval history on Anoplophora glabripennis (Coleoptera: Cerambycidae). J. Kans. Entomol. Soc. 2002, 75, 308–316. [Google Scholar]
- Tian, R.M.; Lu, Q.; Zhang, Y.F.; Sang, H. Induce and control of Anoplophora glabripennis. J. Inn. Mong. For. Sci. Technol. 2009, 35, 42–46. [Google Scholar]
- Van der Kooi, C.J.; Stavenga, D.G.; Arikawa, K.; Belusic, G.; Kelber, A. Evolution of insect color vision: From spectral sensitivity to visual ecology. Annu. Rev. Entomol. 2021, 66, 435–461. [Google Scholar] [CrossRef]
- Warrant, E.; Dacke, M. Vision and visual navigation in nocturnal insects. Annu. Rev. Entomol. 2011, 56, 239–254. [Google Scholar] [CrossRef]
- Hall, L.P.; Graves, F.; Myrick, A.; Hoover, K.; Baker, T.C. Labial and maxillary palp recordings of the Asian longhorned beetle, Anoplophora glabripennis, reveal olfactory and hygroreceptive capabilities. J. Insect. Physiol. 2019, 117, 103905. [Google Scholar] [CrossRef]
- Zapponi, L.; Nieri, R.; Zaffaroni-Caorsi, V.; Pugno, N.M.; Mazzoni, V. Vibrational calling signals improve the efficacy of pheromone traps to capture the brown marmorated stink bug. J. Pest Sci. 2022, 96, 587–597. [Google Scholar] [CrossRef]
- Zhu, E.Y.; Guntur, A.R.; He, R.; Stern, U.; Yang, C.H. Egg-laying demand induces aversion of UV light in Drosophila females. Curr. Biol. 2014, 24, 2797–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prokopy, R.J.; Owens, E.D. Visual detection of plants by herbivorous insects. Annu. Rev. Entomol. 1983, 28, 337–364. [Google Scholar] [CrossRef]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The use of push-pull strategies in integrated pest management. Annu. Rev. Entomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [Green Version]
- Peng, H.; Anne-Violette, L.; Cesar, R.-S.; Nicolas, D. Bottom-up forces in agroecosystems and their potential impact on arthropod pest management. Annu. Rev. Entomol. 2022, 67, 239–259. [Google Scholar] [CrossRef]
- Shih, P.Y.; Sugio, A.; Simon, J.C. Molecular mechanisms underlying host plant specificity in Aphids. Annu. Rev. Entomol. 2023, 68, 431–450. [Google Scholar] [CrossRef]
- Tumlinson, J.H. Complex and beautiful: Unraveling the intricate communication systems among plants and insects. Annu. Rev. Entomol. 2023, 68, 1–12. [Google Scholar] [CrossRef]
- Li, J.; Luo, Y.; Jin, Y. Electroantennogram activity of Ash-leaf maple (Acer negundo) volatiles to Anoplophora glabripennis (Motsch.). J. BeiJing For. Univ. 1999, 21, 1–5. [Google Scholar]
- Li, J.G.; Jin, Y.J.; Luo, Y.Q.; Xu, Z.C.; Chen, H.J. Leaf volatiles from host tree Acer negundo: Diurnal rhythm and behavior responses of Anoplophora glabripennis to volatiles in field. Acta Bot. Sin. 2003, 45, 177–182. [Google Scholar]
- Jin, Y.; Li, J.; Li, J.; Luo, Y.; Stephen, A.T. Olfactory response of Anoplophora glabripennis to volatile compounds from ash leaf maple (Acer negundo) under drought stress. Sci. Silvae Sin. 2004, 40, 99–105. [Google Scholar] [CrossRef]
- Wickham, J.D. Semiochemicals of the Asian Longhorned Beetle, Anoplophora glabripennis (Motschulsky), (Coleoptera: Cerambycidae). Ph.D. Thesis, State University of New York, New York, NY, USA, 2009. [Google Scholar]
- Nehme, M.E.; Keena, M.A.; Zhang, A.; Baker, T.C.; Hoover, K. Attraction of Anoplophora glabripennis to male-produced pheromone and plant volatiles. Environ. Entomol. 2009, 38, 1745–1755. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Gao, W.; Cheng, X.; Zhou, Y.; Cui, W. Electroantennogram response of Anoplophora glabripennis (Motsch.) to Acer negundo volatiles. For. Pest Dis. 2016, 35, 9–14. [Google Scholar]
- Tang, H.; Li, D.; Liu, Y.; Tokoro, M.; Nakashima, T. A preliminary study on the repellents of Anoplophora glabripennis. J. Beijing For. Univ. 1999, 1, 37–41. [Google Scholar]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Wang, J.; Gao, P.; Luo, Y.; Tao, J. Characterization and expression profiling of odorant-binding proteins in Anoplophora glabripennis Motsch. Gene 2019, 693, 25–36. [Google Scholar] [CrossRef]
- Andersson, M.N.; Grosse-Wilde, E.; Keeling, C.I.; Bengtsson, J.M.; Yuen, M.M.S.; Li, M.; Hillbur, Y.; Bohlmann, J.; Hansson, B.S.; Schlyter, F. Antennal transcriptome analysis of the chemosensory gene families in the tree killing bark beetles, Ips typographus and Dendroctonus ponderosae (Coleoptera: Curculionidae: Scolytinae). BMC Genom. 2013, 14, 198. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, D.Z.; Min, S.F.; Mi, F.; Zhou, S.S.; Wang, M.Q. Analysis of chemosensory gene families in the beetle Monochamus alternatus and its parasitoid Dastarcus helophoroides. Comp. Biochem. Physiol. Part D 2014, 11, 1–8. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, X.; Zhang, Y.; Zheng, Y.; Fan, Z.; Zhang, R. Identification of olfactory genes in Monochamus saltuarius and effects of Bursaphelenchus xylophilus infestation on their expression. Forests 2022, 13, 258. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Chen, Q.; Zhao, H.; Wang, J.; Wen, M.; Xi, J.; Ren, B. Identification and evolution of olfactory genes in the small poplar longhorn beetle Saperda populnea. Comp. Biochem. Physiol. Part D 2018, 26, 58–68. [Google Scholar] [CrossRef]
- Li, G.; Chen, X.; Shang, T. cDNA cloning, expression and ligand binding properties of the odorant binding protein AglaOBP12 in the Asian longhorned beetle, Anoplopha glabripennis (Coleoptera: Cerambycidae). Acta Entomol. Sin. 2017, 60, 1141–1154. [Google Scholar] [CrossRef]
- Li, G.; Chen, X.; Shang, T.; Yao, F. Cloning, expression, and binding properties of odorant binding protein 1 from Anoplophora glabripennis Motschulsky (Coleoptera: Cerambycidae). J. Environ. Entomol. 2017, 39, 919–929. [Google Scholar] [CrossRef]
- Wang, Q. Identification of Olfactory Related Genes and Functional Analysis of the Key Odorant Binding Proteins from Two Species of Anoplophora. Ph.D. Thesis, Beijing Forestry University, Beijing, China, 2019. [Google Scholar]
- Xu, T.; Hansen, L.; Cha, D.H.; Hao, D.; Zhang, L.; Teale, S.A. Identification of a female-produced pheromone in a destructive invasive species: Asian longhorn beetle, Anoplophora glabripennis. J. Pest Sci. 2020, 93, 1321–1332. [Google Scholar] [CrossRef]
- Crook, D.J.; Lance, D.R.; Mastro, V.C. Identification of a potential third component of the male-produced pheromone of Anoplophora glabripennis and its effect on behavior. J. Chem. Ecol. 2014, 40, 1241–1250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceja-Navarro, J.A.; Karaoz, U.; Bill, M.; Hao, Z.; White, R.A.; Arellano, A.; Ramanculova, L.; Filley, T.R.; Berry, T.D.; Conrad, M.E.; et al. Gut anatomical properties and microbial functional assembly promote lignocellulose deconstruction and colony subsistence of a wood-feeding beetle. Nat. Microbiol. 2019, 4, 864–875. [Google Scholar] [CrossRef] [Green Version]
- William, J.; Mattson, J. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol. System. 1980, 11, 119–161. [Google Scholar] [CrossRef]
- Dillon, R.J.; Dillon, V.M. The gut bacteria of insects: Nonpathogenic interactions. Annu. Rev. Entomol. 2004, 49, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Scully, E.D.; Hoover, K.; Carlson, J.E.; Tien, M.; Geib, S.M. Midgut transcriptome profiling of Anoplophora glabripennis, a lignocellulose degrading cerambycid beetle. BMC Genom. 2013, 14, 850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeling, C.I.; Bohlmann, J. Diterpene resin acids in conifers. Phytochemistry 2006, 67, 2415–2423. [Google Scholar] [CrossRef]
- Salem, H.; Kaltenpoth, M. Beetle-bacterial symbioses: Endless forms most functional. Annu. Rev. Entomol. 2022, 67, 201–219. [Google Scholar] [CrossRef]
- DiGuistini, S.; Wang, Y.; Liao, N.Y.; Taylor, G.; Tanguay, P.; Feau, N.; Henrissat, B.; Chan, S.K.; Hesse-Orce, U.; Alamouti, S.M.; et al. Genome and transcriptome analyses of the mountain pine beetle-fungal symbiont Grosmannia clavigera, a lodgepole pine pathogen. Proc. Natl. Acad. Sci. USA 2011, 108, 2504–2509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walling, L.L. Avoiding effective defenses: Strategies employed by phloem-feeding insects. Plant Physiol. 2008, 146, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Yan, Y. The study on the model of ecological control of long-horn beetle. J. Agric. Univ. Hebei 1999, 22, 83–87. [Google Scholar]
- Li, M.; Guo, X.; Zhuang, S.; Wang, D. Study on diversity of mixed forest and its pest resistance for Anoplopora glabripennis (Motschlsky). Sci. Silvae Sin. 2005, 41, 157–164. [Google Scholar] [CrossRef]
- Cao, B.; Xu, X. Effects of mixed forest of Ailanthus altissima and Populus bolleana on host choice of Anoplophora glabripennis. Sci. Silvae Sin. 2006, 42, 56–60. [Google Scholar] [CrossRef]
- Reddy, G.; Guerrero, A. Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 2004, 9, 253–261. [Google Scholar] [CrossRef]
- Xu, T.; Teale, S.A. Chemical ecology of the Asian Longhorn Beetle, Anoplophora glabripennis. J. Chem. Ecol. 2021, 47, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Oliver, J.E.; Aldrich, J.R.; Wang, B.; Mastro, V.C. Stimulatory beetle volatiles for the Asian longhorned beetle, Anoplophora glabripennis (Motschulsky). Z. Nat. C 2002, 57, 553–558. [Google Scholar] [CrossRef]
- Zhang, A.; Oliver, J.E.; Chauhan, K.; Zhao, B.; Xia, L.; Xu, Z. Evidence for contact sex recognition pheromone of the Asian longhorned beetle, Anoplophora glabripennis (Coleoptera: Cerambycidae). Naturwissenschaften 2003, 90, 410–413. [Google Scholar] [CrossRef]
- Nehme, M.E.; Keena, M.A.; Zhang, A.; Baker, T.C.; Xu, Z.; Hoover, K. Evaluating the use of male-produced pheromone components and plant volatiles in two trap designs to monitor Anoplophora glabripennis. Environ. Entomol. 2010, 39, 169–176. [Google Scholar] [CrossRef]
- Wickham, J.D.; Xu, Z.; Teale, S.A. Evidence for a female-produced, long range pheromone of Anoplophora glabripennis (Coleoptera: Cerambycidae). Insect Sci. 2012, 19, 355–371. [Google Scholar] [CrossRef]
- Meng, P.S.; Trotter, R.T.; Keena, M.A.; Baker, T.C.; Yan, S.; Schwartzberg, E.G.; Hoover, K. Effects of pheromone and plant volatile release rates and ratios on trapping Anoplophora glabripennis (Coleoptera: Cerambycidae) in China. Environ. Entomol. 2014, 43, 1379–1388. [Google Scholar] [CrossRef]
- Yu, H. Effects of Different Attractants on Trapping Anoplophora glabripennis and Anoplophora chinensis. Master’s Thesis, Beijing Forestry University, Beijing, China, 2016. [Google Scholar]
- Zhu, N.; Zhang, D.; Wu, L.; Hu, Q.; Fan, J. Attractiveness of aggregation pheromone and host plant volatiles to Anoplophora glabripennis and A. chinensis (Coleoptera: Cerambycidae). Acta Entomol. Sin. 2017, 60, 421–430. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, S.; Meng, Z.; Wang, J.; Liu, Y.; Zhang, A.; Yan, S. Improvement of trap color for Anoplophora glabripennis and determination of the optimus sustained-release amount of attractants. Sci. Silvae Sin. 2017, 53, 168–174. [Google Scholar] [CrossRef]
- Huang, J.; Qu, B.; Fang, G.; Li, X.; Zong, S. The drivers of the Asian longhorned beetle disaster show significant spatial heterogeneity. Ecol. Indic. 2020, 117, 106680. [Google Scholar] [CrossRef]
- Hoover, K.; Keena, M.; Nehme, M.; Wang, S.; Meng, P.; Zhang, A. Sex-specific trail pheromone mediates complex mate finding behavior in Anoplophora glabripennis. J. Chem. Ecol. 2014, 40, 169–180. [Google Scholar] [CrossRef]
- Hanks, L.M.; Millar, J.G. Sex and aggregation-sex pheromones of Cerambycid beetles: Basic science and practical applications. J. Chem. Ecol. 2016, 42, 631–654. [Google Scholar] [CrossRef] [PubMed]
- Segura, D.F.; Viscarret, M.M.; Ovruski, S.M.; Cladera, J.L. Response of the fruit fly parasitoid Diachasmimorpha longicaudata to host and host-habitat volatile cues. Entomol. Exp. Appl. 2012, 143, 164–176. [Google Scholar] [CrossRef]
- Li, J. Behavioral Responses and Mechanism of Anoplophora glabripennis (Motschulsky) to Host Volatile Compounds. Ph.D. Thesis, Beijing Forest University, Beijing, China, 2001. [Google Scholar]
- Song, B.; Niu, Y.; Jürg, S.; Chen, G.; Peng, D.; Gao, Y.; Sun, H. Pollinator attraction in Cornus capitata (Cornaceae): The relative role of visual and olfactory cues. J. Plant Ecol. 2015, 8, 173–181. [Google Scholar] [CrossRef] [Green Version]
- Lyu, F.; Hai, X.; Wang, Z.; Yan, A.; Liu, B. Integration of visual and olfactory cues in host plant identification by the Asian Longhorned Beetle, Anoplophora glabripennis (Motschulsky) (Coleoptera: Cerambycidae). PLoS ONE 2015, 10, e0142752. [Google Scholar] [CrossRef]
- Lyu, F. Influence of Visual and Olfactory Cues on the Behavioristics Interaction Mechanism of Host Plant-Anoplophora glabripennis-Dastarcus helophoroides. Ph.D. Thesis, Hebei Agricultural University, Baoding, China, 2016. [Google Scholar]
- Cao, B.; Li, Z.; Ji, X.; Xu, X. A Study on scattering effect of the extracts from Ailanthus altissima on Anoplophora glabripennis. J. Nanjing For. Univ. 2004, 28, 47–49. [Google Scholar] [CrossRef]
- Speed, M.P.; Fenton, A.; Jones, M.G.; Ruxton, G.D.; Brockhurst, M.A. Coevolution can explain defensive secondary metabolite diversity in plants. New Phytol. 2015, 208, 1251–1263. [Google Scholar] [CrossRef]
- Divekar, P.A.; Narayana, S.; Divekar, B.A.; Kumar, R.; Gadratagi, B.G.; Ray, A.; Singh, A.K.; Rani, V.; Singh, V.; Singh, A.K.; et al. Plant secondary metabolites as defense tools against herbivores for sustainable crop protection. Int. J. Mol. Sci. 2022, 23, 2690. [Google Scholar] [CrossRef]
- Xia, J.; Guo, Z.; Yang, Z.; Han, H.; Wang, S.; Xu, H.; Yang, X.; Yang, F.; Wu, Q.; Xie, W.; et al. Whitefly hijacks a plant detoxification gene that neutralizes plant toxins. Cell 2021, 184, 1693–1705.e1617. [Google Scholar] [CrossRef]
- Mierziak, J.; Kostyn, K.; Kulma, A. Flavonoids as important molecules of plant interactions with the environment. Molecules 2014, 19, 16240–16265. [Google Scholar] [CrossRef] [Green Version]
- Glynn, C.; Rönnberg-Wästljung, A.; Julkunen-Tiitto, R.; Weih, M. Willow genotype, but not drought treatment, affects foliar phenolic concentrations and leaf-beetle resistance. Entomol. Exp. Appl. 2004, 113, 1–14. [Google Scholar] [CrossRef]
- Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B. Phenolic glycosides of the Salicaceae and their role as anti-herbivore defenses. Phytochemistry 2011, 72, 1497–1509. [Google Scholar] [CrossRef]
- Zvereva, E.L.; Rank, N.E. Fly parasitoid Megaselia opacicornis uses defensive secretions of the leaf beetle Chrysomela lapponica to locate its host. Oecologia 2004, 140, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Kolehmainen, J.; Julkunen-Tiitto, R.; Roininen, H.; Tahvanainen, J. Phenolic glucosides as feeding cues for willow-feeding leaf beetles. Entomol. Exp. Appl. 1995, 74, 235–243. [Google Scholar] [CrossRef]
- Fernández, P.C.; Braccini, C.L.; Dávila, C.; Barrozo, R.B.; Aráoz, M.V.C.; Cerrillo, T.; Gershenzon, J.; Reichelt, M.; Zavala, J.A. The use of leaf surface contact cues during oviposition explains field preferences in the willow sawfly Nematus oligospilus. Sci. Rep. 2019, 9, 4946. [Google Scholar] [CrossRef] [Green Version]
- Sevillya, G.; Adato, O.; Snir, S. Detecting horizontal gene transfer: A probabilistic approach. BMC Genom. 2020, 21, 106. [Google Scholar] [CrossRef]
- Wybouw, N.; Pauchet, Y.; Heckel, D.G.; Van Leeuwen, T. Horizontal gene transfer contributes to the evolution of arthropod herbivory. Genome Biol. Evol. 2016, 8, 1785–1801. [Google Scholar] [CrossRef] [Green Version]
- Keeling, P.J.; Palmer, J.D. Horizontal gene transfer in eukaryotic evolution. Nat. Rev. Genet. 2008, 9, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Emamalipour, M.; Seidi, K.; Zununi Vahed, S.; Jahanban-Esfahlan, A.; Jaymand, M.; Majdi, H.; Amoozgar, Z.; Chitkushev, L.T.; Javaheri, T.; Jahanban-Esfahlan, R.; et al. Horizontal gene transfer: From evolutionary flexibility to disease progression. Front. Cell Dev. Biol. 2020, 8, 229. [Google Scholar] [CrossRef]
- Husnik, F.; McCutcheon, J.P. Functional horizontal gene transfer from bacteria to eukaryotes. Nat. Rev. Microbiol. 2018, 16, 67–79. [Google Scholar] [CrossRef]
- Wang, H.; Sun, S.; Ge, W.; Zhao, L.; Hou, B.; Wang, K.; Lyu, Z.; Chen, L.; Xu, S.; Guo, J.; et al. Horizontal gene transfer of Fhb7 from fungus underlies Fusarium head blight resistance in wheat. Science 2020, 368, eaba5435. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.R.V.; Henneberry, T.J.; Anderson, P. History, current status, and collaborative research projects for Bemisia tabaci. Crop Prot. 2001, 20, 709–723. [Google Scholar] [CrossRef] [Green Version]
- Hanks, L.M.; Millar, J.G. Field bioassays of cerambycid pheromones reveal widespread parsimony of pheromone structures, enhancement by host plant volatiles, and antagonism by components from heterospecifics. Chemoecology 2013, 23, 21–44. [Google Scholar] [CrossRef]
- Ray, A.M.; Barbour, J.D.; McElfresh, J.S.; Moreira, J.A.; Swift, I.; Wright, I.M.; Žunič, A.; Mitchell, R.F.; Graham, E.E.; Alten, R.L.; et al. 2,3-Hexanediols as sex attractants and a female-produced sex pheromone for cerambycid beetles in the prionine genus Tragosoma. J. Chem. Ecol. 2012, 38, 1151–1158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wickham, J.D.; Harrison, R.D.; Lu, W.; Guo, Z.; Millar, J.G.; Hanks, L.M.; Chen, Y. Generic lures attract cerambycid beetles in a tropical montane rain forest in southern China. J. Econ. Entomol. 2014, 107, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Wickham, J.D.; Millar, J.G.; Hanks, L.M.; Zou, Y.; Wong, J.C.; Harrison, R.D.; Chen, Y. (2R,3S)-2,3-Octanediol, a female-produced sex pheromone of Megopis costipennis (Coleoptera: Cerambycidae: Prioninae). Environ. Entomol. 2016, 45, 223–228. [Google Scholar] [CrossRef]
- Silva, W.D.; Millar, J.G.; Hanks, L.M.; Bento, J.M. (6E,8Z)-6,8-pentadecadienal, a novel attractant pheromone produced by males of the Cerambycid beetles Chlorida festiva and Chlorida costata. J. Chem. Ecol. 2016, 42, 1082–1085. [Google Scholar] [CrossRef] [PubMed]
- Engl, T.; Kaltenpoth, M. Influence of microbial symbionts on insect pheromones. Nat. Prod. Rep. 2018, 35, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Perlmutter, J.I.; Bordenstein, S.R. Microorganisms in the reproductive tissues of arthropods. Nat. Rev. Microbiol. 2020, 18, 97–111. [Google Scholar] [CrossRef]
- Engl, T.; Eberl, N.; Gorse, C.; Krüger, T.; Schmidt, T.H.P.; Plarre, R.; Adler, C.; Kaltenpoth, M. Ancient symbiosis confers desiccation resistance to stored grain pest beetles. Mol. Ecol. 2018, 27, 2095–2108. [Google Scholar] [CrossRef]
- Wang, L.; Li, C.; Wang, X.; Wang, G.; Shang, S.; Dou, Z.; Luo, Y. Gut lignocellulose activity and microbiota in Asian longhorned beetle and their predicted contribution to larval nutrition. Front. Microbiol. 2022, 13, 899865. [Google Scholar] [CrossRef]
- Geib, S.M.; Jimenez-Gasco Mdel, M.; Carlson, J.E.; Tien, M.; Hoover, K. Effect of host tree species on cellulase activity and bacterial community composition in the gut of larval Asian longhorned beetle. Environ. Entomol. 2009, 38, 686–699. [Google Scholar] [CrossRef]
- Hu, P.; Wang, J.; Cui, M.; Tao, J.; Luo, Y. Antennal transcriptome analysis of the Asian longhorned beetle Anoplophora glabripennis. Sci. Rep. 2016, 6, 26652. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Hao, E.; Li, Y.; Yang, H.; Sun, P.; Lu, P.; Qiao, H. Antennal transcriptome analysis of olfactory genes and tissue expression profiling of odorant binding proteins in Semanotus bifasciatus (cerambycidae: Coleoptera). BMC Genom. 2022, 23, 461. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hu, P.; Gao, P.; Tao, J.; Luo, Y. Antennal transcriptome analysis and expression profiles of olfactory genes in Anoplophora chinensis. Sci. Rep. 2017, 7, 15470. [Google Scholar] [CrossRef] [Green Version]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Giannone, F.; Moino, S.; Rassati, D. Exploiting trap color to improve surveys of longhorn beetles. J. Pest Sci. 2021, 94, 871–883. [Google Scholar] [CrossRef]
- Cavaletto, G.; Faccoli, M.; Marini, L.; Spaethe, J.; Magnani, G.; Rassati, D. Effect of trap color on captures of bark-and wood-boring beetles (Coleoptera; Buprestidae and Scolytinae) and associated predators. Insects 2020, 11, 749. [Google Scholar] [CrossRef]
- Kim, K.N.; Huang, Q.Y.; Lei, C.L. Advances in insect phototaxis and application to pest management: A review. Pest Manag. Sci. 2019, 75, 3135–3143. [Google Scholar] [CrossRef]
- Lei, B.; Cai, X.M.; Luo, Z.X.; Li, Z.Q.; Chen, Z.M. Decreased capture of natural enemies of pests in light traps with light-emitting diode technology. Ann. Appl. Biol. 2018, 173, 251–260. [Google Scholar]
- McQuate, G.T. Green light synergistally enhances male sweetpotato weevil response to sex pheromone. Sci. Rep. 2014, 4, 4499. [Google Scholar] [CrossRef] [Green Version]
- Nieri, R.; Anfora, G.; Mazzoni, V.; Stacconi, M. Semiochemicals, semiophysicals and their integration for the development of innovative multi-modal systems for agricultural pests’ monitoring and control. Entomol. Gen. 2022, 42, 167–183. [Google Scholar] [CrossRef]
- Lyu, F.; Hai, X.; Wang, Z.; Yan, A.; Bi, Y.; Liu, B. Diurnal rhythm of four types of adult behaviors of Anoplophora glabripennis. J. Northeast For. Univ. 2015, 43, 90–95. [Google Scholar] [CrossRef]
- Briscoe, A.D.; Chittka, L. The evolution of color vision in insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futahashi, R.; Kawahara-Miki, R.; Kinoshita, M.; Yoshitake, K.; Yajima, S.; Arikawa, K.; Fukatsu, T. Extraordinary diversity of visual opsin genes in dragonflies. Proc. Natl. Acad. Sci. USA 2015, 112, E1247–E1256. [Google Scholar] [CrossRef] [Green Version]
- Meece, M.; Rathore, S.; Buschbeck, E.K. Stark trade-offs and elegant solutions in arthropod visual systems. J. Exp. Biol. 2021, 224, jeb215541. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, C.R.; Fujimoto, M.S.; Lord, N.P.; Shin, S.; Mckenna, D.D.; Suvorov, A.; Martin, G.J.; Bybee, S.M. Overcoming the loss of blue sensitivity through opsin duplication in the largest animal group, beetles. Sci. Rep. 2017, 7, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terakita, A. The opsins. Genome Biol. 2005, 6, 213. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Attractive Effect | Type | Semichemical Compound | Number | Reference(s) |
|---|---|---|---|---|
| Attraction | Alcohols | Butanol, pentanol, linalool, Cis-3-hexen-1-ol | 4 | [64,65,66,67] |
| Aldehyde | Nonanal | 1 | [67] | |
| Olefins | β-caryophyllene, δ-3-carene, S-β-pinene, R-α-pinene, camphene, D-limonene (attracts only females), phellandrene | 7 | [44,67,68,69] | |
| Ester | Ethyl acetate | 1 | [44] | |
| Repellent | Alcohols | trans-2-hexenol, (E)-1-pentylene-3-ol | 2 | [25] |
| Olefins | Heptylene, ocimene, myrcene, β-caryophyllene, β-pinene | 5 | [25,44,67,69,70] | |
| Ketones | Geranyl acetone | 1 | [25] | |
| Ester | 3-Hexenyl acetate | 1 | [68,69] |
| OBP | High Expression Tissue | Chemical Type(s) | Optimal Ligand(s) | Ki (Μm/L) | References |
|---|---|---|---|---|---|
| AglaOBP1 | ML, FA, MA, FL & | Host kairomone | Cis-3-Hexen-1-ol | 5.88 | [78,79,80] |
| Cis-2-Hexen-1-ol | 8.33 | ||||
| α-Pinene | 9.07 | ||||
| Host kairomone and volatile pheromone | β-caryophyllene | 7.47 | |||
| AglaOBP12 | FA, MA, ML, FL | Host kairomone and volatile pheromone | β-caryophyllene | 0.74 | [77,80,81] |
| Host kairomone | Cis-3-Hexenyl acetate | 0.77 | |||
| Dodecanal | 0.82 | ||||
| 1-tetradecanol | 0.96 | ||||
| Volatile pheromone | α-Farnesol | 1.03 | |||
| AglaOBP45 | MA | Host kairomone | Benzyl alcohol | 0.92 | [79] |
| 2-Pentanol | 0.96 | ||||
| α-Ocimene | 1.02 | ||||
| α-Pinene | 1.92 | ||||
| Long-range sex pheromone | Hexadecanal | 0.87 | |||
| Female contact and trail sex pheromone | (Z)-9-tricosene | 2.02 | |||
| (Z)-9-pentacosene | 6.81 | ||||
| AglaOBP46 | MA, FA, ML, FL | Host kairomone | 1-Dodecanol | 0.74 | [79] |
| Benzyl alcohol | 0.82 | ||||
| 2-Pentanol | 2.12 | ||||
| (+)-longifolene | 1.78 | ||||
| D-limonene | 2.01 |
| Study | Chemical Matter | Number Captured | Interval Time | Site | Reference |
|---|---|---|---|---|---|
| 1 | (-)-linalool | 3.00 ± 1.00 | 7 days | Green house | [68] |
| 2 | (-)-linalool + MP | F: 3.70 ± 3.20 | 7 days | Ningxia, China | [97] |
| M: 1.70 ± 1.20 | 7 days | ||||
| 3 | Heptanal, nonanal, hexadecanal (1:7:1) | 4.70 ± 2.20 | 7 days | Ningxia, China | [98] |
| 4 | Heptanal, nonanal, hexadecanal + HK | 3.20 ± 1.20 | 7 days | ||
| 5 | MP + PV (1:1) | 2.25 | 7 days | Syracuse, NY, USA | [99] |
| 6 | PV (8:9:1) | 1.10 ± 0.89 | 7 days | Cixi, China | [100] |
| 7 | MK | 3.40 ± 1.10 | 7 days | Cixi, China | [101] |
| 8 | Brown trap + 2-pentanol | F: 1.52 ± 0.12 | 7 days | Hengshui, China | [102] |
| M: 1.00 ± 0.09 | |||||
| 9 | 25 mg α-longipinene | 0.90 ± 0.50 | 7 days | Huanyuan, China | [80] |
| 5 mg α-longipinene + pv | 1.10 ± 0.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lyu, F.; Hai, X.; Wang, Z. A Review of the Host Plant Location and Recognition Mechanisms of Asian Longhorn Beetle. Insects 2023, 14, 292. https://doi.org/10.3390/insects14030292
Lyu F, Hai X, Wang Z. A Review of the Host Plant Location and Recognition Mechanisms of Asian Longhorn Beetle. Insects. 2023; 14(3):292. https://doi.org/10.3390/insects14030292
Chicago/Turabian StyleLyu, Fei, Xiaoxia Hai, and Zhigang Wang. 2023. "A Review of the Host Plant Location and Recognition Mechanisms of Asian Longhorn Beetle" Insects 14, no. 3: 292. https://doi.org/10.3390/insects14030292