Sericin Ser3 Ectopic Expressed in Posterior Silk Gland Affects Hemolymph Immune Melanization Response via Reducing Melanin Synthesis in Silkworm

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animal Preparation
2.2. Hemolymph Melanization Speed
2.3. Plasma Melanin Content
2.4. Bactericidal Capacity of Plasma Melanism
2.5. Real-Time Fluorescence Quantitative Polymerase Chain Reaction (PCR)
2.6. Enzyme Activity
2.7. Hemolymph Protease Inhibitor Activity
2.8. Plasma Redox Substrate Content
2.9. High Temperature Resistance
2.10. Data Analysis
3. Results
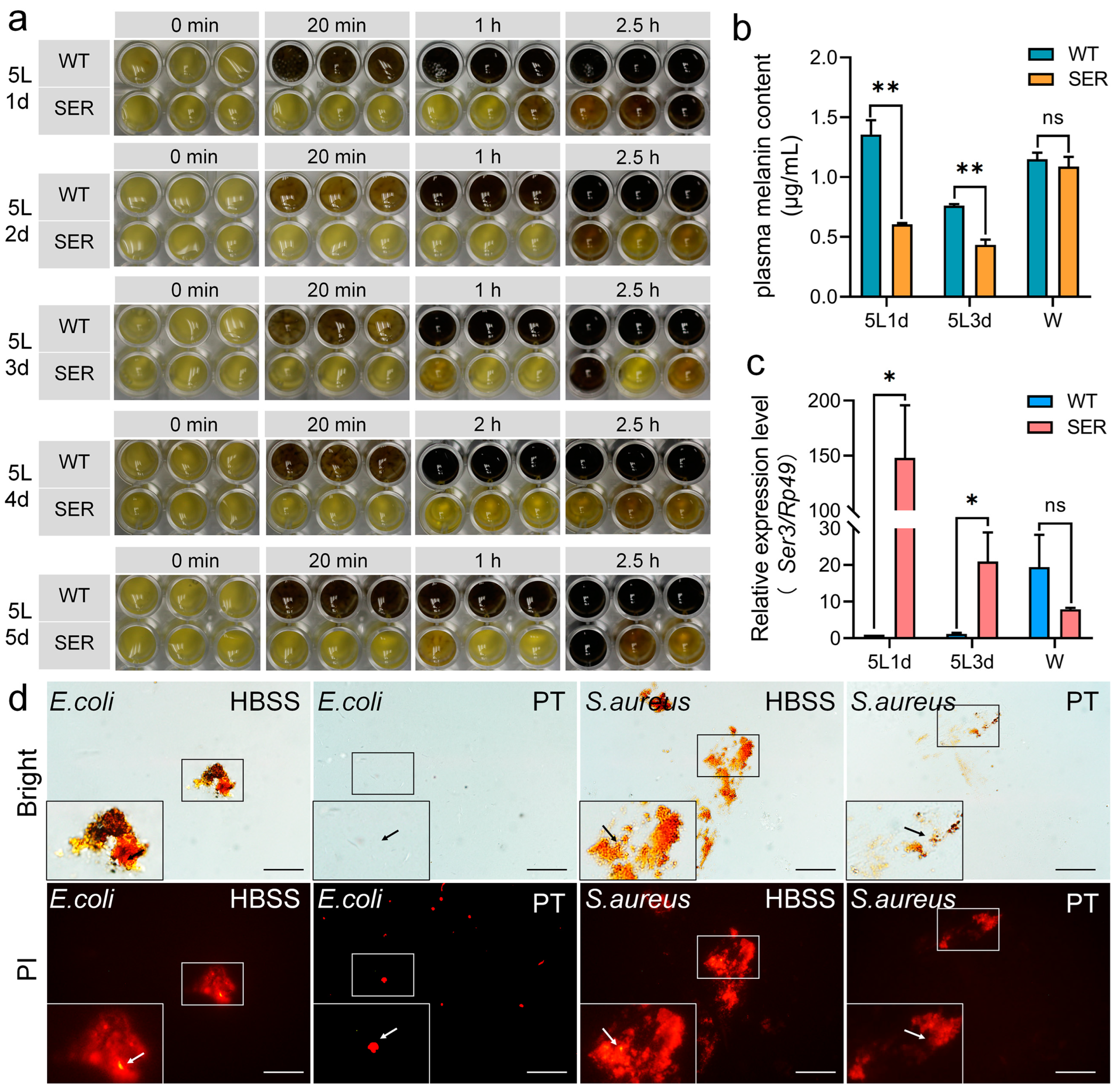
3.1. The Recombinant Ser3 Gene Expressed in the Posterior Silk Gland Affects the Hemolymph Immune Melanization Response of Silkworms
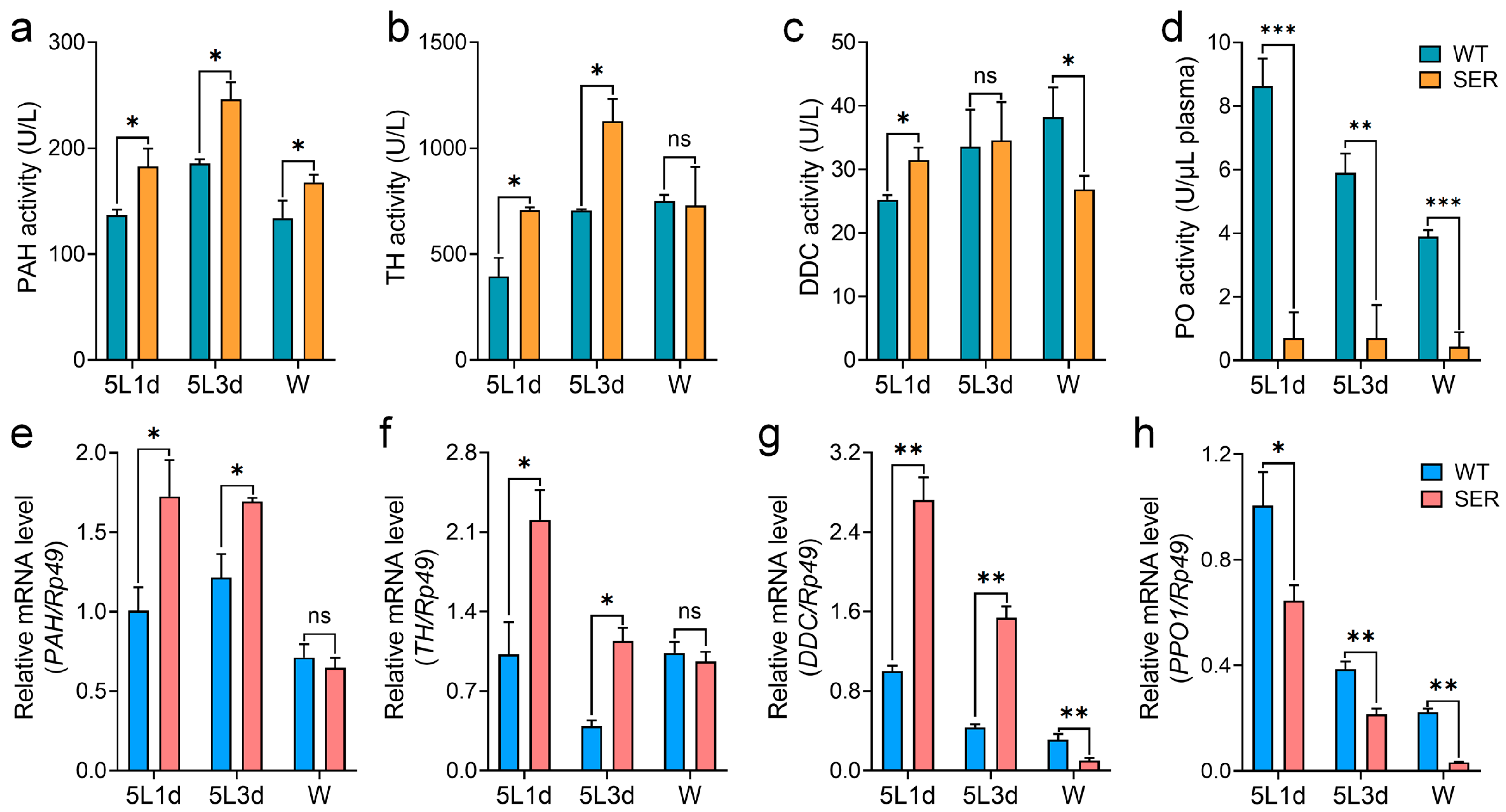
3.2. Mutant Silkworm Affects Hemolymph Melanization by Changing the Serine Protease Cascade
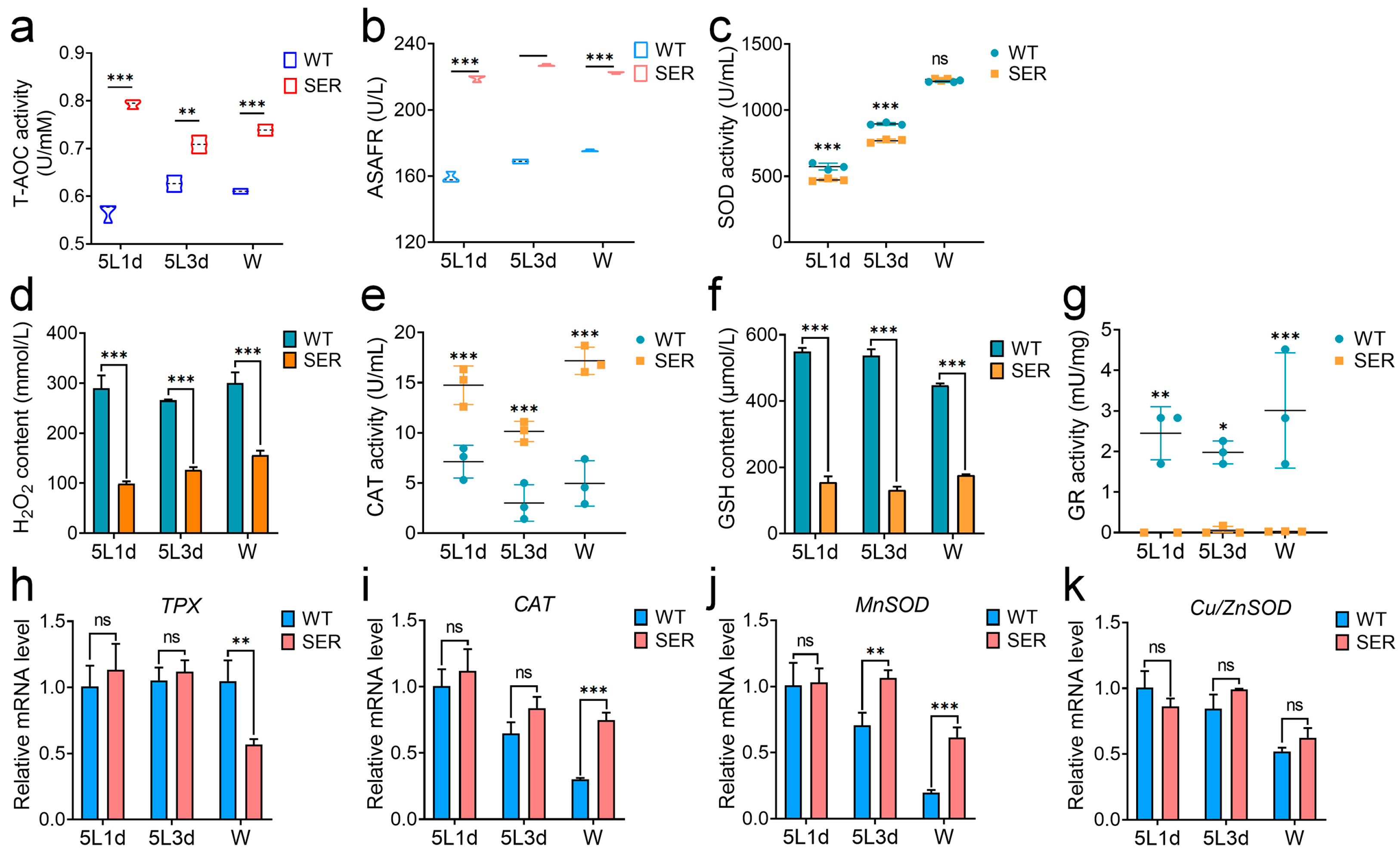
3.3. Mutant Silkworm Affects Hemolymph Melanization by Changing the Level of Hemolymph Redox Metabolism
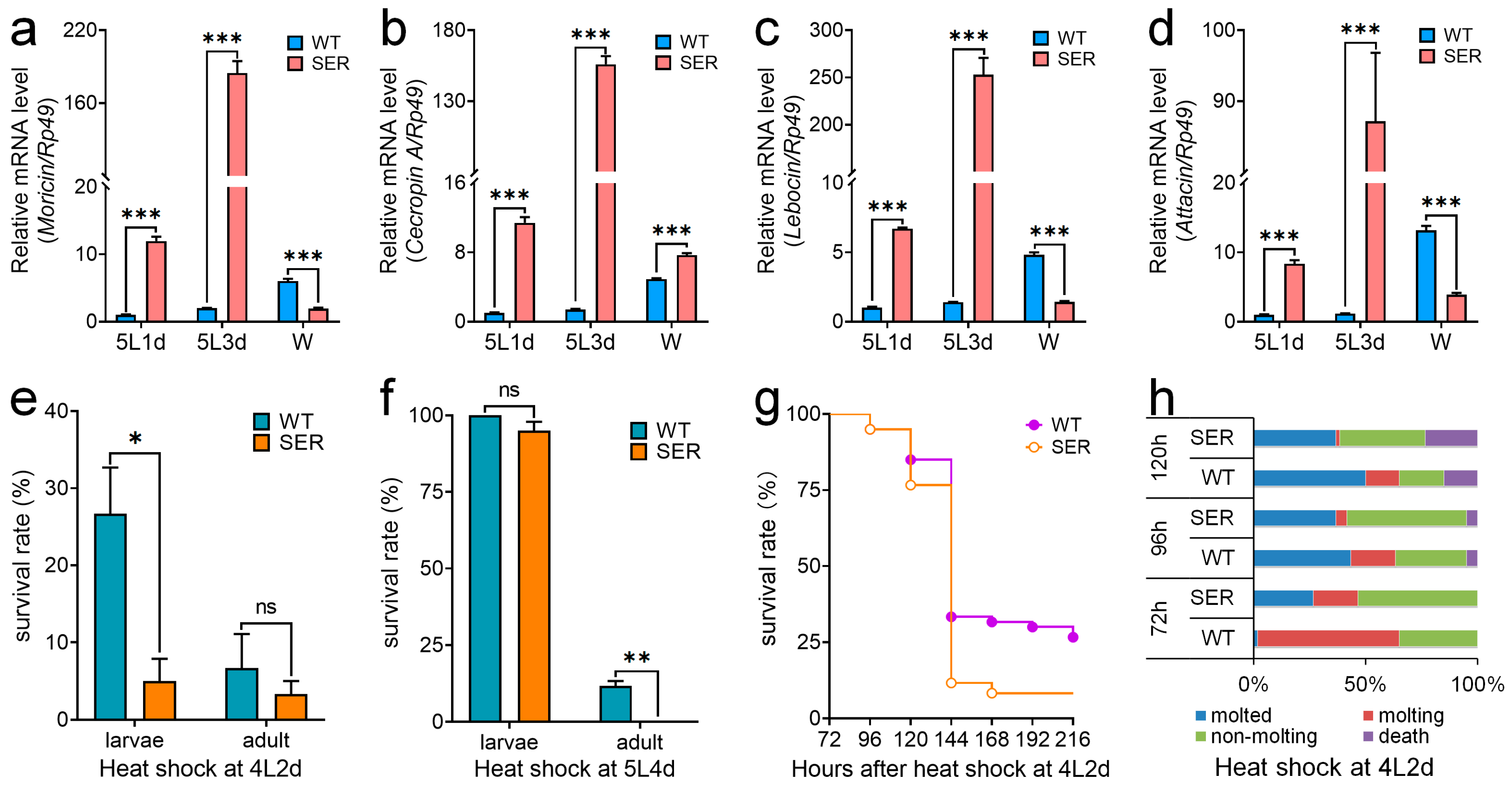
3.4. Mutant Silkworm Affects the Immune Resistance of Silkworms
4. Discussion
4.1. The Recombinant Ser3 Expression in Posterior Silk Glands Affects Hemolymph Metabolism
4.2. The Recombinant Ser3 Expression in the Posterior Silk Gland Affects the Immune Ability of Silkworms
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Chen, X.; Wang, Y.; Wang, Y.; Li, Q.; Liang, X.; Wang, G.; Li, J.; Peng, R.; Sima, Y.; Xu, S. Ectopic expression of sericin enables efficient production of ancient silk with structural changes in silkworm. Nat. Commun. 2022, 13, 6295. [Google Scholar] [CrossRef]
- Omenetto, F.G.; Kaplan, D.L. New opportunities for an ancient material. Science 2010, 329, 528–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, A.A.; Yamaji, Y.; Kajiwara, K.; Takaki, K.; Mori, H.; Liew, M.W.O.; Kotani, E.; Maruta, R. A Bioengineering Approach for the Development of Fibroblast Growth Factor-7-Functionalized Sericin Biomaterial Applicable for the Cultivation of Keratinocytes. Int. J. Mol. Sci. 2022, 23, 9953. [Google Scholar] [CrossRef] [PubMed]
- Yamada, K.; Masuda, K.; Ida, S.; Tada, H.; Bando, M.; Abe, K.; Tatematsu, K.I.; Sezutsu, H.; Oyama, T.; Chikamatsu, K.; et al. In vitro assessment of antitumor immune responses using tumor antigen proteins produced by transgenic silkworms. J. Mater. Sci. Mater. Med. 2021, 32, 58. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, H.; Sezutsu, H.; Tamura, T.; Yamada, K. Production of an active feline interferon in the cocoon of transgenic silkworms using the fibroin H-chain expression system. Biochem. Biophys. Res. Commun. 2007, 355, 976–980. [Google Scholar] [CrossRef]
- Hino, R.; Tomita, M.; Yoshizato, K. The generation of germline transgenic silkworms for the production of biologically active recombinant fusion proteins of fibroin and human basic fibroblast growth factor. Biomaterials 2006, 27, 5715–5724. [Google Scholar] [CrossRef]
- Teulé, F.; Miao, Y.G.; Sohn, B.H.; Kim, Y.S.; Hull, J.J.; Fraser, M.J., Jr.; Lewis, R.V.; Jarvis, D.L. Silkworms transformed with chimeric silkworm/spider silk genes spin composite silk fibers with improved mechanical properties. Proc. Natl. Acad. Sci. USA 2012, 109, 923–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Dong, Q.; Yu, Y.; Niu, B.; Ji, D.; Li, M.; Huang, Y.; Chen, X.; Tan, A. Mass spider silk production through targeted gene replacement in Bombyx mori. Proc. Natl. Acad. Sci. USA 2018, 115, 8757–8762. [Google Scholar] [CrossRef] [Green Version]
- Otsuki, R.; Yamamoto, M.; Matsumoto, E.; Iwamoto, S.I.; Sezutsu, H.; Suzui, M.; Takaki, K.; Wakabayashi, K.; Mori, H.; Kotani, E. Bioengineered silkworms with butterfly cytotoxin-modified silk glands produce sericin cocoons with a utility for a new biomaterial. Proc. Natl. Acad. Sci. USA 2017, 114, 6740–6745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Wang, L.; Wang, Y.; Tao, H.; Yin, W.; Sima, Y.; Wang, Y.; Xu, S.Q. High yield exogenous HPL protein production in the Bombyx mori silk gland provides novel insight into recombinant expression systems. Sci. Rep. 2015, 5, 13839. [Google Scholar] [CrossRef] [Green Version]
- Tomita, M.; Munetsuna, H.; Sato, T.; Adachi, T.; Hino, R.; Hayashi, M.; Shimizu, K.; Nakamura, N.; Tamura, T.; Yoshizato, K. Transgenic silkworms produce recombinant human type III procollagen in cocoons. Nat. Biotechnol. 2003, 21, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Tamura, T.; Thibert, C.; Royer, C.; Kanda, T.; Abraham, E.; Kamba, M.; Komoto, N.; Thomas, J.L.; Mauchamp, B.; Chavancy, G.; et al. Germline transformation of the silkworm Bombyx mori L. using a piggyBac transposon-derived vector. Nat. Biotechnol. 2000, 18, 81–84. [Google Scholar] [CrossRef]
- Uchino, K.; Sezutsu, H.; Imamura, M.; Kobayashi, I.; Tatematsu, K.; Iizuka, T.; Yonemura, N.; Mita, K.; Tamura, T. Construction of a piggyBac-based enhancer trap system for the analysis of gene function in silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2008, 38, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Horn, C.; Offen, N.; Nystedt, S.; Häcker, U.; Wimmer, E.A. piggyBac-based insertional mutagenesis and enhancer detection as a tool for functional insect genomics. Genetics 2003, 163, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Nolan, T.; Bower, T.M.; Brown, A.E.; Crisanti, A.; Catteruccia, F. piggyBac-mediated germline transformation of the malaria mosquito Anopheles stephensi using the red fluorescent protein dsRED as a selectable marker. J. Biol. Chem. 2002, 277, 8759–8762. [Google Scholar] [CrossRef] [Green Version]
- Yonemura, N.; Tamura, T.; Uchino, K.; Kobayashi, I.; Tatematsu, K.; Iizuka, T.; Sezutsu, H.; Muthulakshmi, M.; Nagaraju, J.; Kusakabe, T. PhiC31 integrase-mediated cassette exchange in silkworm embryos. Mol. Genet. Genom. 2012, 287, 731–739. [Google Scholar] [CrossRef]
- Nakayama, G.; Kawaguchi, Y.; Koga, K.; Kusakabe, T. Site-specific gene integration in cultured silkworm cells mediated by phiC31 integrase. Mol. Genet. Genom. 2006, 275, 1–8. [Google Scholar] [CrossRef]
- Imamura, M.; Nakai, J.; Inoue, S.; Quan, G.X.; Kanda, T.; Tamura, T. Targeted gene expression using the GAL4/UAS system in the silkworm Bombyx mori. Genetics 2003, 165, 1329–1340. [Google Scholar] [CrossRef]
- Tatematsu, K.; Kobayashi, I.; Uchino, K.; Sezutsu, H.; Iizuka, T.; Yonemura, N.; Tamura, T. Construction of a binary transgenic gene expression system for recombinant protein production in the middle silk gland of the silkworm Bombyx mori. Transgenic Res. 2010, 19, 473–487. [Google Scholar] [CrossRef]
- Baci, G.M.; Cucu, A.A.; Giurgiu, A.I.; Muscă, A.S.; Bagameri, L.; Moise, A.R.; Bobiș, O.; Rațiu, A.C.; Dezmirean, D.S. Advances in Editing Silkworms (Bombyx mori) Genome by Using the CRISPR-Cas System. Insects 2021, 13, 28. [Google Scholar] [CrossRef]
- Brady, D.; Saviane, A.; Cappellozza, S.; Sandrelli, F. An Efficient Workflow for Screening and Stabilizing CRISPR/Cas9-Mediated Mutant Lines in Bombyx mori. Methods Protoc. 2020, 4, 4. [Google Scholar] [CrossRef] [PubMed]
- Tomihara, K.; Andolfatto, P.; Kiuchi, T. Allele-specific knockouts reveal a role for apontic-like in the evolutionary loss of larval melanin pigmentation in the domesticated silkworm, Bombyx mori. Insect Mol. Biol. 2022, 31, 701–710. [Google Scholar] [CrossRef]
- Jiang, L.; Xia, Q. The progress and future of enhancing antiviral capacity by transgenic technology in the silkworm Bombyx mori. Insect Biochem. Mol. Biol. 2014, 48, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yashwant, R.S.; Thomas, D.S.; Manoharan, C.; Roy, G.; Kunjupillai, V.; Mishra, R.K.; Nongthomba, U.; Gopalapillai, R. Transgenic Silkworms Overexpressing Relish and Expressing Drosomycin Confer Enhanced Immunity to Multiple Pathogens. Mol. Biotechnol. 2022, 64, 711–724. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Xia, F.; Jiang, L.; Guo, H.; Xu, G.; Sun, Q.; Wang, B.; Wang, Y.; Lu, Z.; Xia, Q. Enhanced antiviral immunity against Bombyx mori cytoplasmic polyhedrosis virus via overexpression of peptidoglycan recognition protein S2 in transgenic silkworms. Dev. Comp. Immunol. 2018, 87, 84–89. [Google Scholar] [CrossRef]
- Jiang, L.; Cheng, T.; Zhao, P.; Yang, Q.; Wang, G.; Jin, S.; Lin, P.; Xiao, Y.; Xia, Q. Resistance to BmNPV via overexpression of an exogenous gene controlled by an inducible promoter and enhancer in transgenic silkworm, Bombyx mori. PLoS ONE 2012, 7, e41838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.F.; Wang, G.; Li, J.L.; Qu, Y.X.; Liang, X.Y.; Chen, X.D.; Sima, Y.H.; Xu, S.Q. Influence of Hyperproteinemia on Insect Innate Immune Function of the Circulatory System in Bombyx mori. Biology 2021, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Cooper, D.; Wuebbolt, C.; Heryanto, C.; Eleftherianos, I. The prophenoloxidase system in Drosophila participates in the anti-nematode immune response. Mol. Immunol. 2019, 109, 88–98. [Google Scholar] [CrossRef]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Li, H.; Hao, F.; Li, N.; Liu, X.; Wang, G.; Wang, Y.; Tang, H. Developmental Changes for the Hemolymph Metabolome of Silkworm (Bombyx mori L.). J. Proteome Res. 2015, 14, 2331–2347. [Google Scholar] [CrossRef] [Green Version]
- Peng, L.H.; Liu, S.; Xu, S.Y.; Chen, L.; Shan, Y.H.; Wei, W.; Liang, W.Q.; Gao, J.Q. Inhibitory effects of salidroside and paeonol on tyrosinase activity and melanin synthesis in mouse B16F10 melanoma cells and ultraviolet B-induced pigmentation in guinea pig skin. Phytomedicine 2013, 20, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, L.; Lin, Z.; Zou, Z.; Lu, Z. Serpin-5 regulates prophenoloxidase activation and antimicrobial peptide pathways in the silkworm, Bombyx mori. Insect Biochem. Mol. Biol. 2016, 73, 27–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suwanchaichinda, C.; Ochieng, R.; Zhuang, S.; Kanost, M.R. Manduca sexta serpin-7, a putative regulator of hemolymph prophenoloxidase activation. Insect Biochem. Mol. Biol. 2013, 43, 555–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nappi, A.; Poirié, M.; Carton, Y. The role of melanization and cytotoxic by-products in the cellular immune responses of Drosophila against parasitic wasps. Adv. Parasitol. 2009, 70, 99–121. [Google Scholar] [CrossRef] [PubMed]
- Sugumaran, M.; Barek, H. Critical Analysis of the Melanogenic Pathway in Insects and Higher Animals. Int. J. Mol. Sci. 2016, 17, 1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwama, R.; Ashida, M. Biosynthesis of prophenoloxidase in hemocytes of larval hemolymph of the silkworm, Bombyx mori. Insect Biochem. 1986, 16, 547–555. [Google Scholar] [CrossRef]
- Tokura, A.; Fu, G.S.; Sakamoto, M.; Endo, H.; Tanaka, S.; Kikuta, S.; Tabunoki, H.; Sato, R. Factors functioning in nodule melanization of insects and their mechanisms of accumulation in nodules. J. Insect Physiol. 2014, 60, 40–49. [Google Scholar] [CrossRef]
- Satoh, D.; Horii, A.; Ochiai, M.; Ashida, M. Prophenoloxidase-activating enzyme of the silkworm, Bombyx mori. Purification, characterization, and cDNA cloning. J. Biol. Chem. 1999, 274, 7441–7453. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Liu, H.; Fu, H.; Zhang, L.; Guo, P.; Xia, Q.; Zhao, P. Silkworm serpin32 functions as a negative-regulator in prophenoloxidase activation. Dev. Comp. Immunol. 2019, 91, 123–131. [Google Scholar] [CrossRef]
- Li, B.; Yu, H.Z.; Ye, C.J.; Ma, Y.; Li, X.; Fan, T.; Chen, F.S.; Xu, J.P. Bombyx mori Serpin6 regulates prophenoloxidase activity and the expression of antimicrobial proteins. Gene 2017, 610, 64–70. [Google Scholar] [CrossRef]
- Zhou, X.S.; Chen, C.; Li, T.H.; Tang, J.J.; Zhu, B.J.; Wei, G.Q.; Qian, C.; Liu, C.L.; Wang, L. A QM protein from Bombyx mori negatively regulates prophenoloxidase activation and melanization by interacting with Jun protein. Insect Mol. Biol. 2019, 28, 578–590. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, X.; Tan, J.; Zhang, K.; Guan, X.; Patterson, L.H.; Ding, H.; Cui, H. A novel Lozenge gene in silkworm, Bombyx mori regulates the melanization response of hemolymph. Dev. Comp. Immunol. 2015, 53, 191–198. [Google Scholar] [CrossRef]
- Bao, Y.Y.; Xue, J.; Wu, W.J.; Wang, Y.; Lv, Z.Y.; Zhang, C.X. An immune-induced reeler protein is involved in the Bombyx mori melanization cascade. Insect Biochem. Mol. Biol. 2011, 41, 696–706. [Google Scholar] [CrossRef]
- Terenius, O.; Bettencourt, R.; Lee, S.Y.; Li, W.; Söderhäll, K.; Faye, I. RNA interference of Hemolin causes depletion of phenoloxidase activity in Hyalophora cecropia. Dev. Comp. Immunol. 2007, 31, 571–575. [Google Scholar] [CrossRef]
- Bae, J.S.; Han, M.; Yao, C.; Chung, J.H. Chaetocin inhibits IBMX-induced melanogenesis in B16F10 mouse melanoma cells through activation of ERK. Chem. Biol. Interact. 2016, 245, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.J.; Lin, C.C.; Lu, T.M.; Li, J.H.; Chen, I.S.; Kuo, Y.H.; Ko, H.H. Chemical constituents derived from Artocarpus xanthocarpus as inhibitors of melanin biosynthesis. Phytochemistry 2015, 117, 424–435. [Google Scholar] [CrossRef]
- Funasaka, Y.; Komoto, M.; Ichihashi, M. Depigmenting effect of alpha-tocopheryl ferulate on normal human melanocytes. Pigment Cell Res. 2000, 13, 170–174. [Google Scholar] [CrossRef]
- Fu, W.; Wu, Z.; Zheng, R.; Yin, N.; Han, F.; Zhao, Z.; Dai, M.; Han, D.; Wang, W.; Niu, L. Inhibition mechanism of melanin formation based on antioxidant scavenging of reactive oxygen species. Analyst 2022, 147, 2703–2711. [Google Scholar] [CrossRef] [PubMed]
- Gebicki, J.M. Oxidative stress, free radicals and protein peroxides. Arch. Biochem. Biophys. 2016, 595, 33–39. [Google Scholar] [CrossRef]
- Cerenius, L.; Lee, B.L.; Söderhäll, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends. Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef]
- Kan, H.; Kim, C.H.; Kwon, H.M.; Park, J.W.; Roh, K.B.; Lee, H.; Park, B.J.; Zhang, R.; Zhang, J.; Söderhäll, K.; et al. Molecular control of phenoloxidase-induced melanin synthesis in an insect. J. Biol. Chem. 2008, 283, 25316–25323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marieshwari, B.N.; Bhuvaragavan, S.; Sruthi, K.; Mullainadhan, P.; Janarthanan, S. Insect phenoloxidase and its diverse roles: Melanogenesis and beyond. J. Comp. Physiol. B 2023, 193, 1–23. [Google Scholar] [CrossRef]
- Lin, Y.R.; Parikh, H.; Park, Y. Stress resistance and lifespan enhanced by downregulation of antimicrobial peptide genes in the Imd pathway. Aging 2018, 10, 622–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Huang, C.; Wang, B.; Guo, H.; Sun, Q.; Xia, F.; Xu, G.; Xia, Q. Enhanced heat tolerance in transgenic silkworm via overexpression of Pyrococcus furiosus superoxide reductase. Insect Biochem. Mol. Biol. 2018, 92, 40–44. [Google Scholar] [CrossRef] [PubMed]
- Kültz, D. DNA damage signals facilitate osmotic stress adaptation. Am. J. Physiol. Ren. Physiol. 2005, 289, F504–F505. [Google Scholar] [CrossRef] [Green Version]
- Tower, J. Heat shock proteins and Drosophila aging. Exp. Gerontol. 2011, 46, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Hayakawa, Y. N-acetyltyrosine-induced redox signaling in hormesis. Biochim. Biophys. Acta Mol. Cell. Res. 2021, 1868, 118990. [Google Scholar] [CrossRef]
- Matsumura, T.; Matsumoto, H.; Hayakawa, Y. Heat stress hardening of oriental armyworms is induced by a transient elevation of reactive oxygen species during sublethal stress. Arch. Insect Biochem. Physiol. 2017, 96, e21421. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Shi, M.; Yang, J.; Ma, L.; Chen, X.; Xu, M.; Peng, R.; Wang, G.; Pan, Z.; Sima, Y.; et al. Sericin Ser3 Ectopic Expressed in Posterior Silk Gland Affects Hemolymph Immune Melanization Response via Reducing Melanin Synthesis in Silkworm. Insects 2023, 14, 245. https://doi.org/10.3390/insects14030245
Wang Y, Shi M, Yang J, Ma L, Chen X, Xu M, Peng R, Wang G, Pan Z, Sima Y, et al. Sericin Ser3 Ectopic Expressed in Posterior Silk Gland Affects Hemolymph Immune Melanization Response via Reducing Melanin Synthesis in Silkworm. Insects. 2023; 14(3):245. https://doi.org/10.3390/insects14030245
Chicago/Turabian StyleWang, Yongfeng, Meijuan Shi, Jiameng Yang, Lu Ma, Xuedong Chen, Meng Xu, Ruji Peng, Guang Wang, Zhonghua Pan, Yanghu Sima, and et al. 2023. "Sericin Ser3 Ectopic Expressed in Posterior Silk Gland Affects Hemolymph Immune Melanization Response via Reducing Melanin Synthesis in Silkworm" Insects 14, no. 3: 245. https://doi.org/10.3390/insects14030245