
An Expanded View on the Morphological Diversity of Long-Nosed Antlion Larvae Further Supports a Decline of Silky Lacewings in the Past 100 Million Years
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Material
2.2. Imaging and Documentation
2.3. Image Processing and Presentation
2.4. Measurements
2.5. Shape Analysis
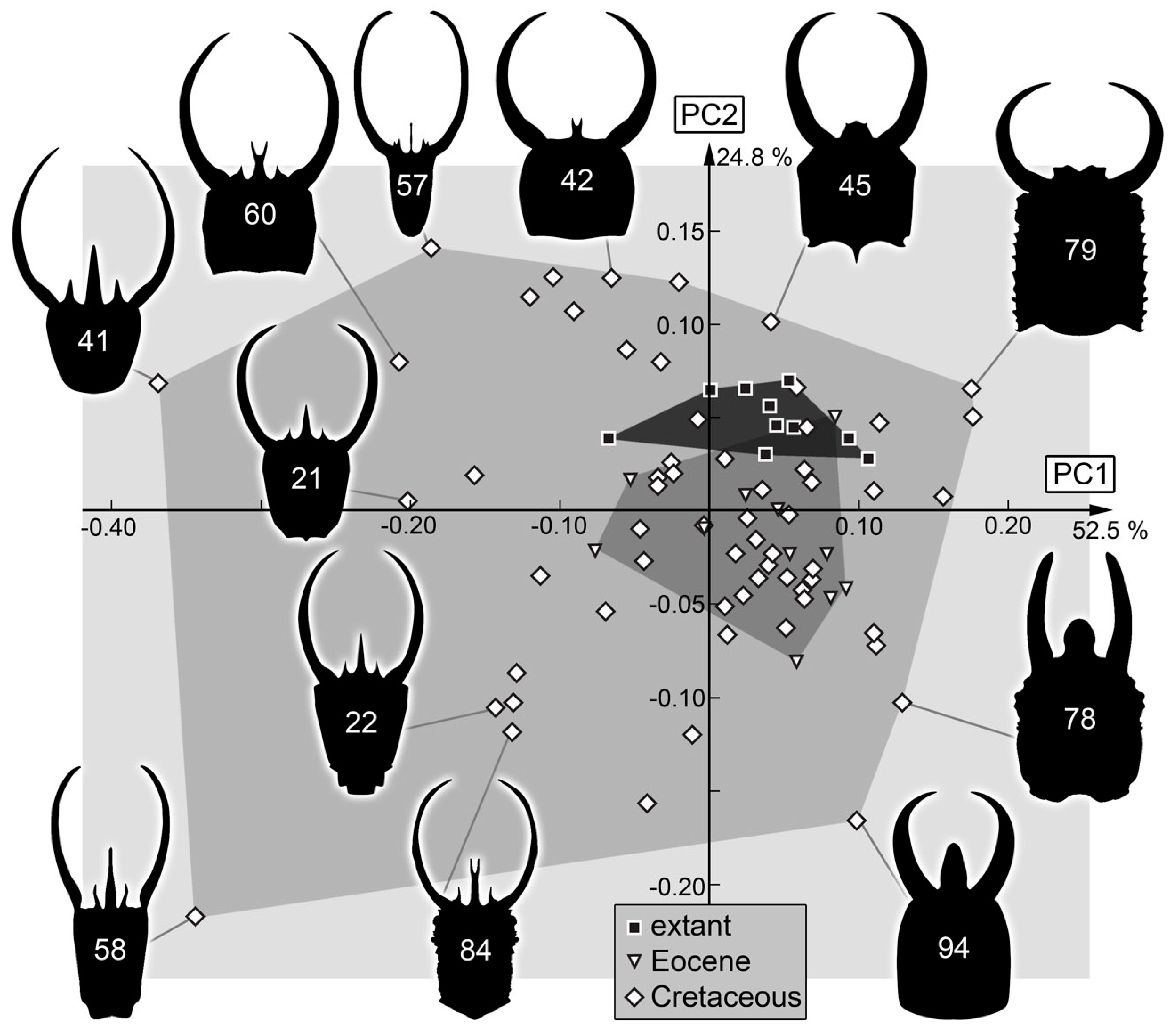
3. Results
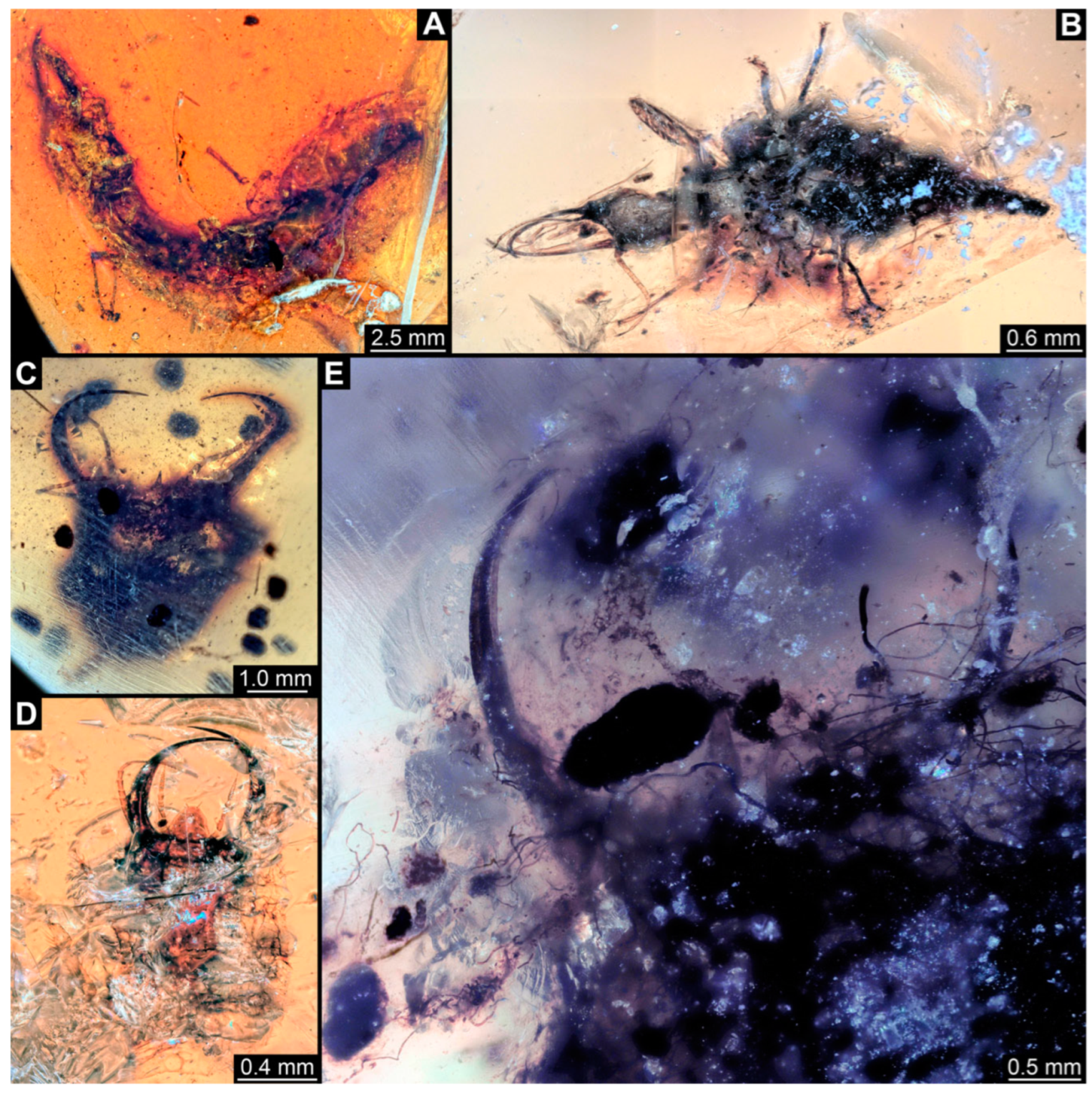
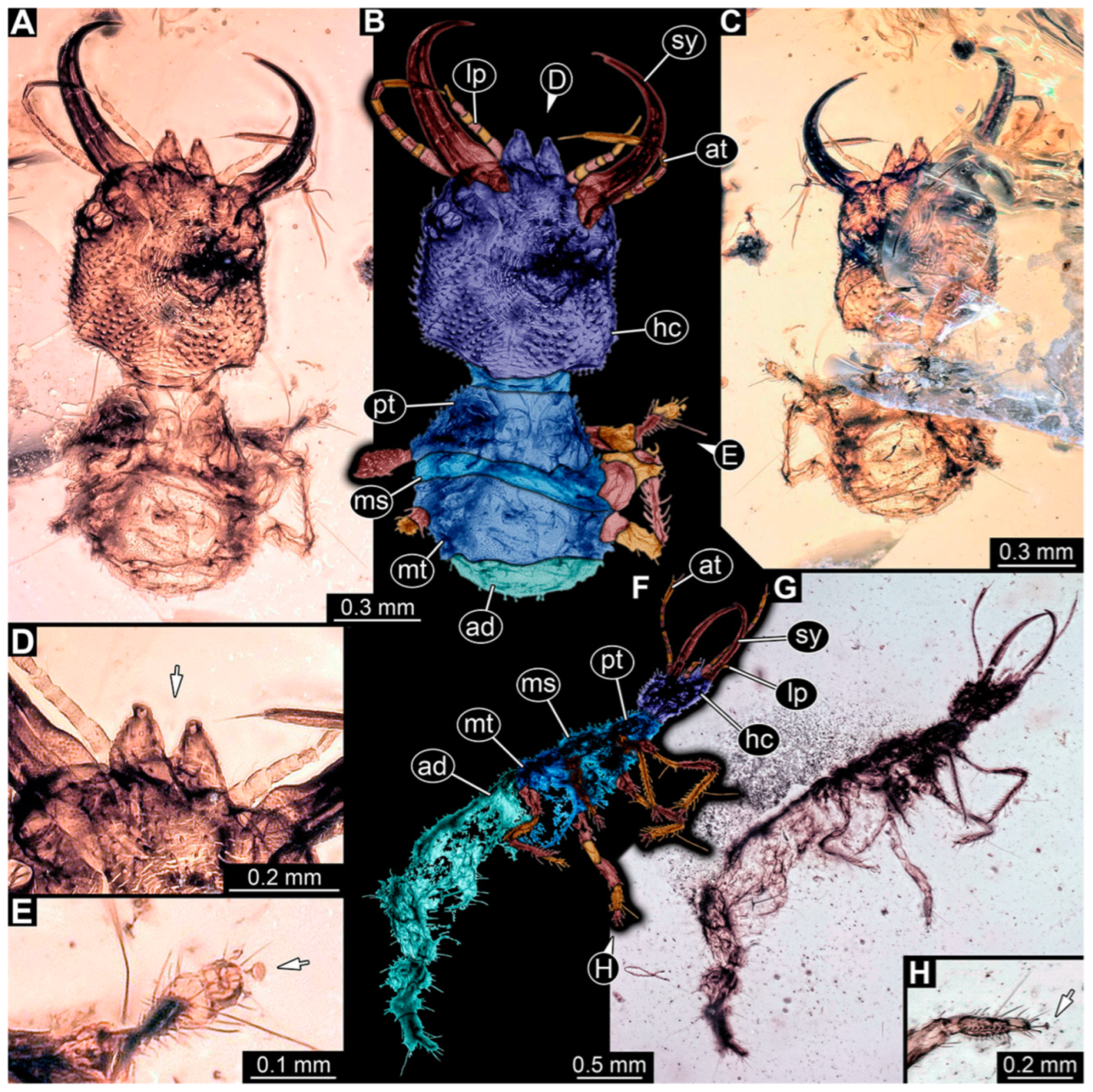
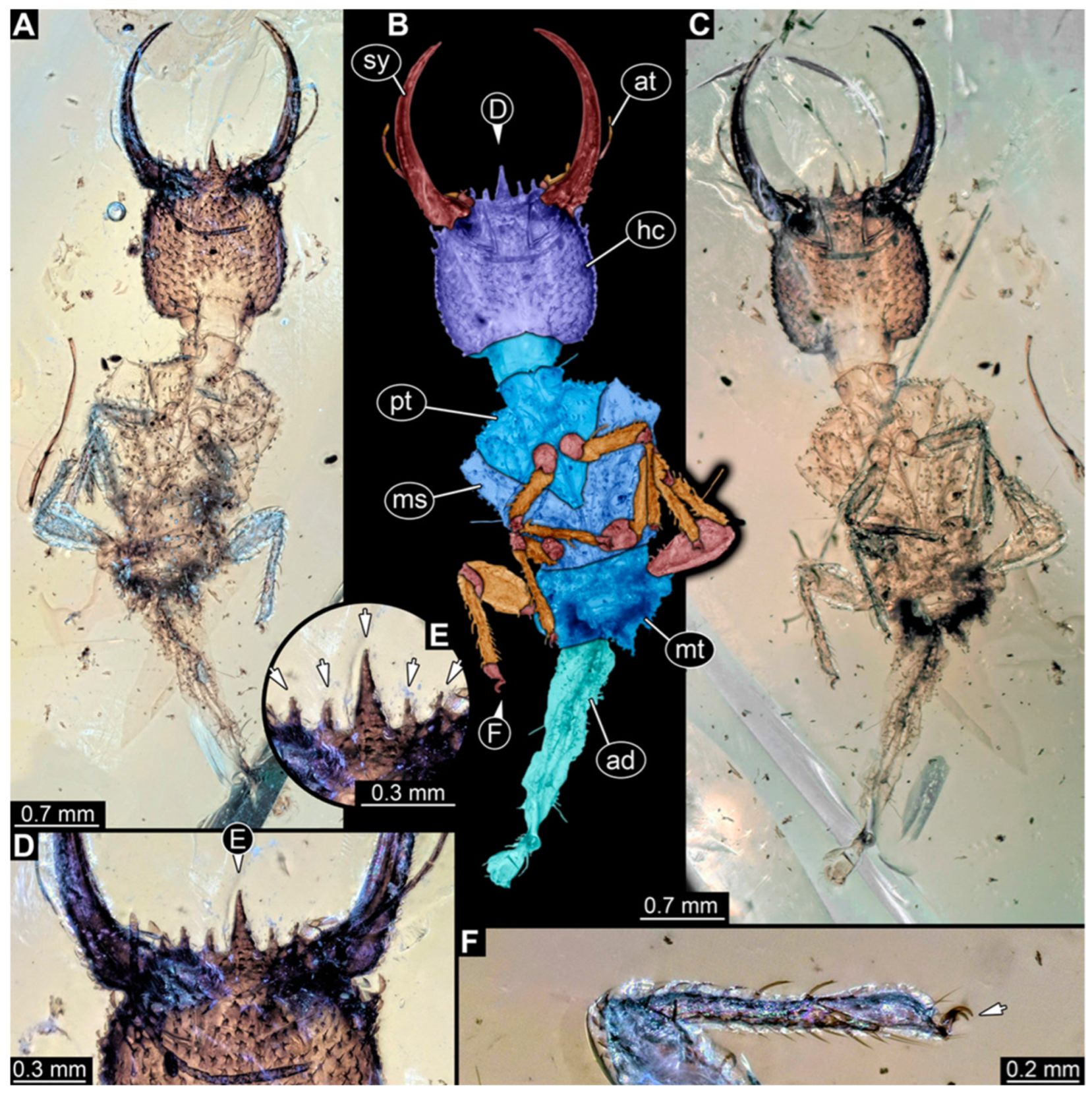
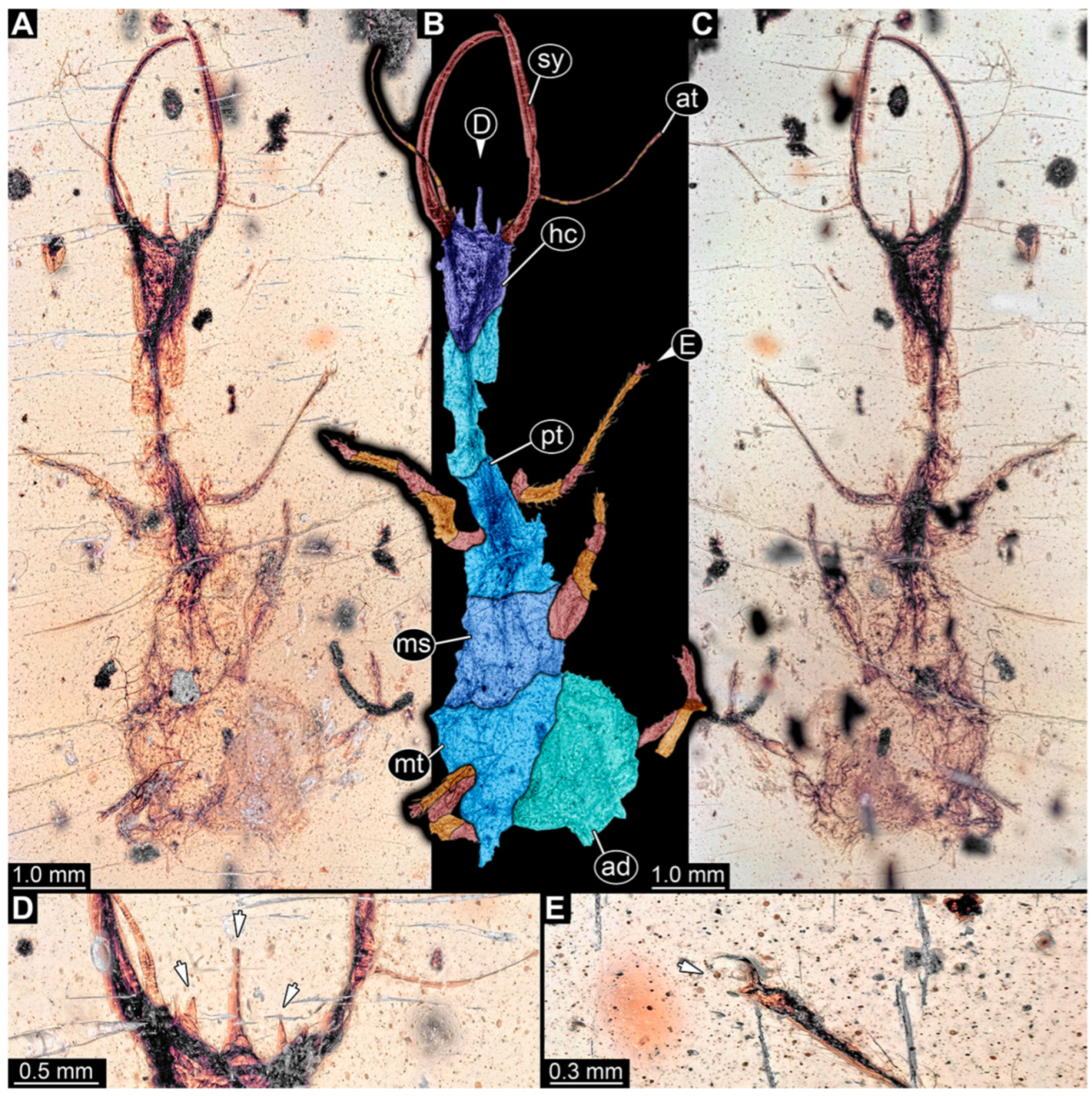
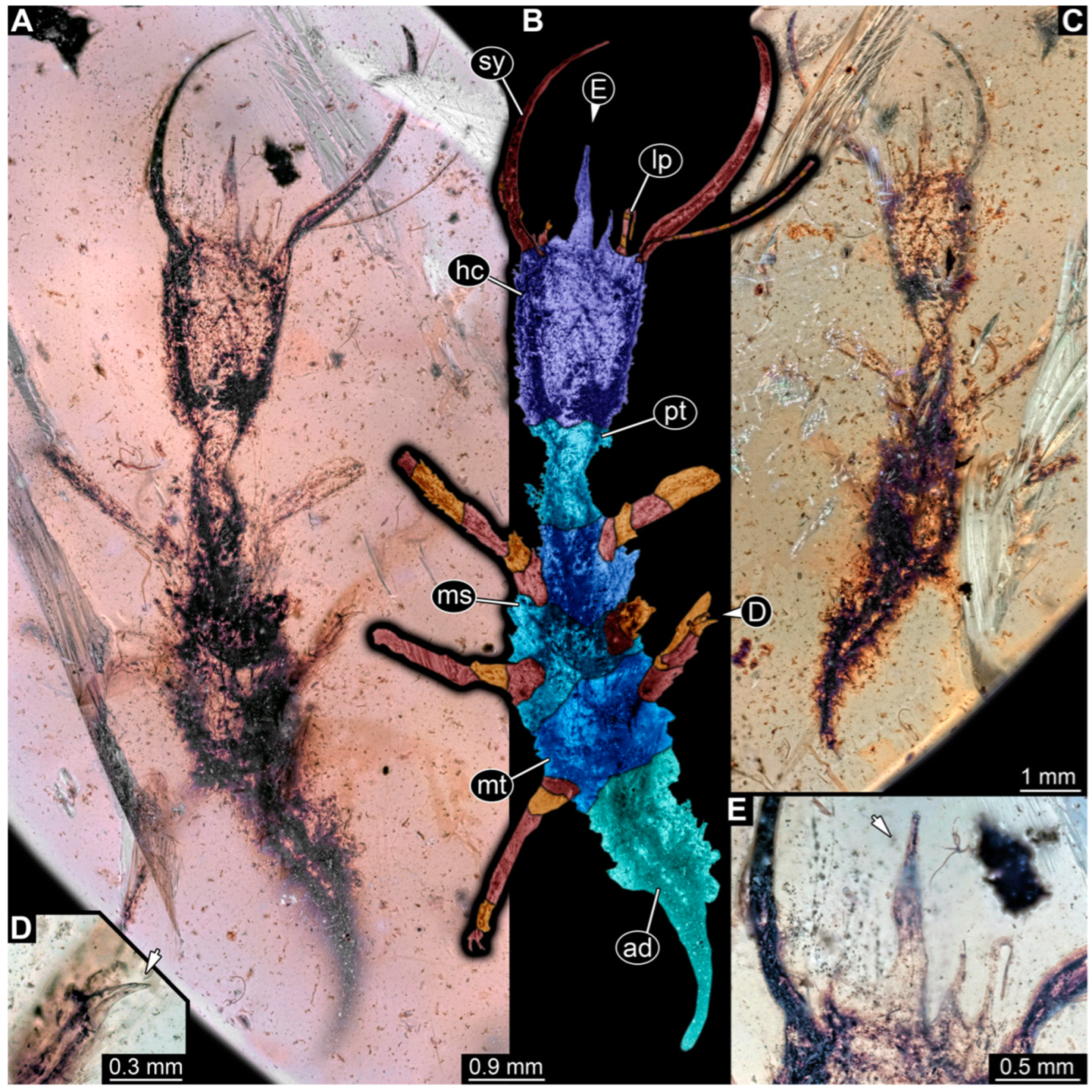
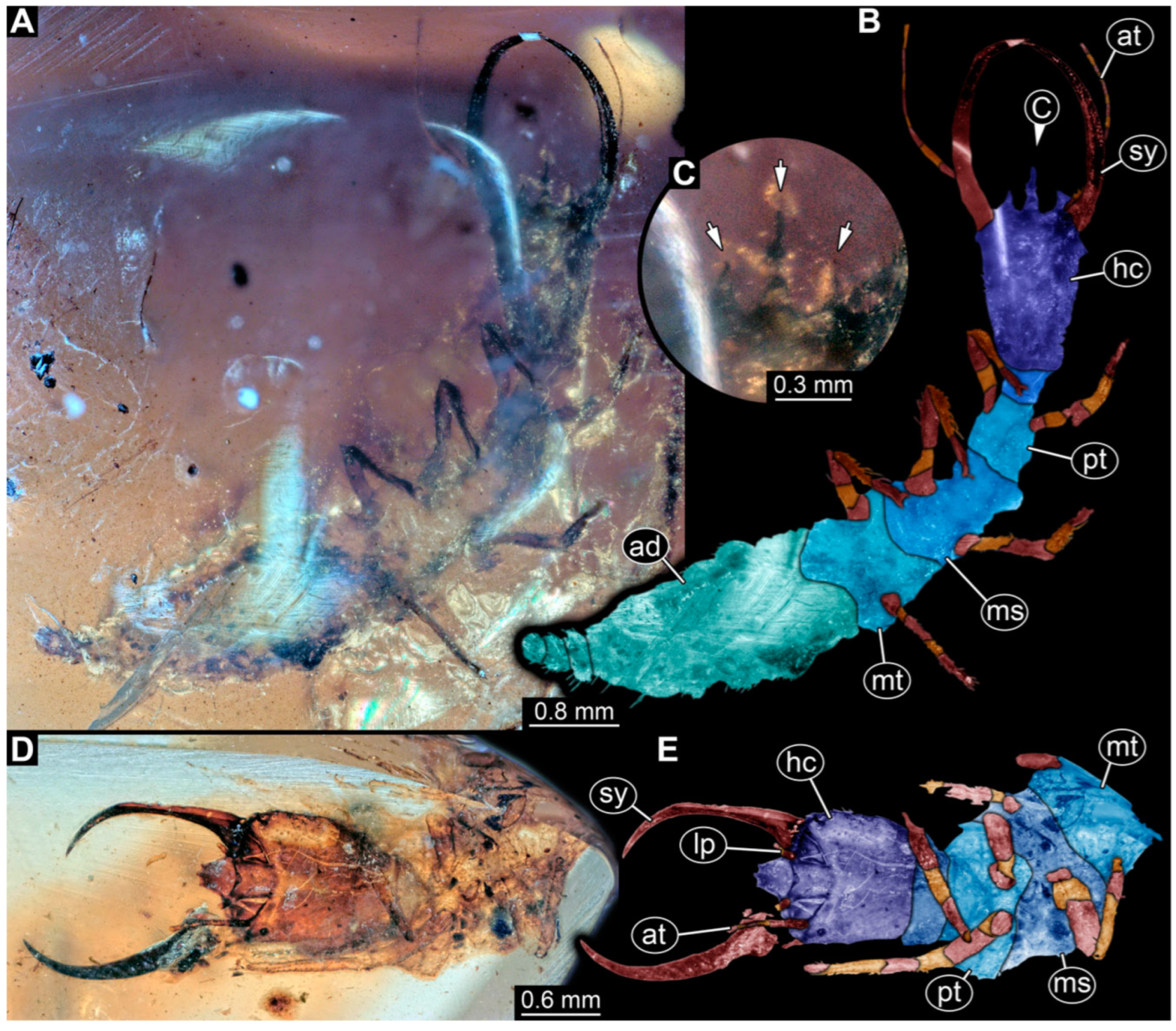
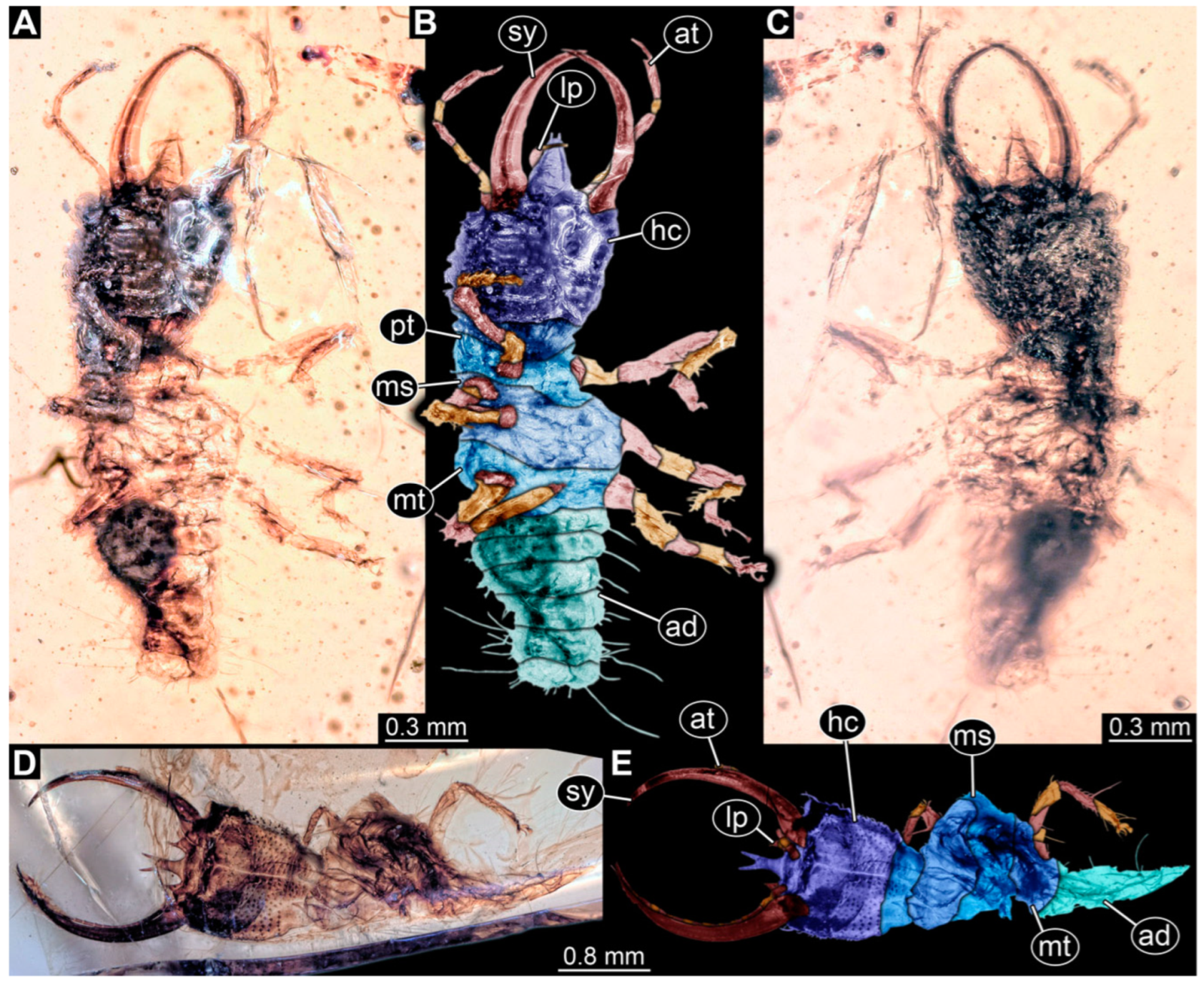
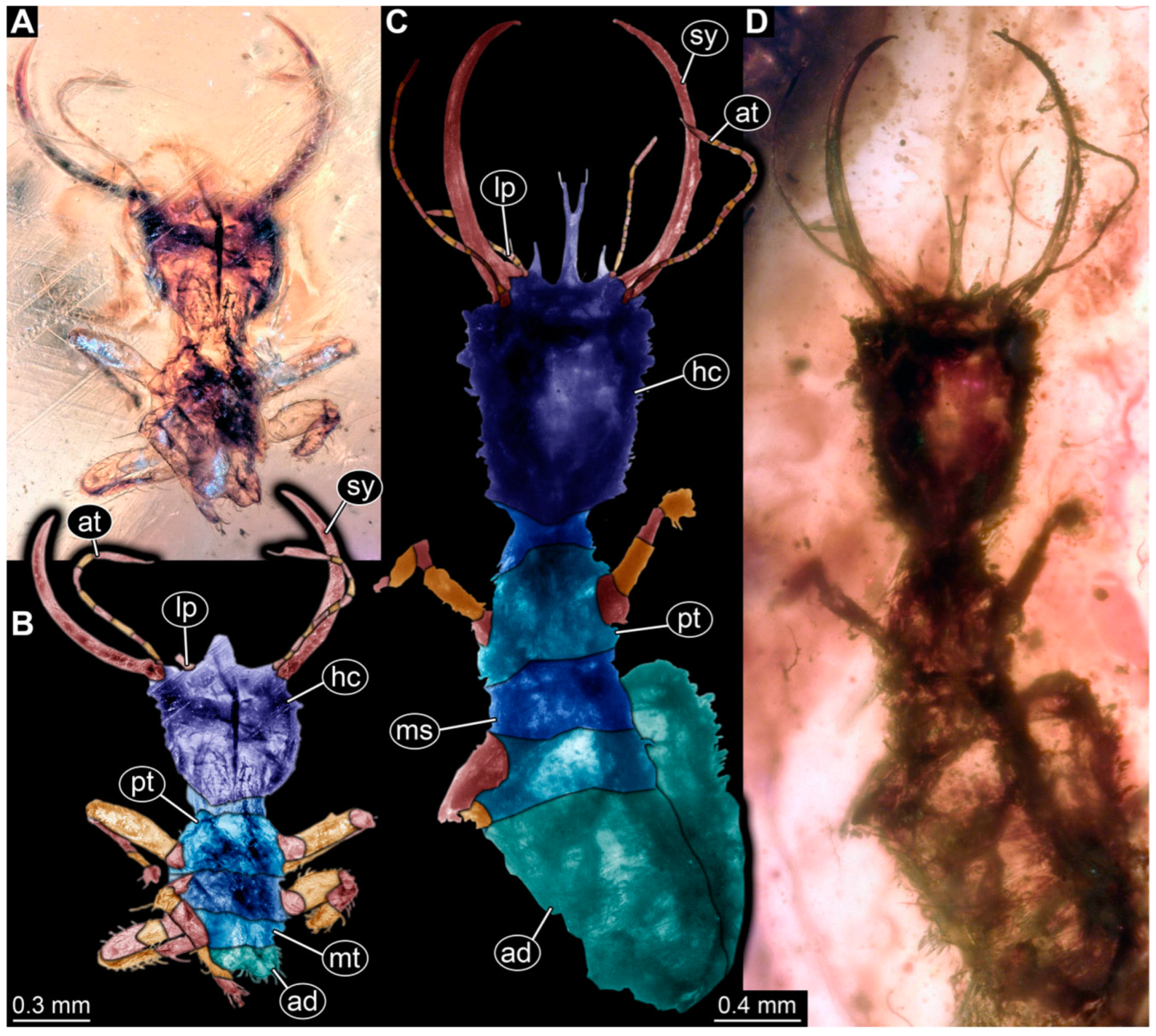
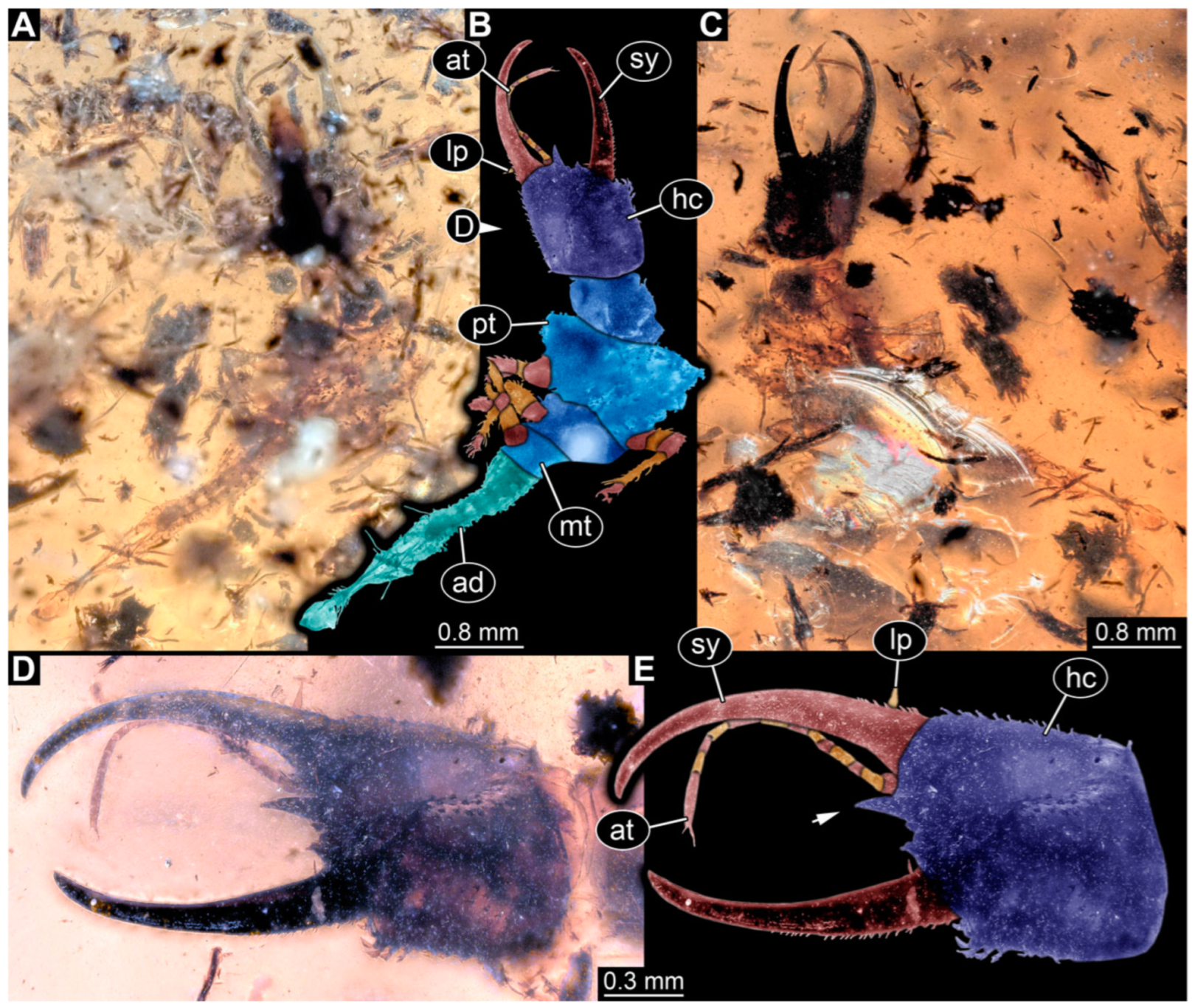
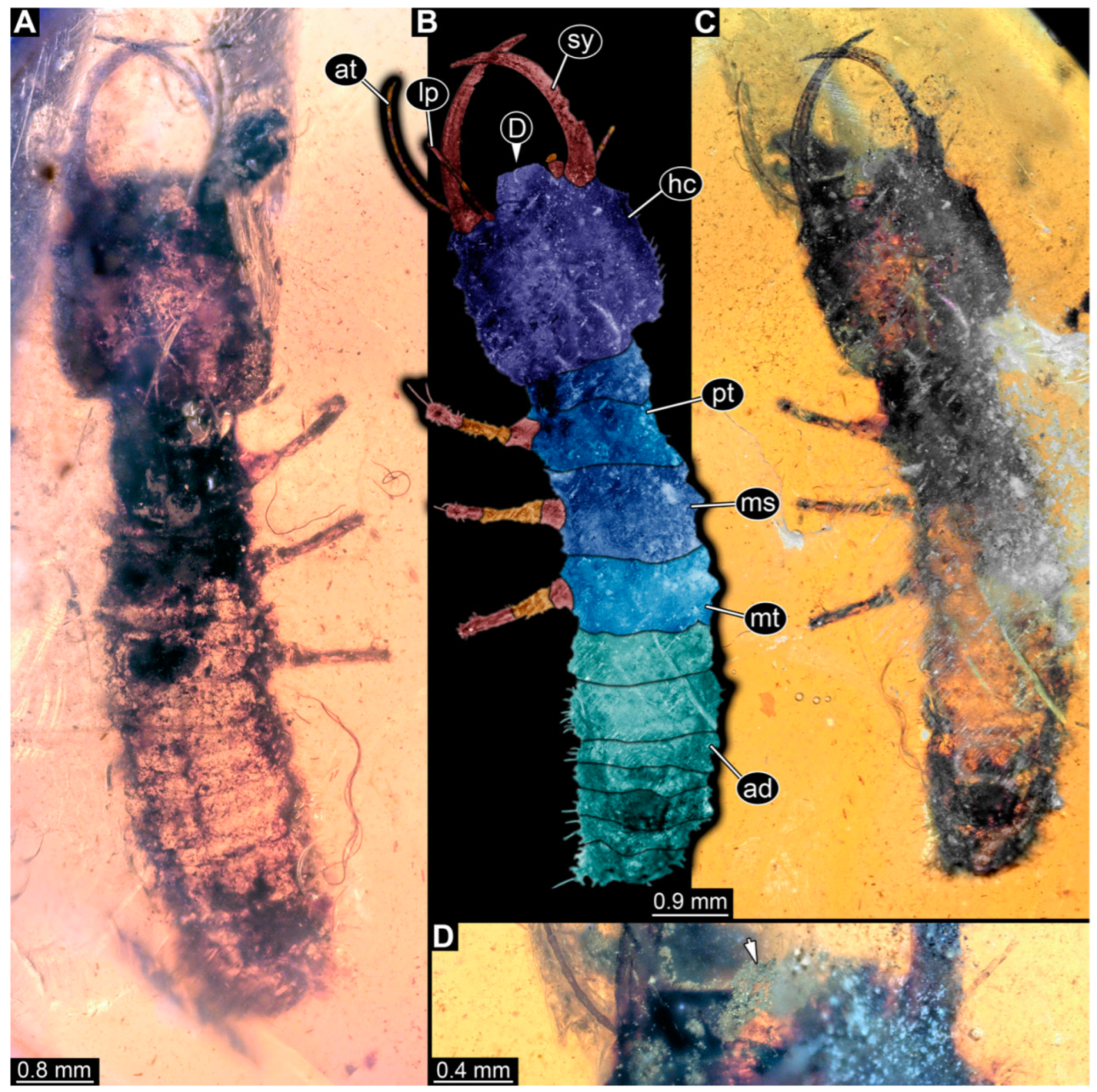
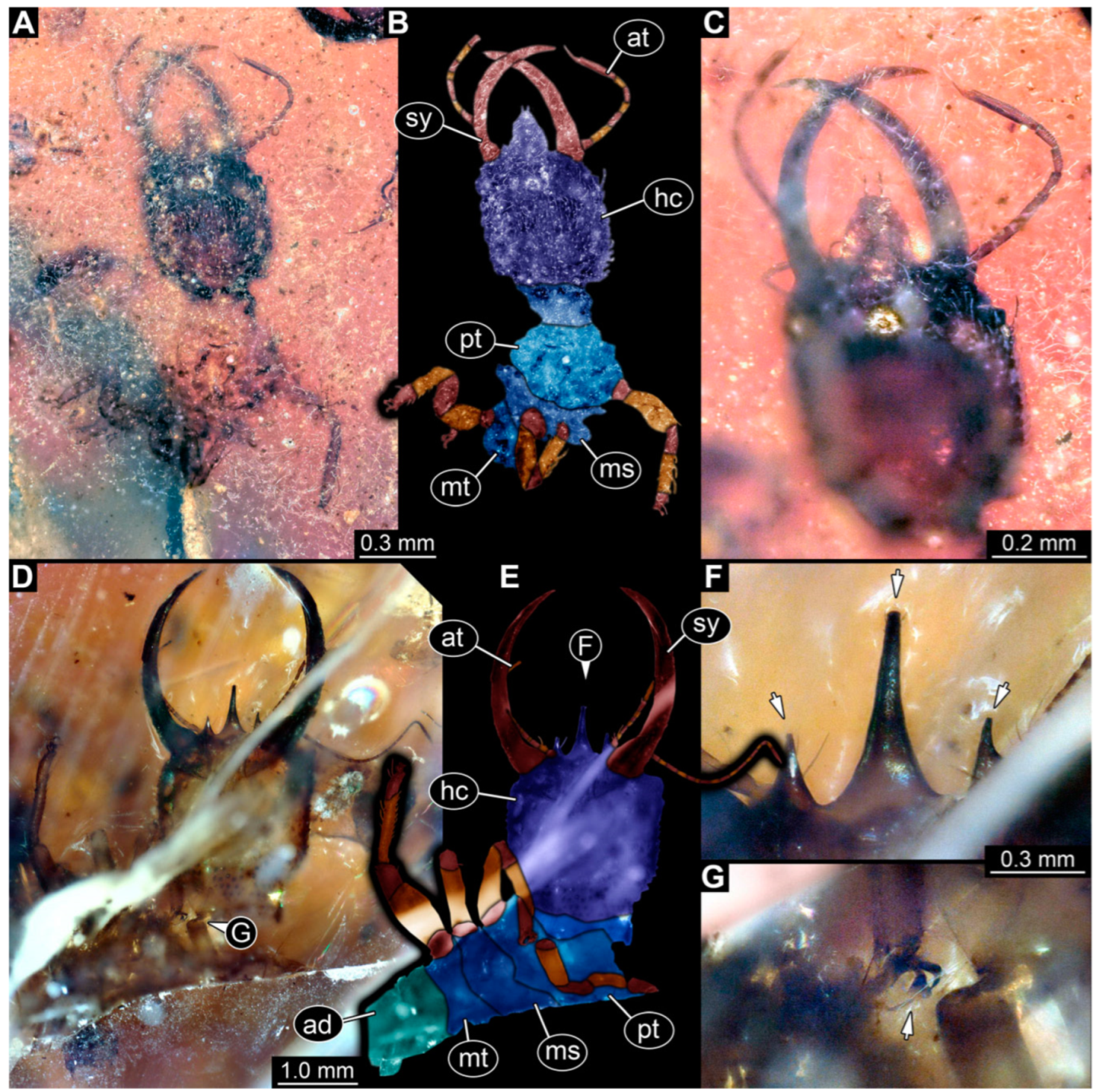
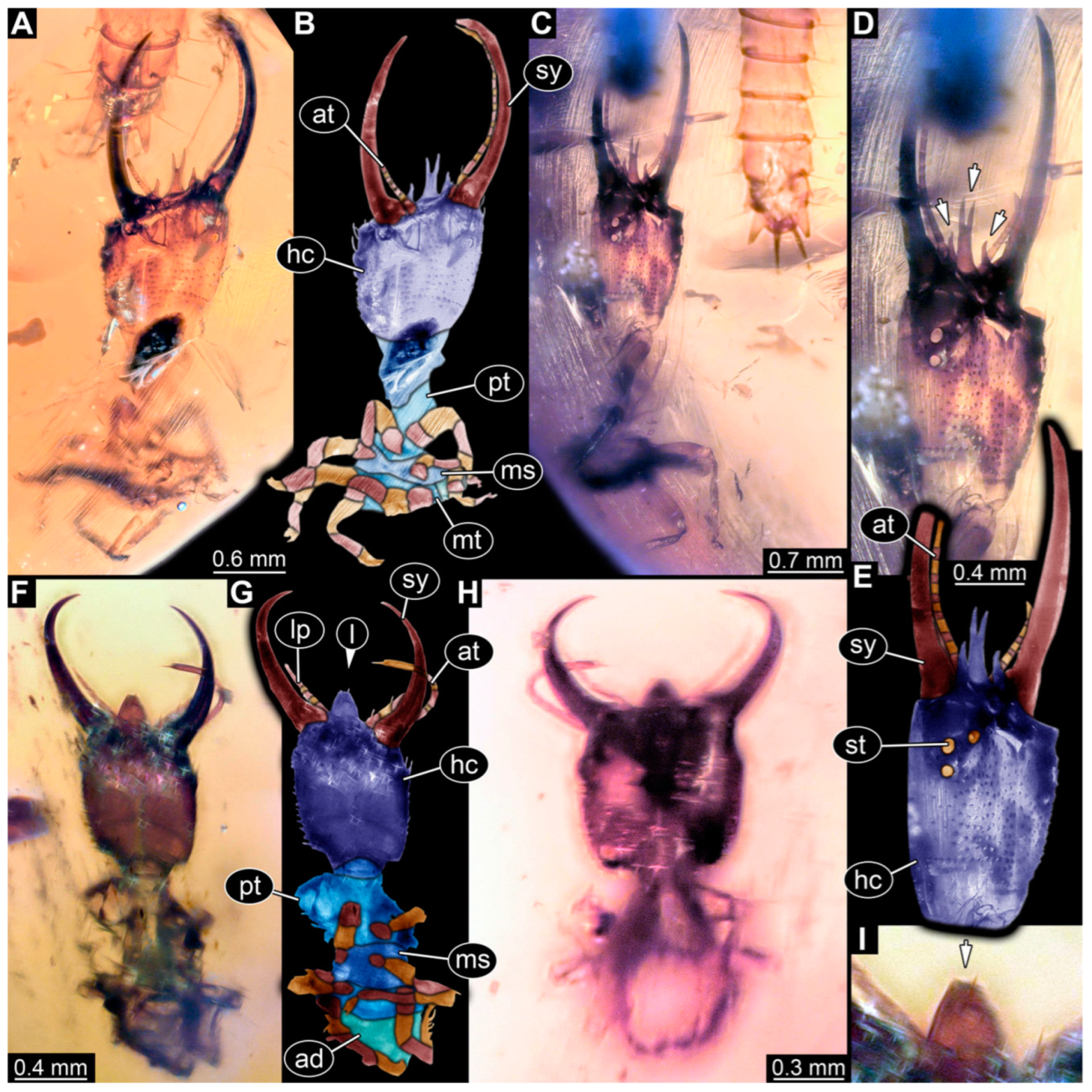
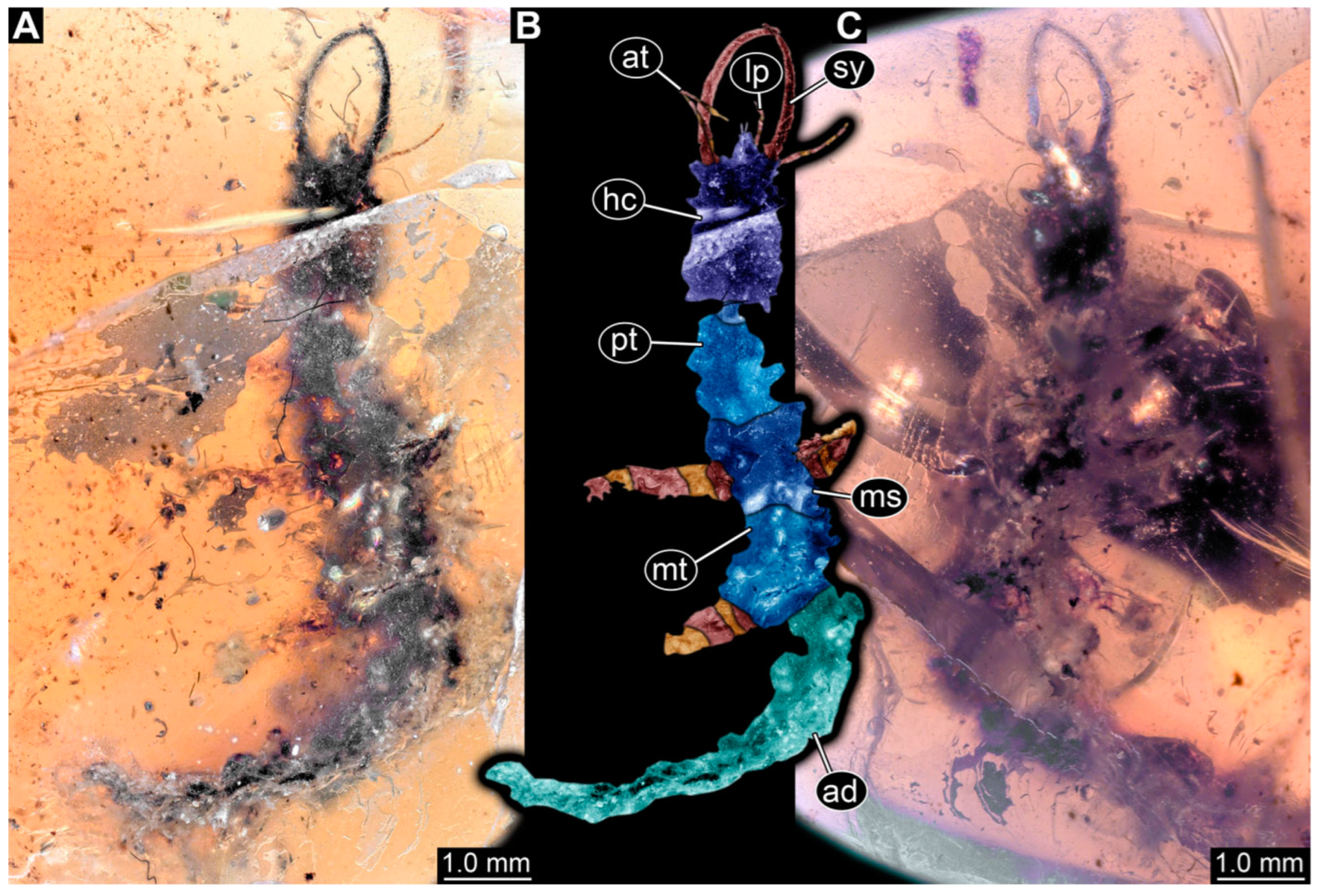
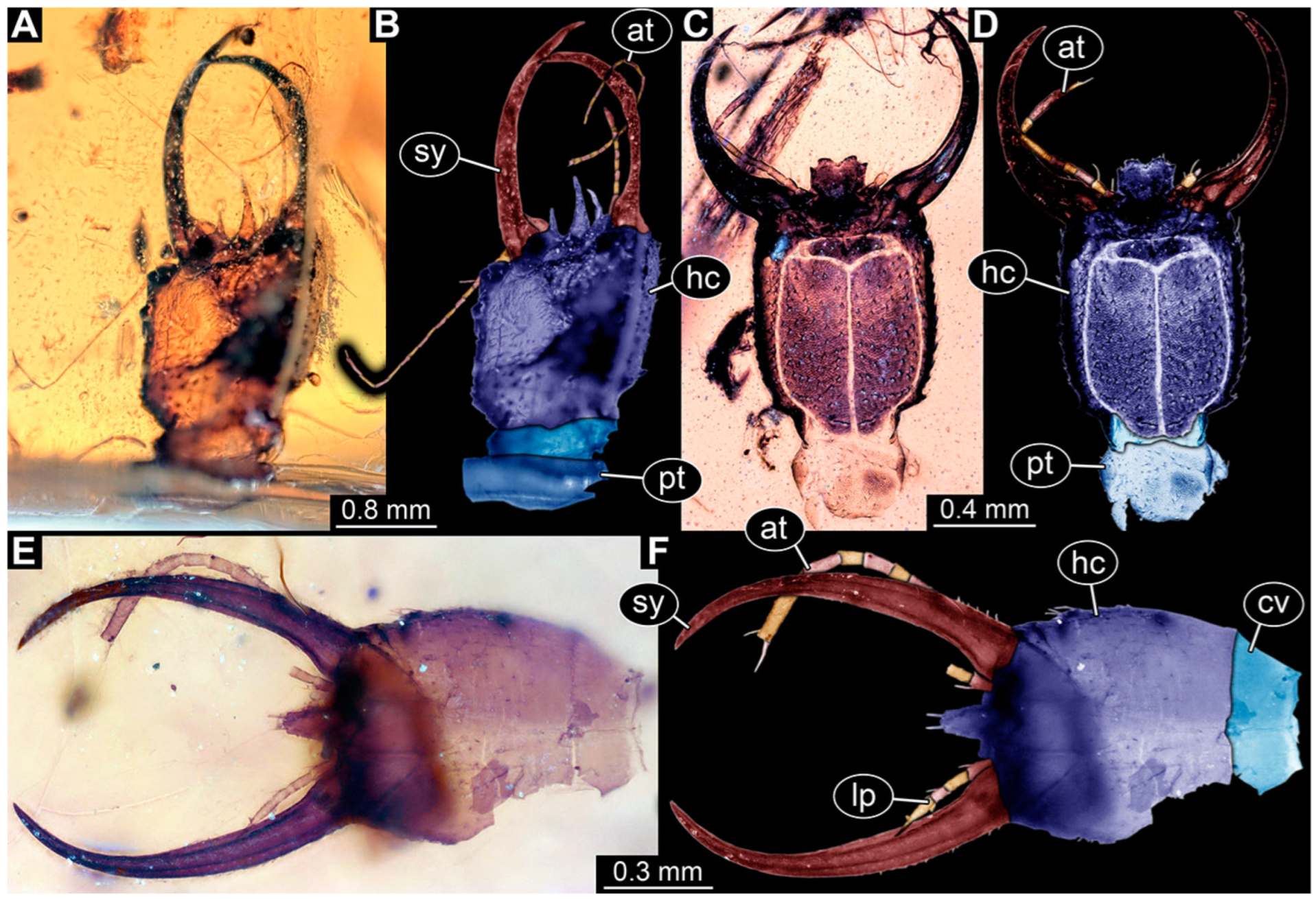
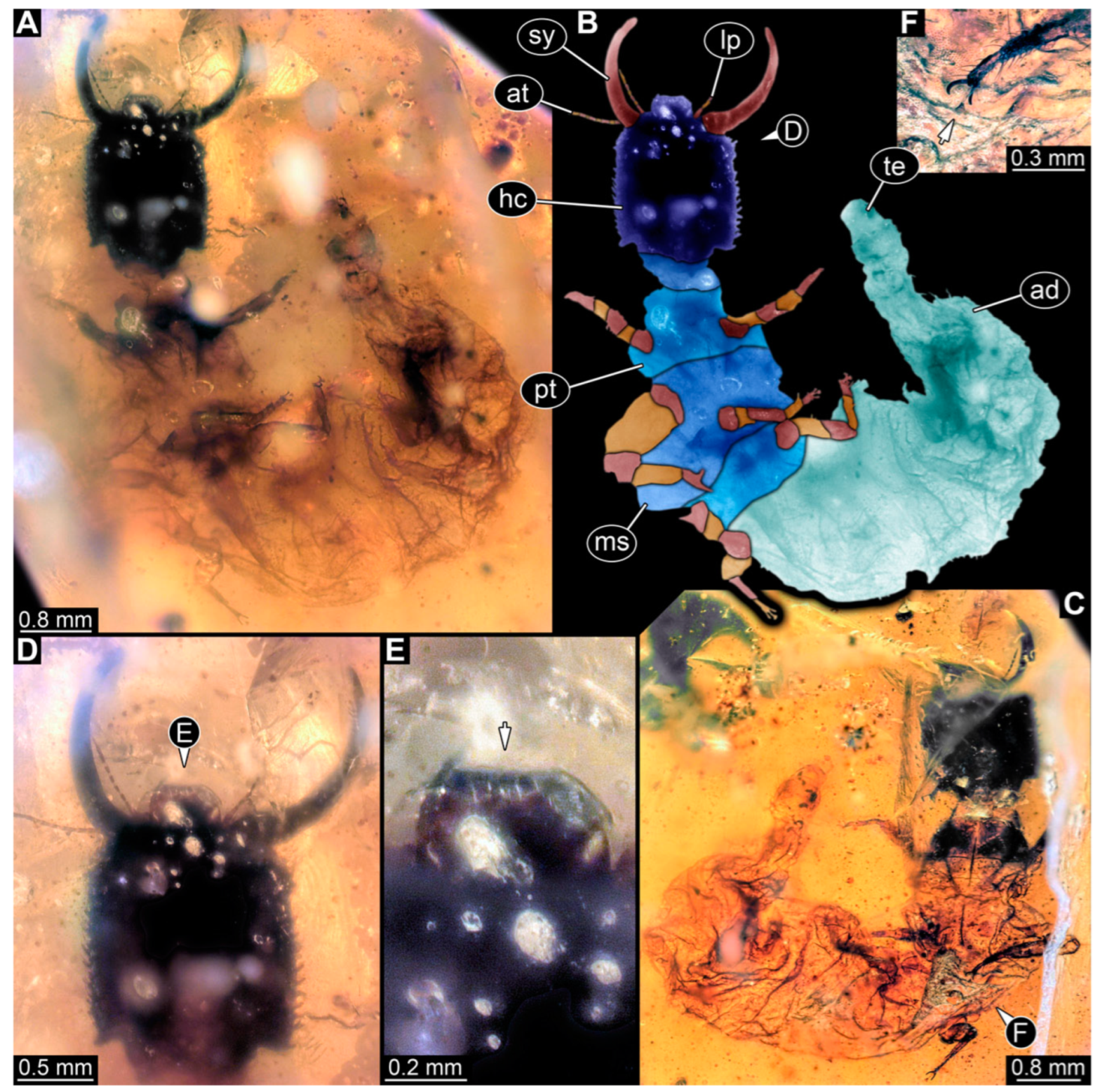
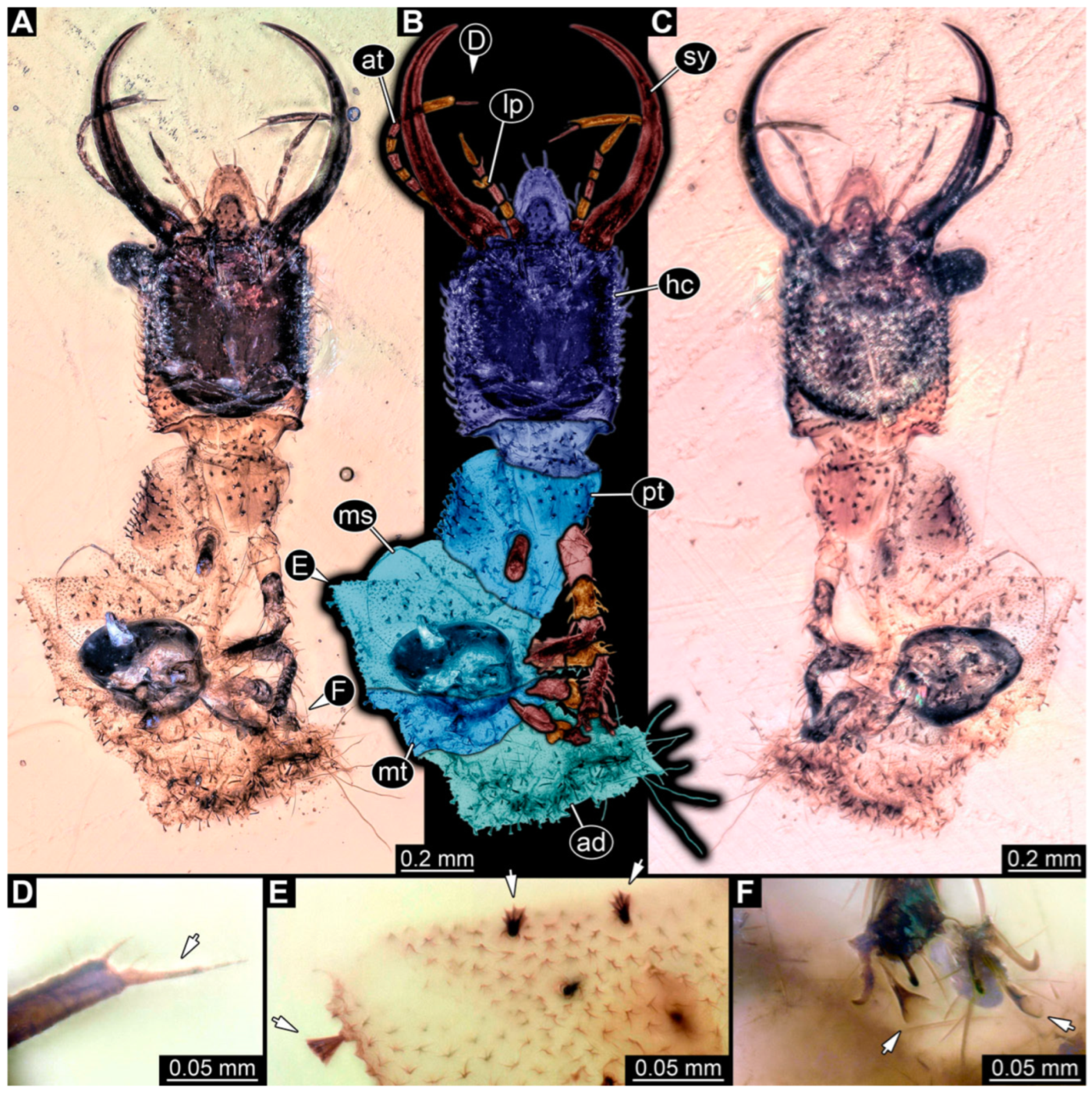
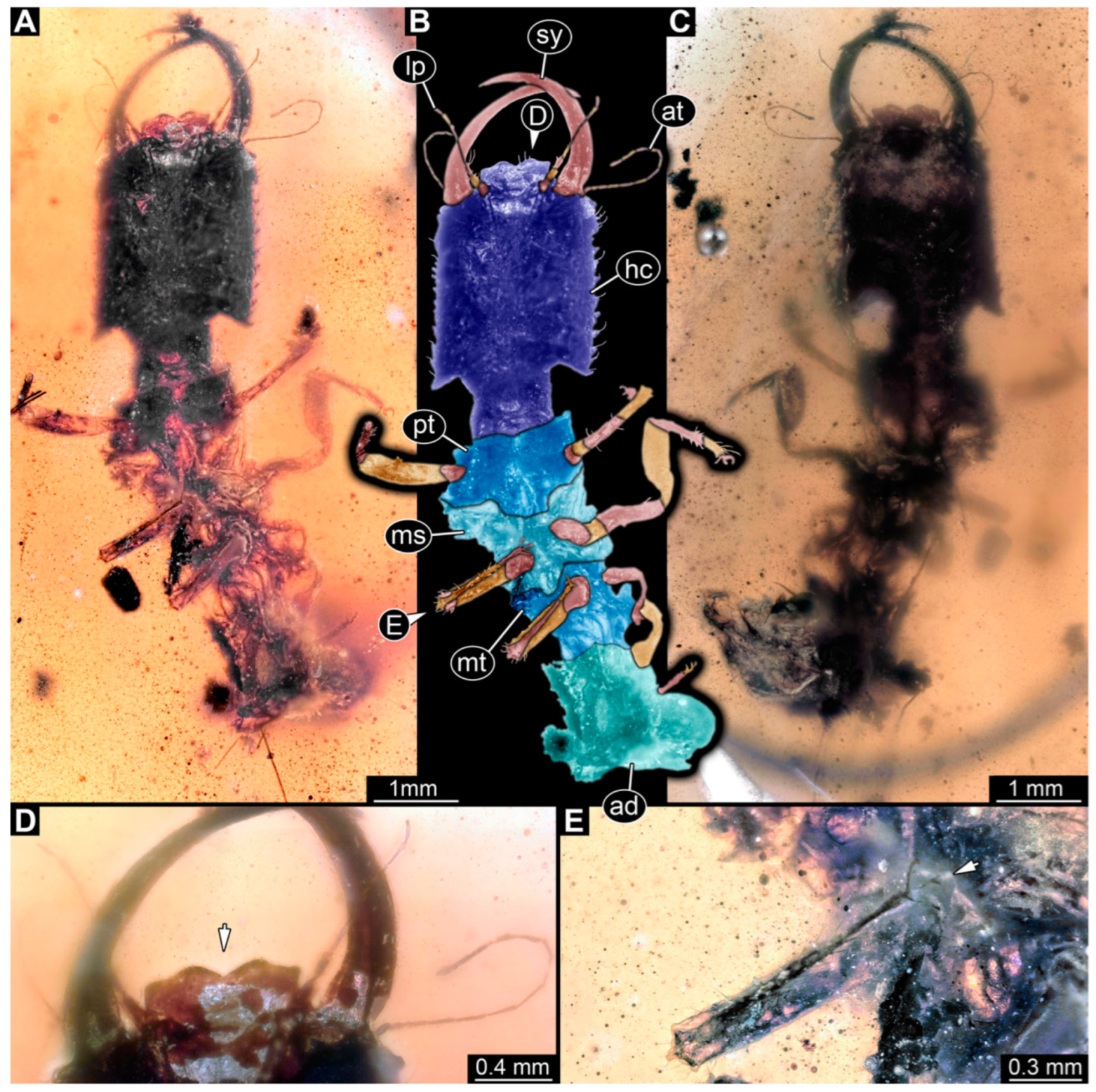
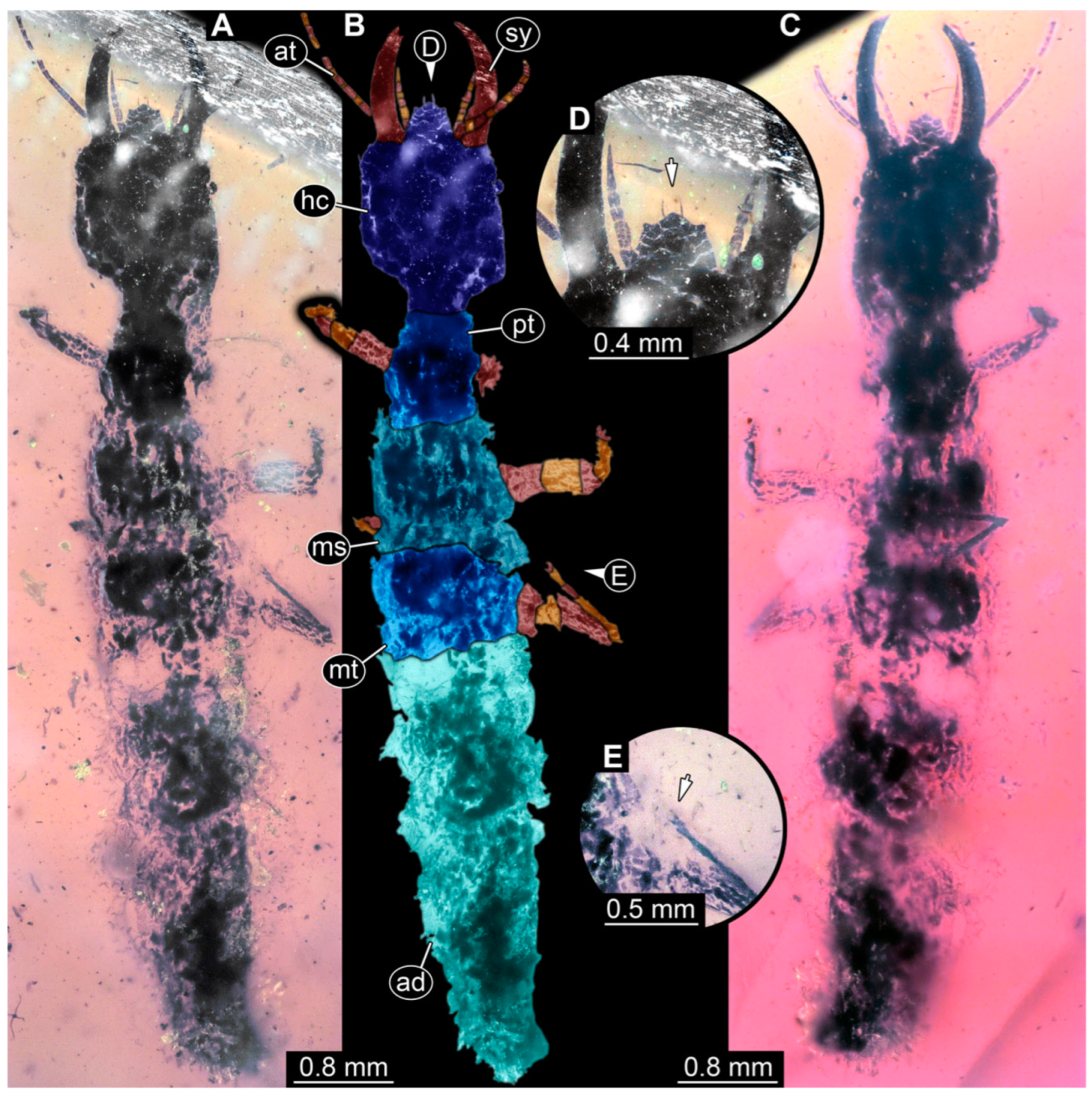
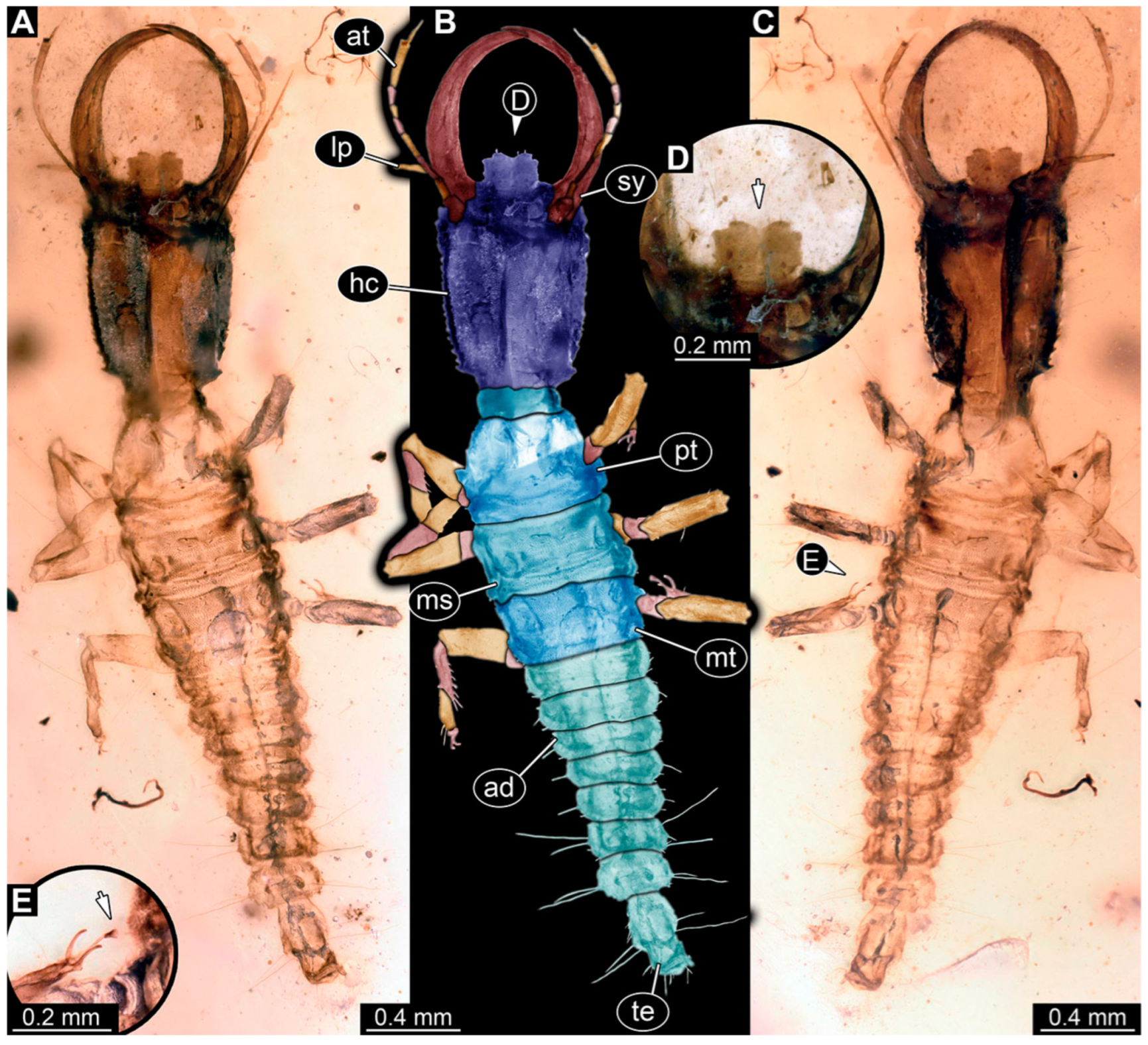
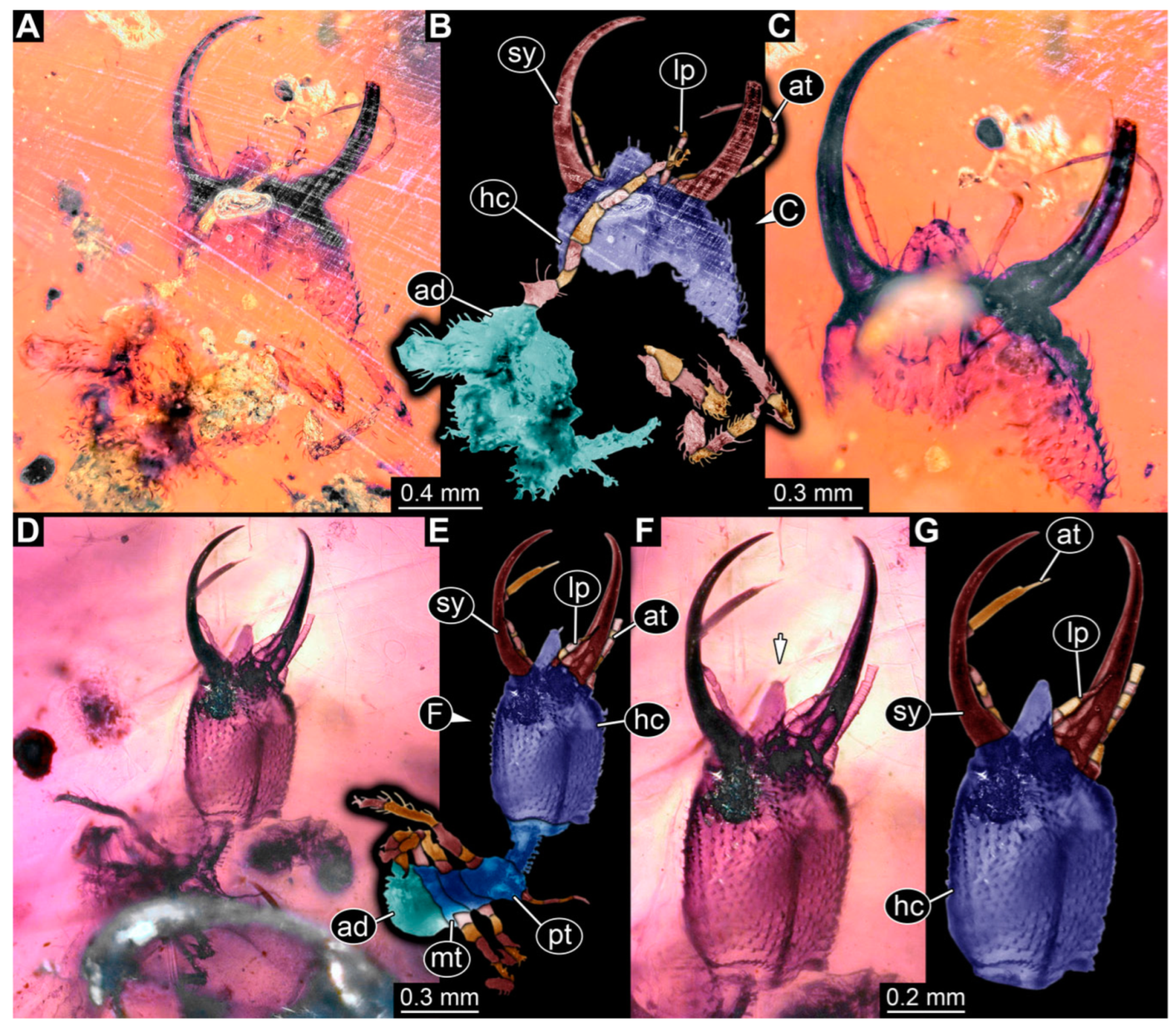
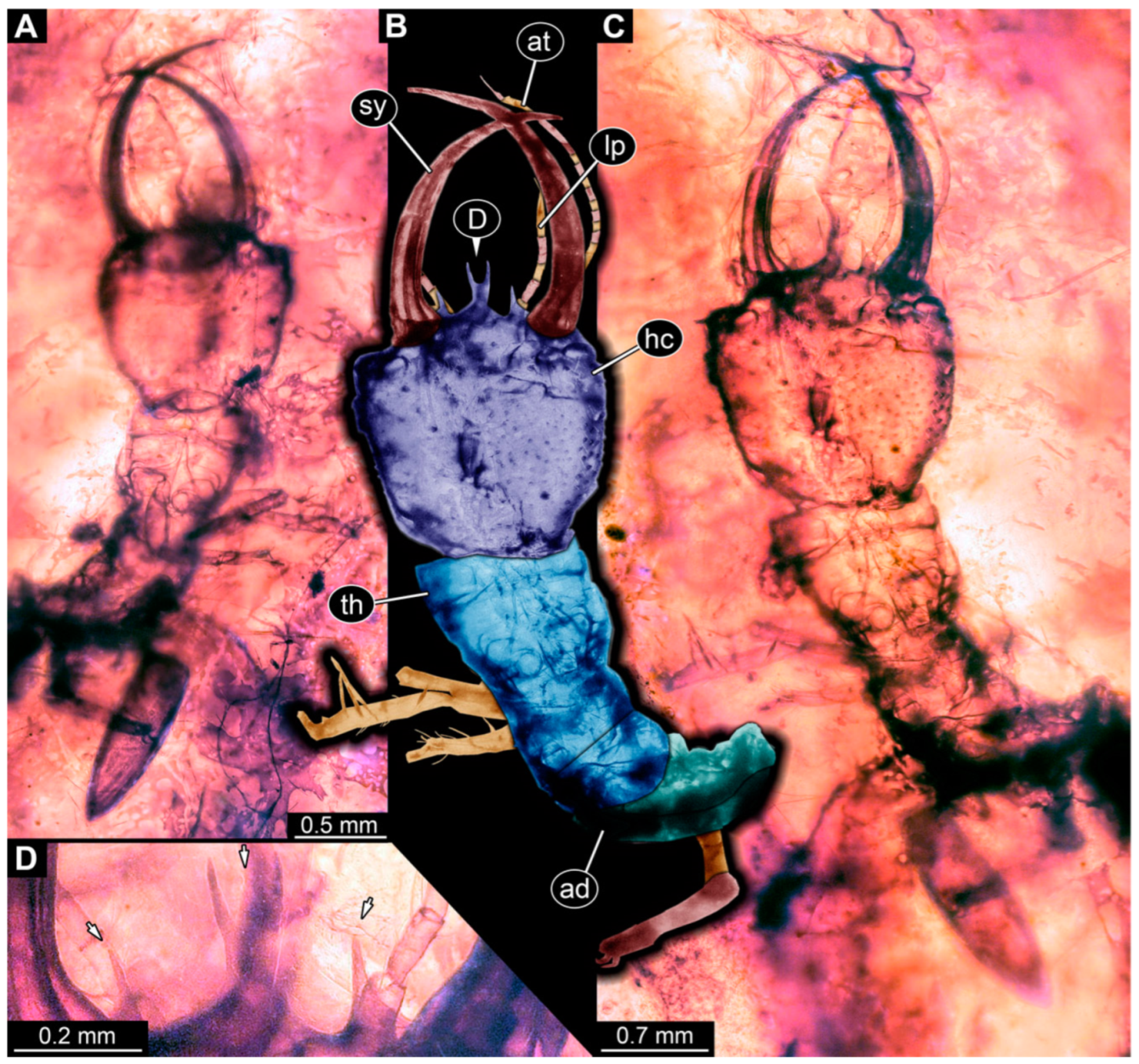
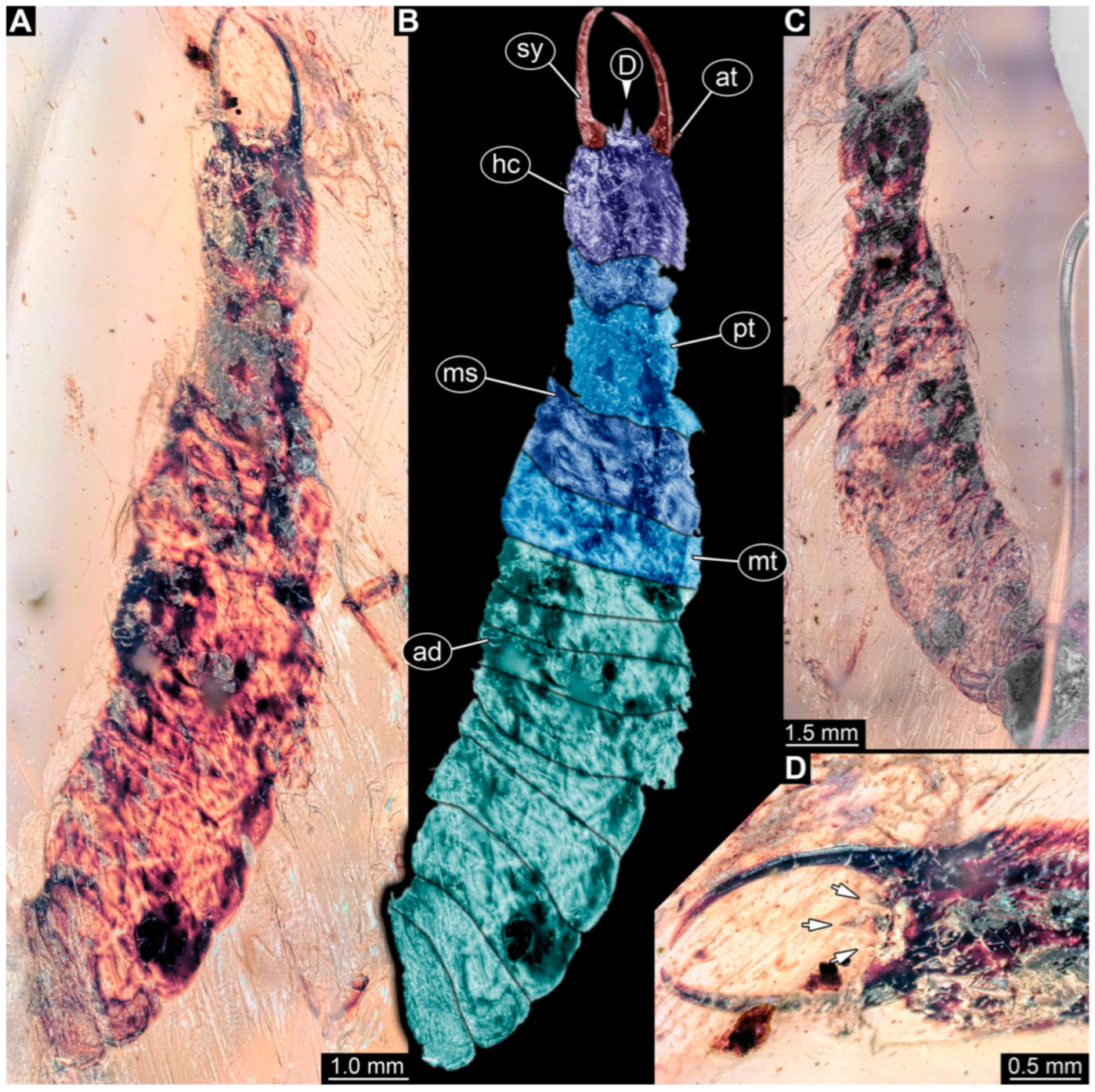
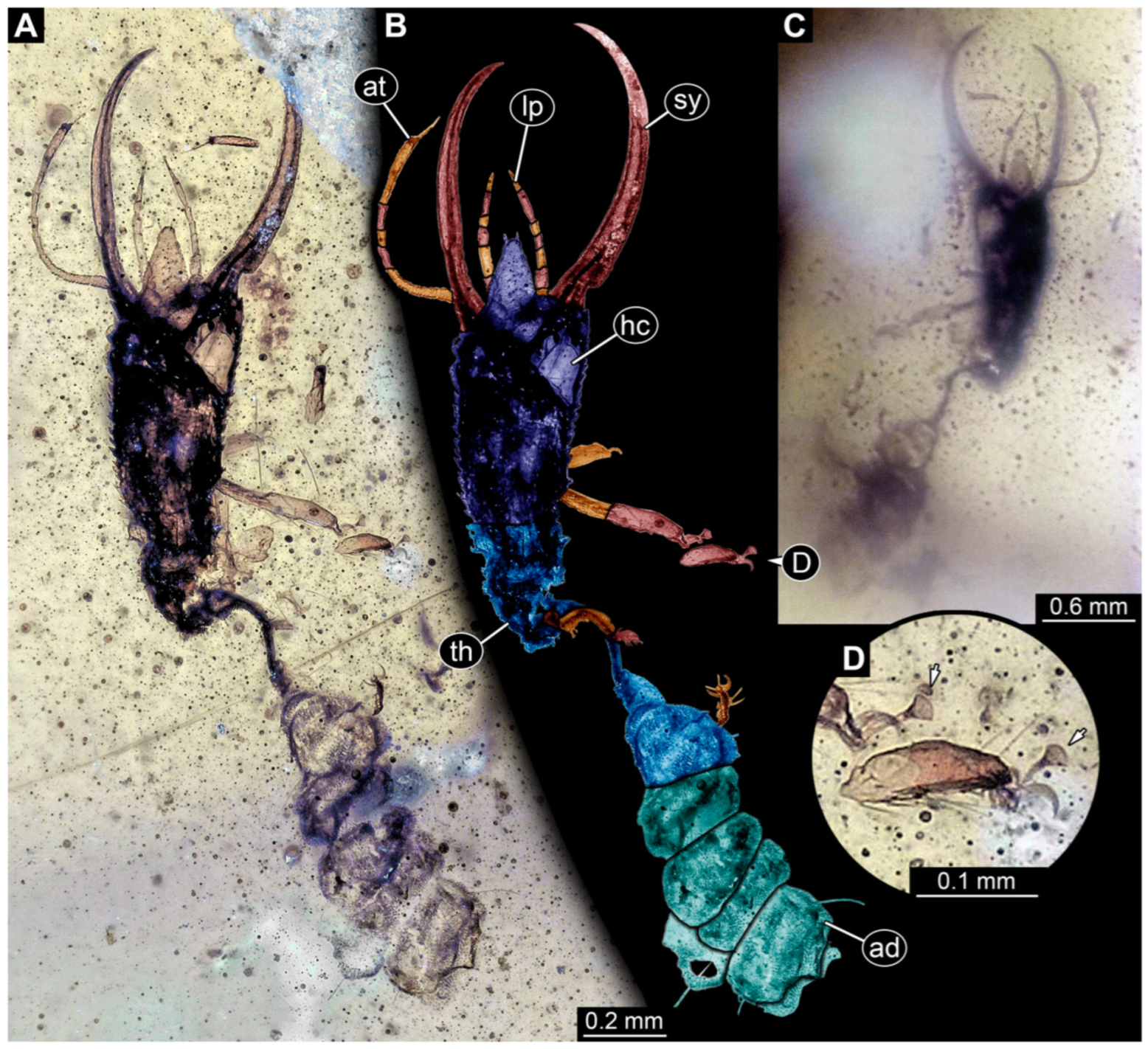
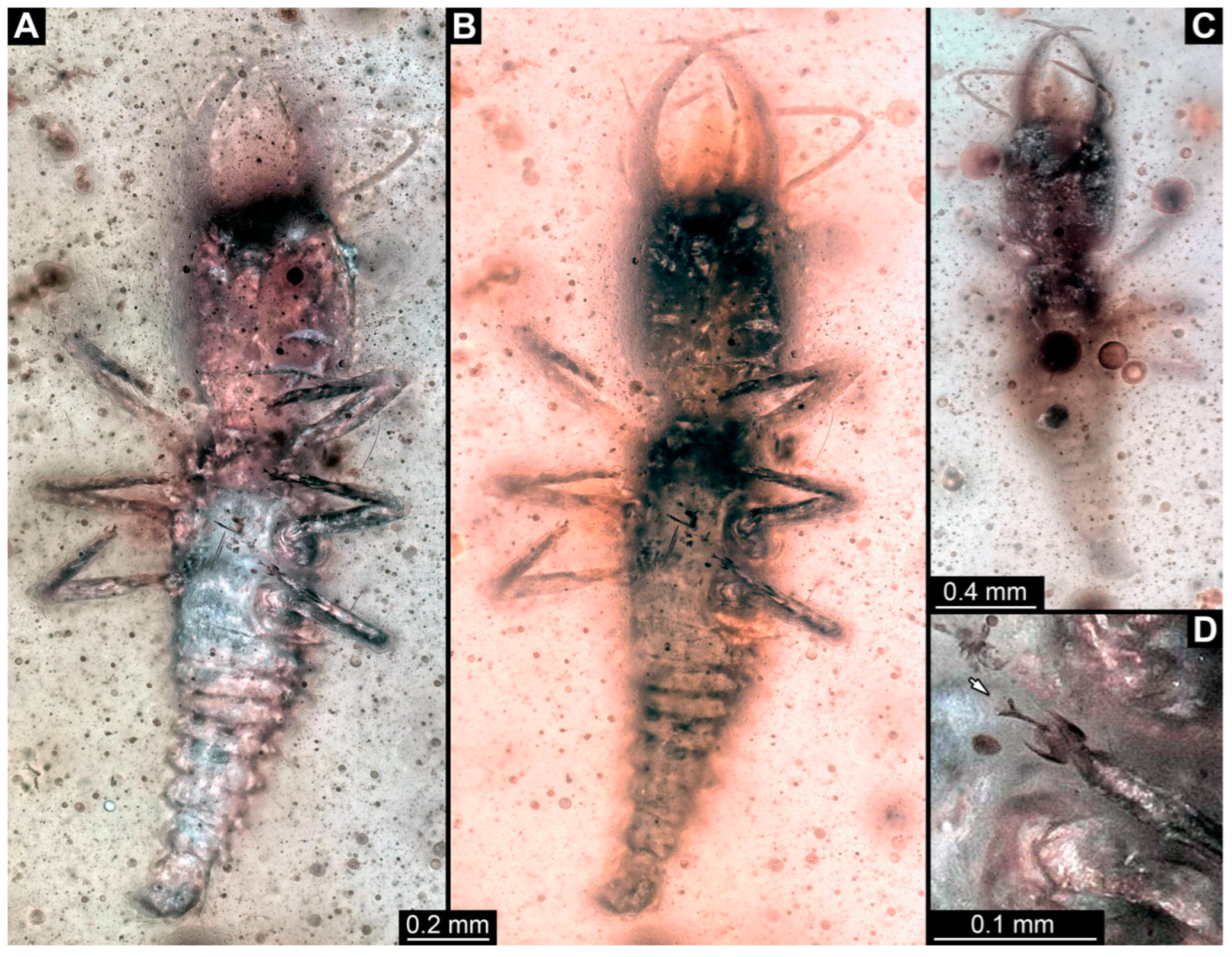
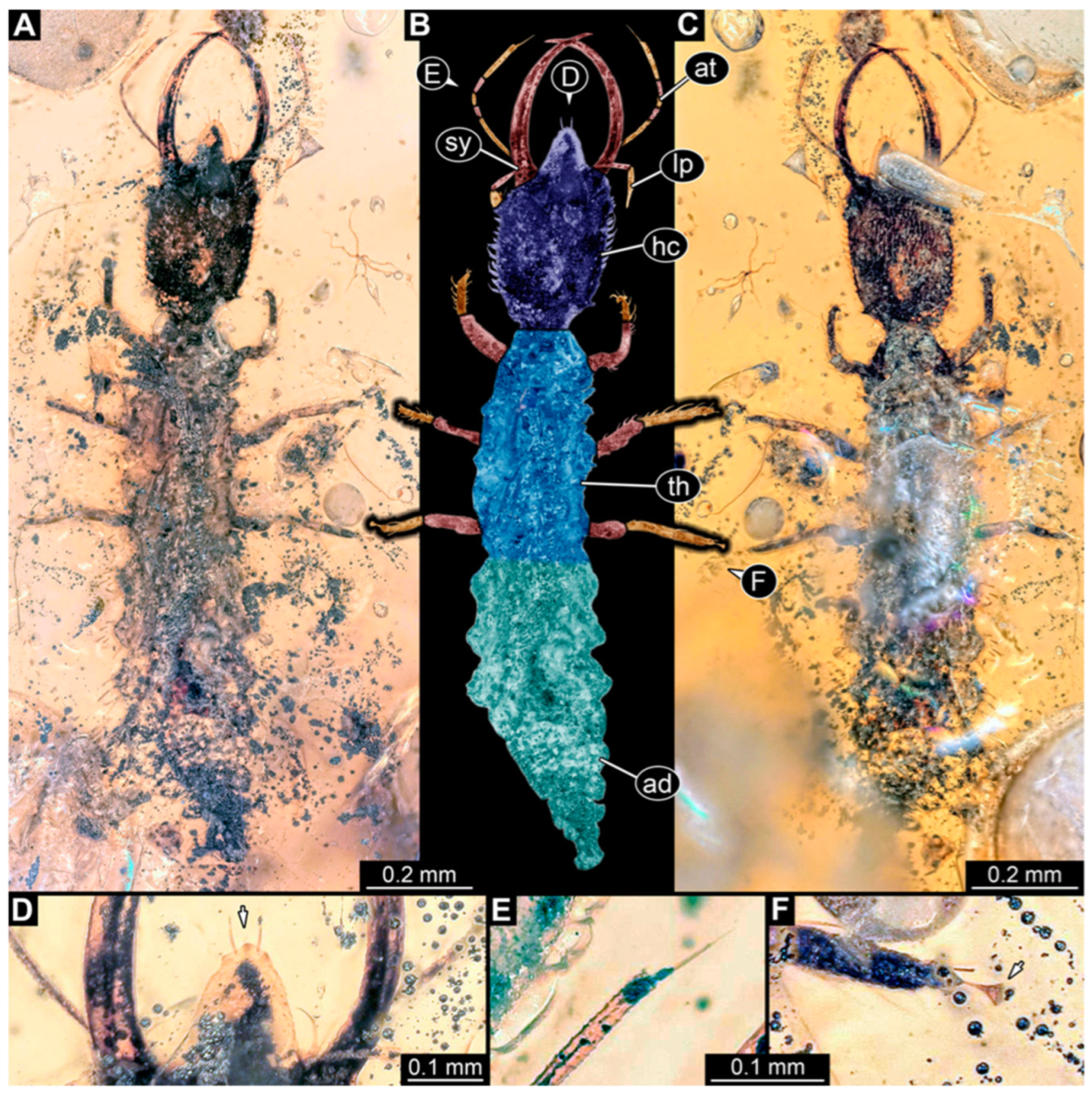
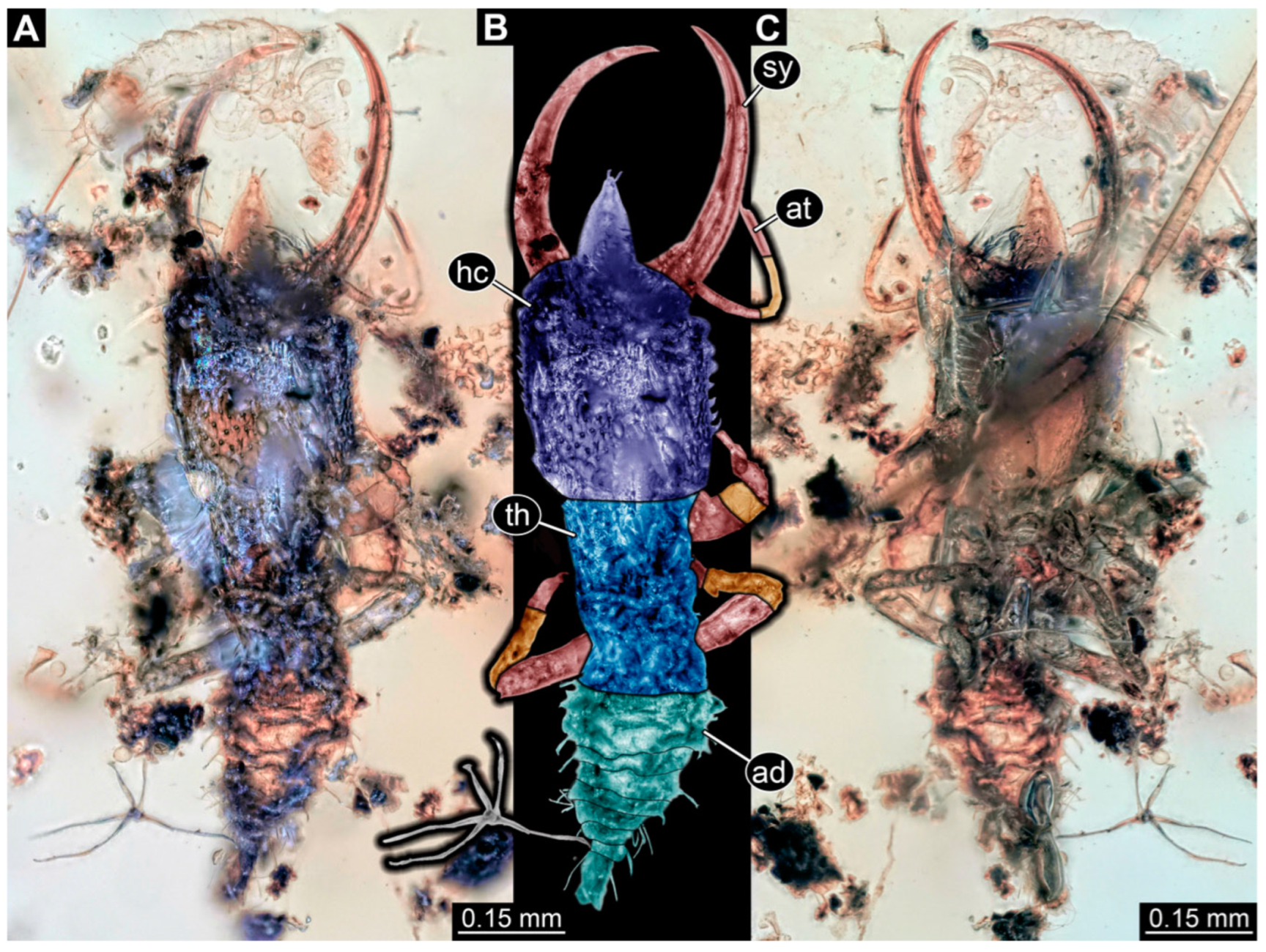
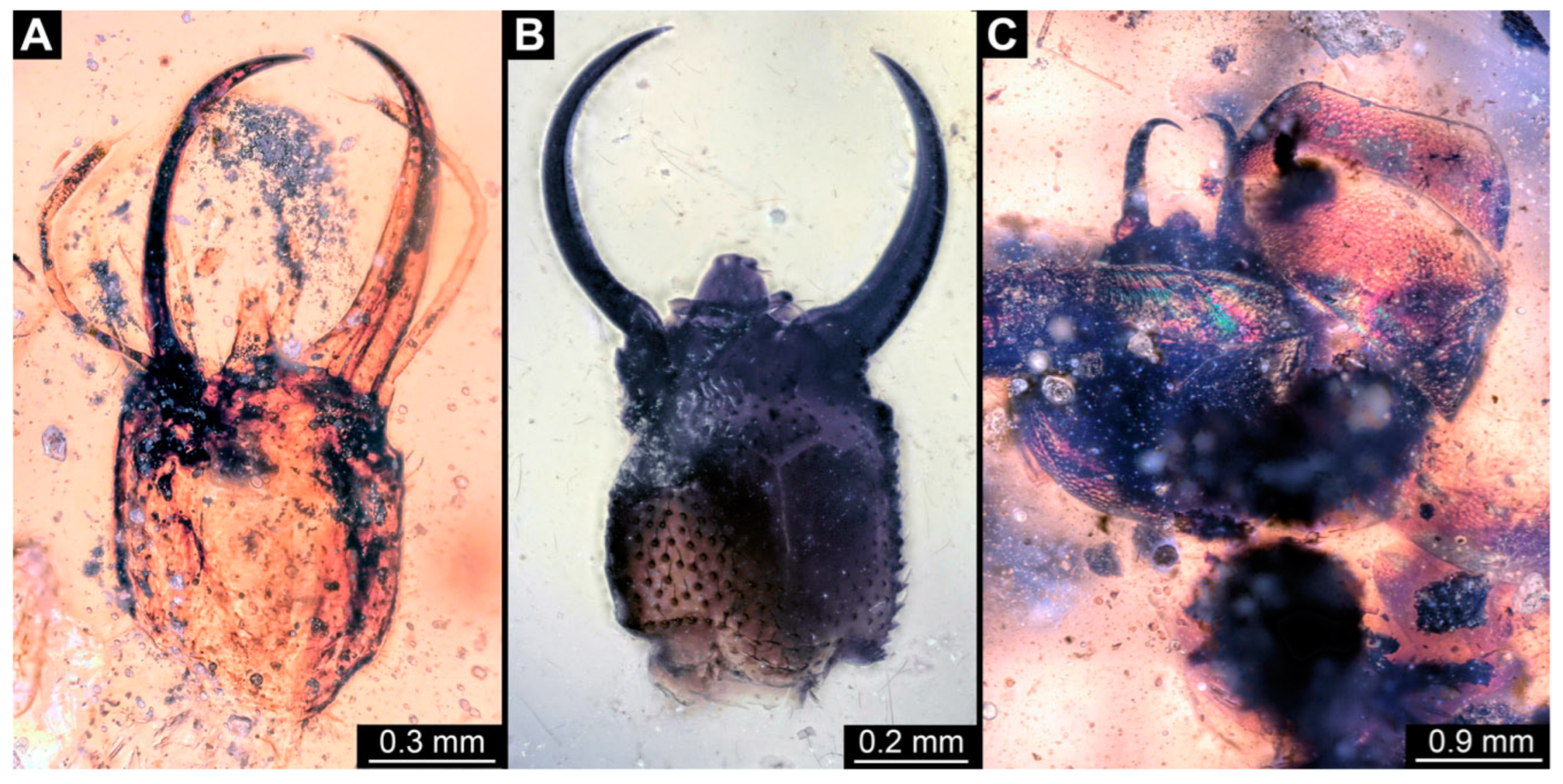
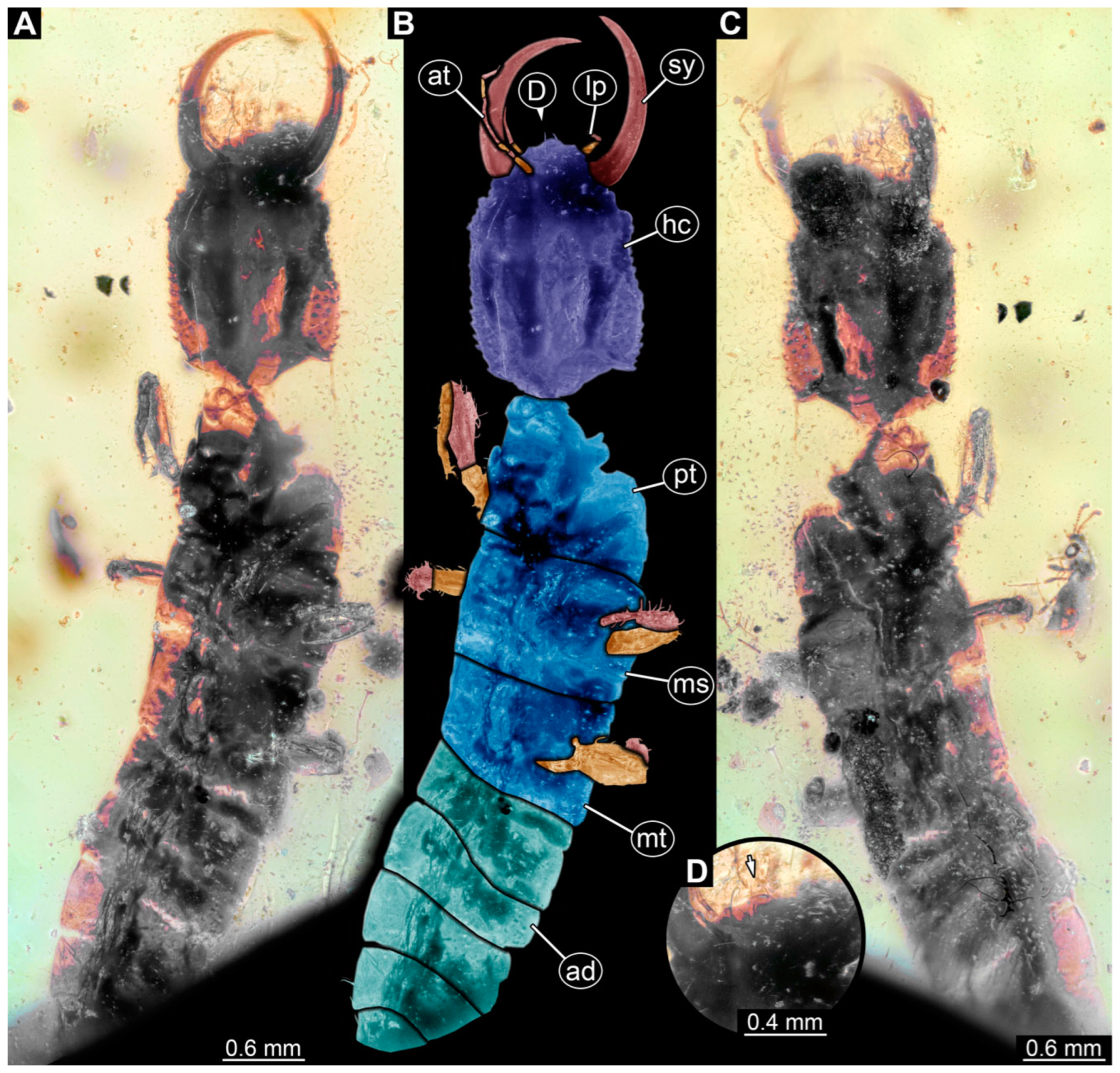
3.1. Descriptions of New Fossil Larvae
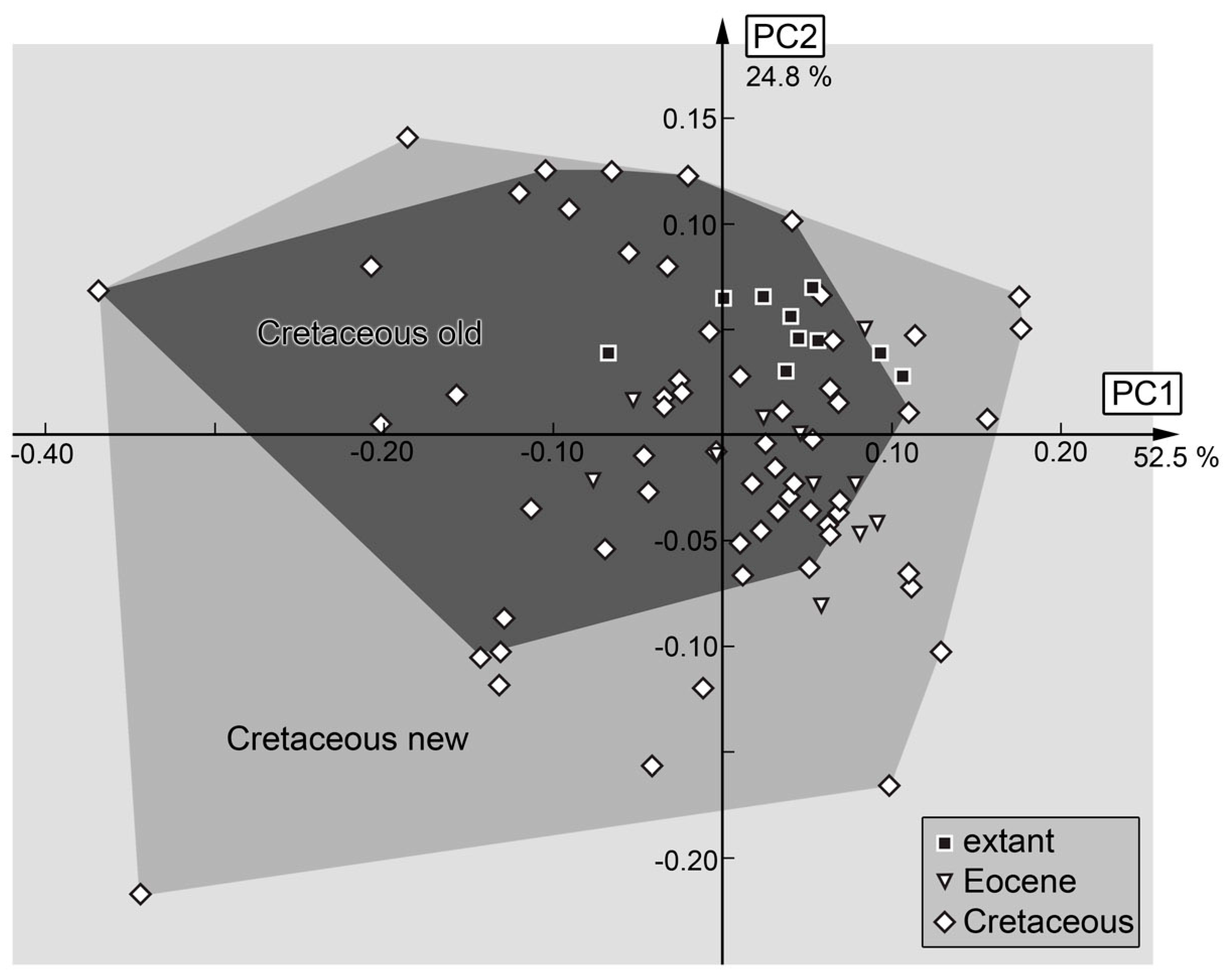
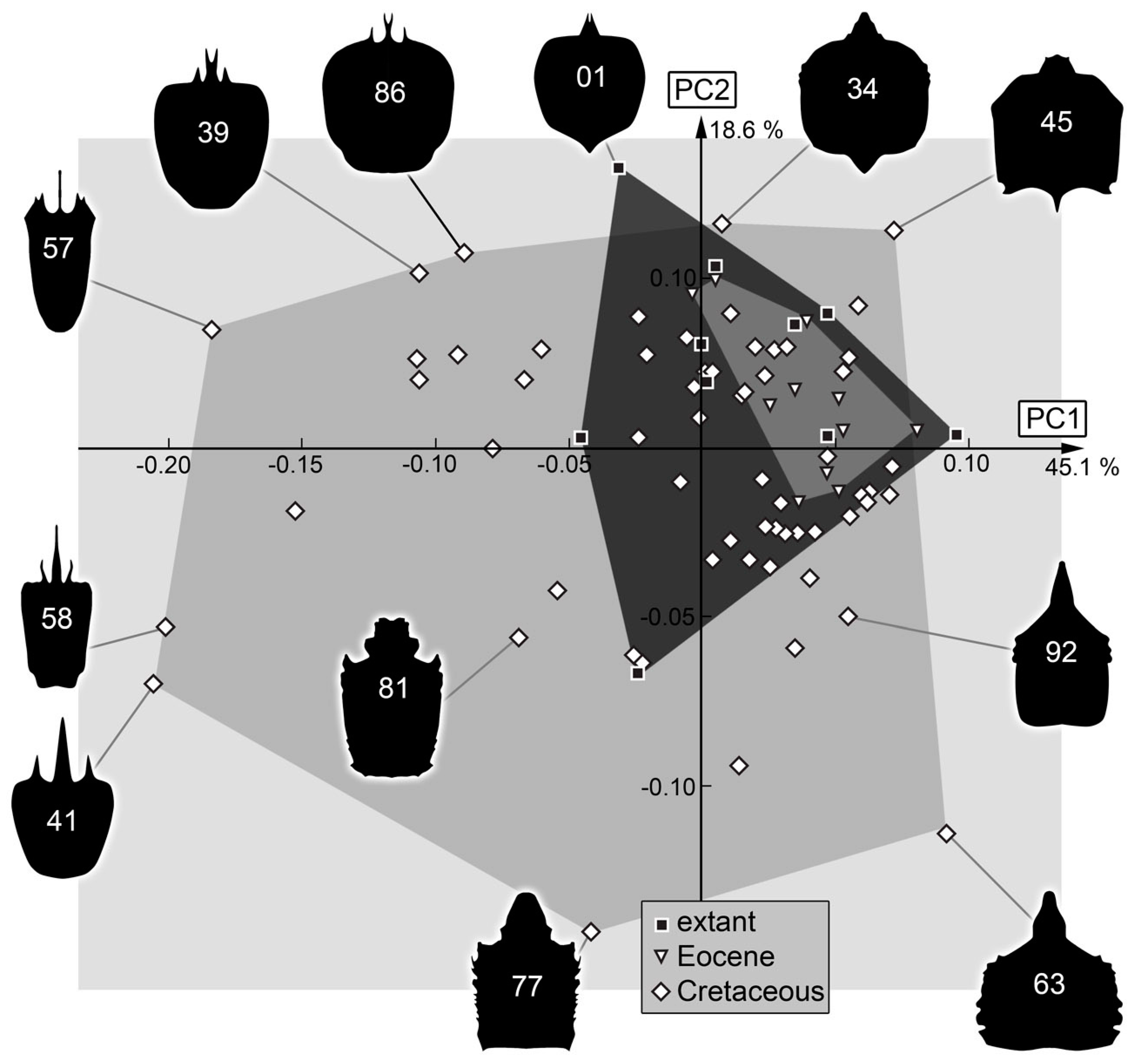
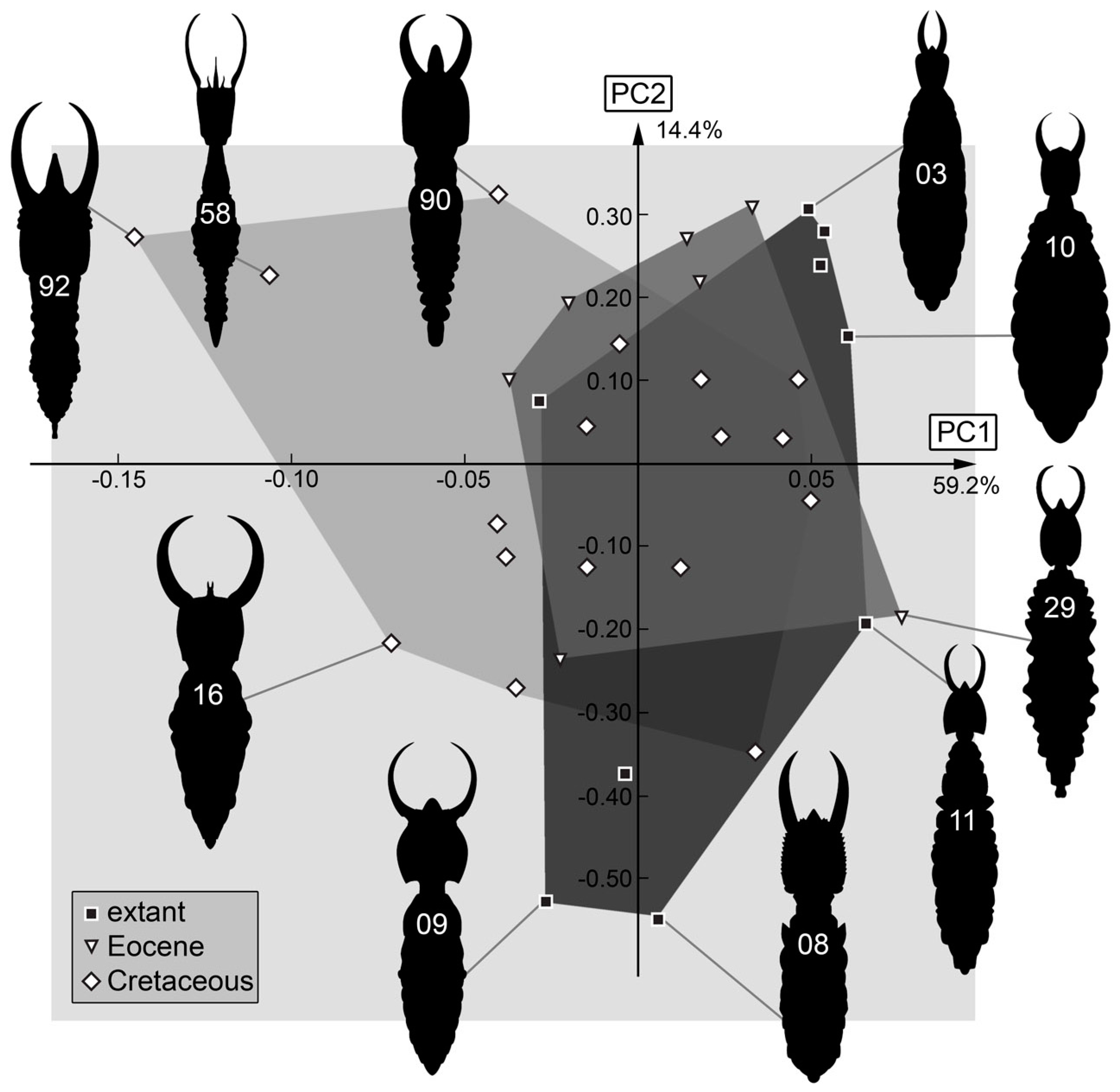
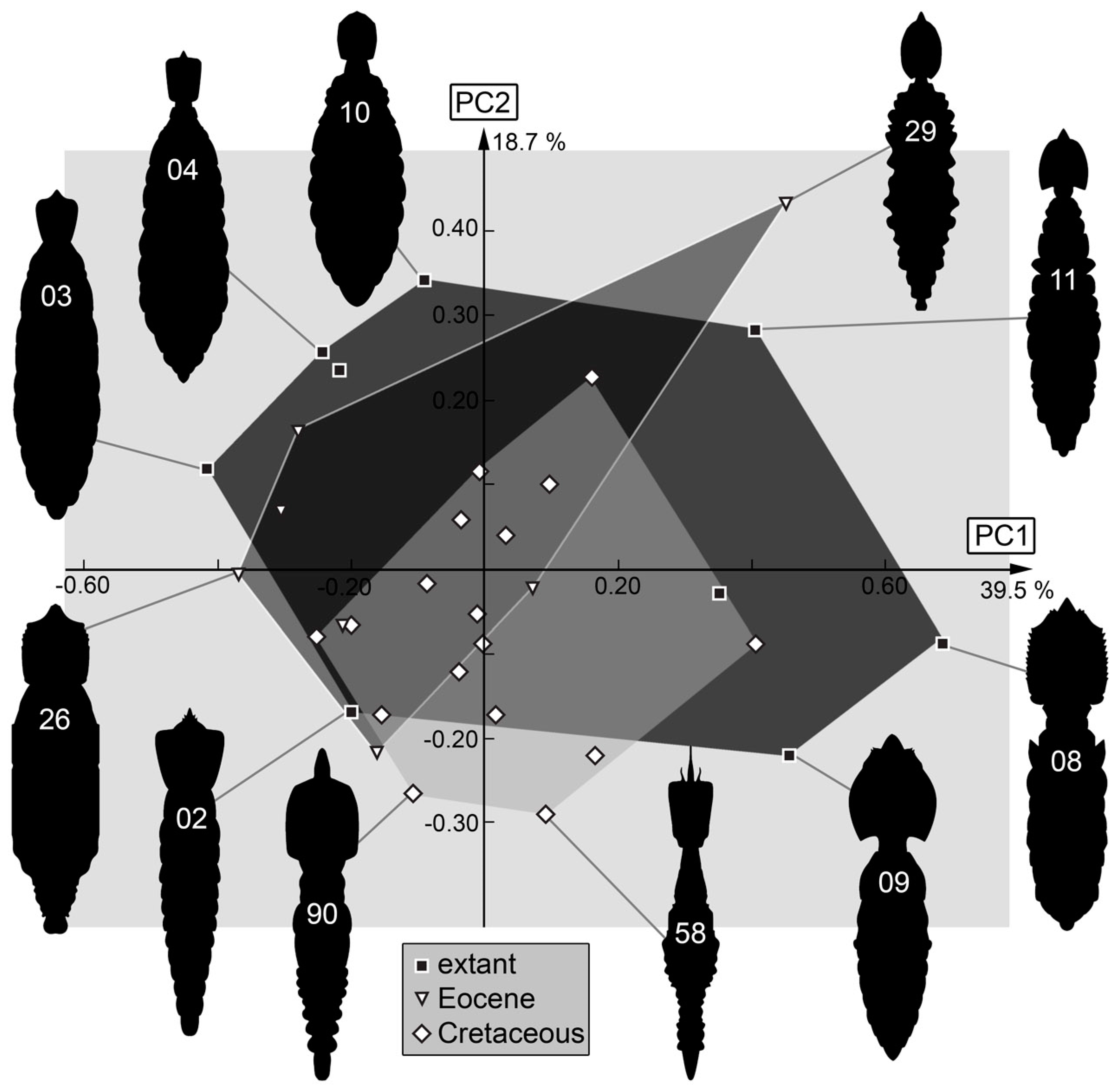
3.2. Shape Analysis
4. Discussion
4.1. General Observation: Loss of Diversity
4.2. Head and Stylet Shape
4.3. Stylet Shape
4.4. Head Capsule Shape
4.5. Body Shape including Stylets
4.6. Body Shape without Stylets
4.7. The Cretaceous Fauna and Its Peculiarities
4.8. Growth and Morphotypes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Amendt, J. Insect decline—A forensic issue? Insects 2021, 12, 324. [Google Scholar] [CrossRef] [PubMed]
- Dunn, R.R. Modern insect extinctions, the neglected majority. Conserv. Biol. 2005, 19, 1030–1036. [Google Scholar]
- Goulson, D. The insect apocalypse, and why it matters. Curr. Biol. 2019, 29, R967–R971. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [PubMed]
- Hallmann, C.A.; Ssymank, A.; Sorg, M.; de Kroon, H.; Jongejans, E. Insect biomass decline scaled to species diversity: General patterns derived from a hoverfly community. Proc. Natl. Acad. Sci. USA 2021, 118, e2002554117. [Google Scholar] [CrossRef]
- Kehoe, R.; Frago, E.; Sanders, D. Cascading extinctions as a hidden driver of insect decline. Ecol. Entomol. 2021, 46, 743–756. [Google Scholar] [CrossRef]
- Leather, S.R. “Ecological Armageddon”-more evidence for the drastic decline in insect numbers. Ann. Appl. Biol. 2017, 172, 1–3. [Google Scholar] [CrossRef]
- Peña, V.; Harvey, B.P.; Agostini, S.; Porzio, L.; Milazzo, M.; Horta, P.; Le Gall, L.; Hall-Spencer, J.M. Major loss of coralline algal diversity in response to ocean acidification. Glob. Change Biol. 2021, 27, 4785–4798. [Google Scholar] [CrossRef]
- Van der Sluijs, J.P. Insect decline, an emerging global environmental risk. Curr. Opin. Environ. Sust. 2020, 46, 39–42. [Google Scholar] [CrossRef]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef]
- Dangles, O.; Casas, J. Ecosystem services provided by insects for achieving sustainable development goals. Ecosys. Serv. 2019, 35, 109–115. [Google Scholar] [CrossRef]
- Elizalde, L.; Arbetman, M.; Arnan, X.; Eggleton, P.; Leal, I.R.; Lescano, M.N.; Saez, A.; Werenkraut, V.; Pirk, G.I. The ecosystem services provided by social insects: Traits, management tools and knowledge gaps. Biol. Rev. 2020, 95, 1418–1441. [Google Scholar] [CrossRef] [PubMed]
- Noriega, J.A.; Hortal, J.; Azcárate, F.M.; Berg, M.P.; Bonada, N.; Briones, M.J.; Del Toro, I.; Goulson, D.; Ibanez, S.; Landis, D.A.; et al. Research trends in ecosystem services provided by insects. Basic Appl. Ecol. 2018, 26, 8–23. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.V.; Garibaldi, L.A.; Hill, R.; Settele, J.; et al. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Ramos, D.L.; Cunha, W.L.; Evangelista, J.; Lira, L.A.; Rocha, M.V.C.; Gomes, P.A.; Frizzas, M.R.; Togni, P.H.B. Ecosystem services provided by insects in Brazil: What do we really know? Neotrop. Entomol. 2020, 49, 783–794. [Google Scholar] [CrossRef]
- Schowalter, T.D.; Noriega, J.A.; Tscharntke, T. Insect effects on ecosystem services—Introduction. Basic Appl. Ecol. 2018, 26, 1–7. [Google Scholar] [CrossRef]
- Haug, C.; Posada Zuluaga, V.; Zippel, A.; Braig, F.; Müller, P.; Gröhn, C.; Weiterschan, T.; Wunderlich, J.; Haug, G.T.; Haug, J.T. The morphological diversity of antlion larvae and their closest relatives over 100 million years. Insects 2022, 13, 587. [Google Scholar] [CrossRef]
- Haug, G.T.; Haug, C.; Pazinato, P.G.; Braig, F.; Perrichot, V.; Gröhn, C.; Müller, P.; Haug, J.T. The decline of silky lacewings and morphological diversity of long-nosed antlion larvae through time. Palaeont. Electron. 2020, 23, a39. [Google Scholar] [CrossRef]
- Haug, G.T.; Baranov, V.; Wizen, G.; Pazinato, P.G.; Müller, P.; Haug, C.; Haug, J.T. The morphological diversity of long-necked lacewing larvae (Neuroptera: Myrmeleontiformia). Bull. Geosci. 2021, 96, 431–457. [Google Scholar] [CrossRef]
- Haug, G.T.; Haug, C.; Haug, J.T. The morphological diversity of spoon-winged lacewing larvae and the first possible fossils from 99 million-year-old Kachin amber, Myanmar. Palaeodiversity 2021, 14, 133–152. [Google Scholar] [CrossRef]
- Haug, G.T.; Haug, C.; van der Wal, S.; Müller, P.; Haug, J.T. Split-footed lacewings declined over time: Indications from the morphological diversity of their antlion-like larvae. PalZ 2022, 96, 29–50. [Google Scholar] [CrossRef]
- Haug, J.T.; Baranov, V.; Schädel, M.; Müller, P.; Gröhn, C.; Haug, C. Challenges for understanding lacewings: How to deal with the incomplete data from extant and fossil larvae of Nevrorthidae? (Neuroptera). Fragm. Entomol. 2020, 52, 137–167. [Google Scholar] [CrossRef]
- Haug, J.T.; Pazinato, P.G.; Haug, G.T.; Haug, C. Yet another unusual new type of lacewing larva preserved in 100-million-year old amber from Myanmar. Riv. Ital. Paleontol. Stratigr. 2020, 126, 821–832. [Google Scholar] [CrossRef]
- Haug, J.T.; Haug, G.T.; Zippel, A.; van der Wal, S.; Müller, P.; Gröhn, C.; Wunderlich, J.; Hoffeins, C.; Hoffeins, H.-W.; Haug, C. Changes in the morphological diversity of larvae of lance lacewings, mantis lacewings and their closer relatives over 100 million years. Insects 2021, 12, 860. [Google Scholar] [CrossRef]
- Haug, J.T.; Linhart, S.; Haug, G.T.; Gröhn, C.; Hoffeins, C.; Hoffeins, H.-W.; Müller, P.; Weiterschan, T.; Wunderlich, J.; Haug, C. The diversity of aphidlion-like larvae over the last 130 million years. Insects 2022, 13, 336. [Google Scholar] [CrossRef]
- Haug, J.T.; van der Wal, S.; Gröhn, C.; Hoffeins, C.; Hoffeins, H.-W.; Haug, C. Diversity and fossil record of larvae of three groups of lacewings with unusual ecology and functional morphology: Ithonidae, Coniopterygidae and Sisyridae. Palaeont. Electron. 2022, 25, a14. [Google Scholar] [CrossRef]
- Zippel, A.; Kiesmüller, C.; Haug, G.T.; Müller, P.; Weiterschan, T.; Haug, C.; Hörnig, M.K.; Haug, J.T. Long-headed predators in Cretaceous amber—Fossil findings of an unusual type of lacewing larva. Palaeoentomology 2021, 4, 475–498. [Google Scholar] [CrossRef]
- Aspöck, U.; Aspöck, H. Kamelhälse, Schlammfliegen, Ameisenlöwen. Wer sind sie? (Insecta: Neuropterida: Raphidioptera, Megaloptera, Neuroptera). Stapfia 1999, 60, 1–34. [Google Scholar]
- Aspöck, U.; Plant, J.D.; Nemeschkal, H.L. Cladistic analysis of Neuroptera and their systematic position within Neuropterida (Insecta: Holometabola: Neuropterida: Neuroptera). Syst. Entomol. 2001, 26, 73–86. [Google Scholar] [CrossRef]
- Hassanpour, M.; Asadi, M.; Jooyandeh, A.; Madadi, H. Lacewings: Research and applied aspects. In Biological Control of Insect and Mite Pests in Iran; Progress in Biological Control; Karimi, J., Madadi, H., Eds.; Springer: Cham, Switzerland, 2021; pp. 175–194. [Google Scholar] [CrossRef]
- New, T.R. The lacewings (Insecta, Neuroptera) of Tasmania. Pap. Proc. R. Soc. Tasman. 1992, 126, 29–45. [Google Scholar] [CrossRef]
- Ardila-Camacho, A.; Martins, C.C.; Aspöck, U.; Contreras-Ramos, A. Comparative morphology of extant raptorial Mantispoidea (Neuroptera: Mantispidae, Rhachiberothidae) suggests a non-monophyletic Mantispidae and a single origin of the raptorial condition within the superfamily. Zootaxa 2021, 4992, 1–89. [Google Scholar] [CrossRef] [PubMed]
- Aspöck, U.; Aspöck, H.; Johnson, J.B.; Donga, T.K.; Duelli, P. Rhachiella malawica gen. nov., spec. nov. from Malawi-another beauty of the Afrotropics (Neuroptera: Rhachiberothidae). Zootaxa 2020, 4808, 131–140. [Google Scholar] [CrossRef]
- Haring, E.; Aspöck, U. Phylogeny of the Neuropterida: A first molecular approach. Syst. Entomol. 2004, 29, 415–430. [Google Scholar] [CrossRef]
- Jandausch, K.; Pohl, H.; Aspöck, U.; Winterton, S.L.; Beutel, R.G. Morphology of the primary larva of Mantispa aphavexelte Aspöck & Aspöck, 1994 (Neuroptera: Mantispidae) and phylogenetic implications to the order of Neuroptera. Arthropod Syst. Phyl. 2018, 76, 529–560. [Google Scholar] [CrossRef]
- Jandausch, K.; Beutel, R.G.; Bellstedt, R. The larval morphology of the spongefly Sisyra nigra (Retzius, 1783) (Neuroptera: Sisyridae). J. Morph. 2019, 280, 1742–1758. [Google Scholar] [CrossRef]
- Liu, X.; Winterton, S.L.; Wu, C.; Piper, R.; Ohl, M. A new genus of mantidflies discovered in the Oriental region, with a higher-level phylogeny of Mantispidae (Neuroptera) using DNA sequences and morphology. Syst. Entomol. 2015, 40, 183–206. [Google Scholar] [CrossRef]
- Nakamine, H.; Yamamoto, S. A new genus and species of thorny lacewing from Upper Cretaceous Kuji amber, northeastern Japan (Neuroptera, Rhachiberothidae). ZooKeys 2018, 802, 109–120. [Google Scholar] [CrossRef]
- Nakamine, H.; Yamamoto, S.; Takahashi, Y. Hidden diversity of small predators: New thorny lacewings from mid-Cretaceous amber from northern Myanmar (Neuroptera: Rhachiberothidae: Paraberothinae). Geological Mag. 2020, 157, 1149–1175. [Google Scholar] [CrossRef]
- Vasilikopoulos, A.; Misof, B.; Meusemann, K.; Lieberz, D.; Flouri, T.; Beutel, R.G.; Niehuis, O.; Wappler, T.; Rust, J.; Peters, R.S.; et al. An integrative phylogenomic approach to elucidate the evolutionary history and divergence times of Neuropterida (Insecta: Holometabola). BMC Evol. Biol. 2020, 20, 64. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.; Garzón-Orduña, I.J.; Winterton, S.L.; Yan, Y.; Aspöck, U.; Aspöck, H.; Yang, D. Mitochondrial phylogenomics illuminates the evolutionary history of Neuropterida. Cladistics 2017, 33, 617–636. [Google Scholar] [CrossRef]
- Winterton, S.L.; Hardy, N.B.; Wiegmann, B.M. On wings of lace: Phylogeny and Bayesian divergence time estimates of Neuropterida (Insecta) based on morphological and molecular data. Syst. Entomol. 2010, 35, 349–378. [Google Scholar] [CrossRef]
- Winterton, S.L.; Gillung, J.P.; Garzón-Orduña, I.J.; Badano, D.; Breitkreuz, L.C.V.; Duelli, P.; Engel, M.S.; Liu, X.; Machado, R.J.P.; Mansell, M.; et al. Evolution of green lacewings (Neuroptera: Chrysopidae): An anchored phylogenomics approach. Syst. Entomol. 2019, 44, 514–526. [Google Scholar] [CrossRef]
- Aspock, H.; Aspock, U. Synopsis der Systematik, Okologie und Biogeographie der Neuropteren Mitteleuropas im Spiegel der Neuropteren-Fauna von Linz und Oberosterreich, sowie Bestimmungsschlussel fur die mitteleuropaischen Neuropteren und Beschreibung von Coniopteryx lentiae nov. spec. Naturkundl. Jahrb. Stadt Linz 1964, 1964, 127–282. [Google Scholar]
- Cover, M.R.; Bogan, M.T. Chapter 41: Minor insect orders. In Thorp and Covich’s Freshwater Invertebrates; Thorp, J., Rogers, D.C., Eds.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 1059–1072. [Google Scholar]
- Gepp, J. Erforschungsstand der Neuropteren. Larven der Erde (mit einem Schlüssel zur Larvaldiagnose der Familien, einer Übersicht von 340 beschriebenen Larven und 600 Literaturzitaten). In Progress in Worlds Neuropterology. In Proceedings of the First International Symposium on Neuropterology, Graz, Austria, 22–26 September 1980; pp. 183–239. [Google Scholar]
- MacLeod, E.G. A Comparative Morphological Study of the Head Capsule and Cervix of Larval Neuroptera (Insecta). Ph.D. Thesis, Harvard University, Cambridge, MA, USA, 1964. [Google Scholar]
- Riek, E.F. Neuroptera (lacewings). In The Insects of Australia; Melbourne University Press: Melbourne, VIC, Australia, 1970; pp. 472–494. [Google Scholar]
- Tauber, C.A.; Tauber, M.J.; Albuquerque, G.S. Neuroptera (lacewings, antlions). In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Cambridge, MA, USA, 2003; pp. 695–707. [Google Scholar]
- Zimmermann, D.; Randolf, S.; Aspöck, U. From chewing to sucking via phylogeny–From sucking to chewing via ontogeny: Mouthparts of Neuroptera. In Insect Mouthparts, Zoological Monographs 5; Krenn, H.W., Ed.; Springer: Berlin, Germany, 2019; pp. 361–385. [Google Scholar]
- Rojht, H.; Budija, F.; Trdan, S. Effect of temperature on cannibalism rate between green lacewings larvae (Chrysoperla carnea [Stephens], Neuroptera, Chrysopidae). Acta Agricult. Sloven. 2009, 93, 5–9. [Google Scholar] [CrossRef]
- Tauber, C.A.; Tauber, M.J.; Albuquerque, G.S. Debris-carrying in larval Chrysopidae: Unraveling its evolutionary history. Ann. Entomol. Soc. Am. 2014, 107, 295–314. [Google Scholar] [CrossRef]
- Tsukaguchi, S. Descriptions of the larvae of Chrysopa Leach (Neuroptera, Chrysopidae) of Japan. Kontyû 1978, 46, 99–122. [Google Scholar]
- Tsukaguchi, S. Taxonomic notes on Brinckochrysa kintoki (Okamoto) (Neuroptera: Chrysopidae). Kontyû 1979, 47, 358–366. [Google Scholar]
- Zhao, C.; Ang, Y.; Wang, M.; Gao, C.; Zhang, K.; Tang, C.; Liu, X.; Li, M.; Yang, D.; Meier, R. Contribution to understanding the evolution of holometaboly: Transformation of internal head structures during the metamorphosis in the green lacewing Chrysopa pallens (Neuroptera: Chrysopidae). BMC Evol. Biol. 2020, 20, 79. [Google Scholar] [CrossRef]
- Krakauer, A.H.; Tauber, C.A. Larvae of Micromus: Generic characteristics and a description of Micromus subanticus (Neuroptera: Hemerobiidae). Ann. Entomol. Soc. Am. 1996, 89, 203–211. [Google Scholar] [CrossRef]
- MacLeod, E.G. The immature stages of Boriomyia fidelis (Banks) with taxonomic notes on the affinities of the genus Boriomyia (Neuroptera: Hemerobiidae). Psyche 1960, 67, 26–40. [Google Scholar] [CrossRef]
- Miller, G.L.; Lambdin, P.L. Redescriptions of the larval stages of Hemerobius stigma Stephens (Neuroptera: Hemerobiidae). Florida Entomol. 1984, 67, 377–382. [Google Scholar] [CrossRef]
- Nakahara, W. Early stages of some Japanese Hemerobiidae including two new species. Kontyû 1954, 21, 41–46. [Google Scholar]
- Tauber, C.A.; Krakauer, A.H. Larval characteristics and generic placement of endemic Hawaiian hemerobiids (Neuroptera). Pacif. Sci. 1997, 51, 413–423. [Google Scholar]
- Ardila-Camacho, A.; Machado, R.J.P.; Contreras-Ramos, A. A review of the biology of Symphrasinae (Neuroptera: Rhachiberothidae), with the description of the egg and primary larva of Plega Navás, 1928. Zool. Anzeiger. 2021, 294, 165–185. [Google Scholar] [CrossRef]
- Parker, F.D.; Stange, L.A. Systematic and biological notes on the tribe Platymantispini (Neuroptera: Mantispidae) and the description of a new species of Plega from Mexico. Canad. Entomol. 1965, 97, 604–612. [Google Scholar] [CrossRef]
- Imms, A.D. A General Textbook of Entomology; Methuen & Co.: London, UK, 1930; p. 703. [Google Scholar]
- Pierre, F. Morphologie, milieu biologique et comportement de trois Crocini nouveaux du Sahara nord-occidental (Planipennes, Nemopteridae). Ann. Soc. Entomol. Fr. 1952, 119, 1–22. [Google Scholar]
- Schaum, H.R. Necrophilus arenarius Roux, die muthmassliche Larve von Nemoptera. Berl. Entomol. Z. 1857, 1, 1–9. [Google Scholar] [CrossRef]
- Wheeler, W.M. Is Necrophylus arenarius Roux the larva of Pterocroce storeyi Withycombe. Psyche 1929, 36, 313–320. [Google Scholar] [CrossRef]
- Withycombe, C.L. XII. Systematic notes on the Crocini (Nemopteridae), with descriptions of new genera and species. Trans. R. Entomol. Soc. Lond. 1923, 71, 269–287. [Google Scholar] [CrossRef]
- Badano, D.; Pantaleoni, R.A. The larvae of European Ascalaphidae (Neuroptera). Zootaxa 2014, 3796, 287–319. [Google Scholar] [CrossRef]
- New, T.R.; Lambkin, K.J. The larva of Norfolius (Neuroptera: Nymphidae). Syst. Entomol. 1989, 14, 93–98. [Google Scholar] [CrossRef]
- Weele, V.D.H.W. Ascalaphiden. Collections zoologiques du baron Edm. de Selys Longchamps. Cat. Syst. Descr. 1908, 8, 1–326. [Google Scholar]
- Pérez-de la Fuente, R.; Engel, M.S.; Delclòs, X.; Peñalver, E. Straight-jawed lacewing larvae (Neuroptera) from Lower Cretaceous Spanish amber, with an account on the known amber diversity of neuropterid immatures. Cretac. Res. 2020, 106, 104200. [Google Scholar] [CrossRef]
- Kimmins, D.E. Keys to the British species of aquatic Megaloptera and Neuroptera. Freshw. Biol. Assoc. Sci. Publ. 1962, 8, 1–23. [Google Scholar]
- Notteghem, P. La Sisyre noire (Sisyra nigra), Névroptère autochtone, parasite de la Pectinatelle (Pectinatella magnifica), Bryozoaire allochtone. Rev. Sci. Bourg. Nat. 2016, 23, 133–140. [Google Scholar]
- Parfin, S.I.; Gurney, A.B. The spongilla-flies, with special reference to those of the western hemisphere (Sisyridae, Neuroptera). Proc. U. S. Natl. Mus. 1956, 105, 421–529. [Google Scholar] [CrossRef]
- Weißmair, W. Präimaginale Stadien, Biologie und Ethologie der europäischen Sisyridae (Neuropterida: Neuroptera). Stapfia 1999, 60, 101–128. [Google Scholar]
- Gurney, A.B. Notes on Dilaridae and Berothidae, with special reference to the immature stages of the Nearctic genera (Neuroptera). Psyche 1947, 54, 145–169. [Google Scholar] [CrossRef]
- Minter, L.R. A comparison of the eggs and first-instar larvae of Mucroberotha vesicaria Tjeder with those of other species in the families Berothidae and Mantispidae (Insecta: Neuroptera). In Advances in Neuropterology, Proceedings of the Third International Symposium on Neuropterology, Berg en Dal, Kruger National Park, South Africa, 3–4 February 1988; Mansell, M.W., Aspöck, H., Eds.; South African Department of Agricultural Development: Pretoria, South Africa, 1990; pp. 115–129. [Google Scholar]
- Möller, A.; Minter, L.R.; Olivier, P.A.S. Larval morphology of Podallea vasseana Navás and Podallea manselli Aspöck & Aspöck from South Africa (Neuroptera: Berothidae). Afric. Entomol. 2006, 14, 1–12. [Google Scholar]
- Tillyard, R.J. Studies in Australian Neuroptera. No. iv. The families Ithonidae, Hemerobiidae, Sisyridae, Berothidae, and the new family Trichomatidae; with a discussion of their characters and relationships, and descriptions of new and little-known genera and species. Proc. Linn. Soc. N. S. W. 1916, 41, 269–332. [Google Scholar]
- Toschi, C.A. Observations on Lomamyia latipennis, with a description of the first instar larva. Pan-Pacif. Entomol. 1964, 40, 21–26. [Google Scholar]
- Dorey, J.B.; Merritt, D.J. First observations on the life cycle and mass eclosion events in a mantis fly (Family Mantispidae) in the subfamily Drepanicinae. Biodiv. Data J. 2017, 5, e21206. [Google Scholar] [CrossRef]
- Hoffman, K.M.; Brushwein, J.R. Descriptions of the larvae and pupae of some North American Mantispinae (Neuroptera: Mantispidae) and development of a system of larval chaetotaxy for Neuroptera. Trans. Amer. Entomol. Soc. 1992, 118, 159–196. [Google Scholar]
- Lucchese, E. Ricerche sulla Mantispa perla Pallas (Neuroptera Planipennia—Fam. Mantispidae). [I.] Nota preventiva su nuovi reperti concernenti l’etologia della larva della 1a età. Ann. Fac. Agr. R. Univ. Stud. Perugia 1955, 11, 242–262. [Google Scholar]
- Gepp, J. Der Bachhaft Osmylus fulvicephalus-240 Jahre nach seiner Beschreibung durch Johannes Antonius Scopoli—Österreichs Insekt des Jahres (Osmylidae, Neruoptera). Carinthia II Mitt. Naturhist. Landesmus. Kärnten 2003, 193, 325–334. [Google Scholar]
- Lestage, J.A. La ponte et la larvule de l’Osmylus chrysops L. (Planipenne). Ann. Biol. Lacust. 1920, 10, 226–230. [Google Scholar]
- Matsuno, S.; Yoshitomi, H. Descriptions of three larvae of Osmylus species from Japan (Neuroptera: Osmylidae), with a proposed naming system for the larval sclerites. Zootaxa 2016, 4189, 348–366. [Google Scholar] [CrossRef]
- Stitz, H. Planipennia. In Biologie der Tiere Deutschlands. Lfg. 33, Teil 35; Schultze, P., Ed.; Borntraeger: Berlin, Germany, 1931; pp. 67–304. [Google Scholar]
- Wundt, H. Der Kopf der Larve von Osmylus chrysops L. (Neuroptera, Planipennia). Zool. Jahrb. Abt. Anat. Ontog. 1961, 79, 557–662. [Google Scholar]
- Lu, X.; Zhang, W.; Liu, X. New long-proboscid lacewings of the mid-Cretaceous provide insights into ancient plant-pollinator interactions. Sci. Rep. 2016, 6, 25382. [Google Scholar] [CrossRef] [Green Version]
- Makarkin, V.N. Re-description of Grammapsychops lebedevi Martynova, 1954 (Neuroptera: Psychopsidae) with notes on the Late Cretaceous psychopsoids. Zootaxa 2018, 4524, 581–594. [Google Scholar] [CrossRef]
- Bakkes, D.K.; Mansell, M.W.; Sole, C.L. Phylogeny and historical biogeography of silky lacewings (Neuroptera: Psychopsidae). Syst. Entomol. 2018, 43, 43–55. [Google Scholar] [CrossRef]
- Oswald, J.D. Revision of the African silky lacewing genus Zygophlebius Navas (Neuroptera: Psychopsidae). Afric. Entomol. 1994, 2, 83–96. [Google Scholar] [CrossRef]
- Peng, Y.; Makarkin, V.N.; Ren, D. Diverse new Middle Jurassic Osmylopsychopidae (Neuroptera) from China shed light on the classification of psychopsoids. J. Syst. Palaeont. 2016, 14, 261–295. [Google Scholar] [CrossRef]
- Badano, D.; Aspöck, U.; Aspöck, H.; Cerretti, P. Phylogeny of Myrmeleontiformia based on larval morphology (Neuropterida: Neuroptera). Syst. Entomol. 2017, 42, 94–117. [Google Scholar] [CrossRef]
- Winterton, S.L.; Lemmon, A.R.; Gillung, J.P.; Garzon, I.J.; Badano, D.; Bakkes, D.K.; Breitkreuz, L.C.V.; Engel, M.S.; Lemmon, E.M.; Liu, X. Evolution of lacewings and allied orders using anchored phylogenomics (Neuroptera, Megaloptera, Raphidioptera). Syst. Entomol. 2018, 43, 330–354. [Google Scholar] [CrossRef]
- Garzón-Orduña, I.J.; Menchaca-Armenta, I.; Contreras-Ramos, A.; Liu, X.; Winterton, S.L. The phylogeny of brown lacewings (Neuroptera: Hemerobiidae) reveals multiple reductions in wing venation. BMC Evol. Biol. 2016, 16, 192. [Google Scholar] [CrossRef]
- Engel, M.S.; Grimaldi, D.A. Diverse Neuropterida in Cretaceous amber, with particular reference to the paleofauna of Myanmar (Insecta). Nova Suppl. Entomol. 2008, 20, 1–86. [Google Scholar]
- Perrichot, V. Environnements Paraliques à Ambre et à Végétaux du Crétacé Nord-aquitain (Charentes, Sud-Ouest de la France). Ph.D. Thesis, Université Rennes 1, Rennes, France, 2003. [Google Scholar]
- MacLeod, E.G. The Neuroptera of the Baltic Amber. I. Ascalaphidae, Nymphidae, and Psychopsidae. Psyche 1970, 77, 045459. [Google Scholar] [CrossRef]
- Engel, M.S.; Winterton, S.L.; Breitkreuz, L.C.V. Phylogeny and evolution of Neuropterida: Where have wings of lace taken us? Ann. Rev. Entomol. 2018, 63, 531–551. [Google Scholar] [CrossRef]
- Gröhn, C. Einschlüsse im Baltischen Bernstein; Verlag Wachholtz: Kiel, Germany, 2015; p. 424. [Google Scholar]
- Scheven, J. Bernstein-Einschlüsse: Eine untergegangene Welt bezeugt die Schöpfung. Erinnerungen an die Welt vor der Sintflut; Kuratorium Lebendige Vorwelt e.V.: Hofheim, Germany, 2004. [Google Scholar]
- Weitschat, W.; Wichard, W. Atlas der Pflanzen und Tiere im Baltischen Bernstein; Friedrich Pfeil: Munich, Germany, 1998. [Google Scholar]
- Ross, A. Amber: The Natural Time Capsule; Firefly Books: Richmond Hill, ON, USA, 2009; p. 112. [Google Scholar]
- Cruickshank, R.D.; Ko, K. Geology of an amber locality in the Hukawng Valley, northern Myanmar. J. Asian Earth Sci. 2003, 21, 441–455. [Google Scholar] [CrossRef]
- Grimaldi, D.A.; Engel, M.S.; Nascimbene, P.C. Fossiliferous Cretaceous amber from Myanmar (Burma): Its rediscovery, biotic diversity, and paleontological significance. Amer. Mus. Nov. 2002, 3361, 1–71. [Google Scholar] [CrossRef]
- Shi, G.; Grimaldi, D.A.; Harlow, G.E.; Wang, J.; Wang, J.; Yang, M.; Lei, W.; Li, Q.; Li, X. Age constraint on Burmese amber based on U–Pb dating of zircons. Cretac. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Yu, T.; Kelly, R.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.; Zhang, H.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B. Fiji: An open-source platform for biological-image analysis. Nat. Meth. 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Iwata, H.; Ukai, Y. SHAPE: A computer program package for quantitative evaluation of biological shapes based on elliptic Fourier descriptors. J. Hered. 2002, 93, 384–385. [Google Scholar] [CrossRef]
- Braig, F.; Haug, J.T.; Schädel, M.; Haug, C. A new thylacocephalan crustacean from the Upper Jurassic lithographic limestones of southern Germany and the diversity of Thylacocephala. Palaeodiversity 2019, 12, 69–87. [Google Scholar] [CrossRef]
- Haug, C.; Zippel, A.; Hassenbach, C.; Haug, G.T.; Haug, J.T. A split-footed lacewing larva from about 100-million-year-old amber indicates a now extinct hunting strategy for neuropterans. Bull. Geosci. 2022, 97, 453–464. [Google Scholar] [CrossRef]
- Badano, D.; Engel, M.S.; Basso, A.; Wang, B.; Cerretti, P. Diverse Cretaceous larvae reveal the evolutionary and behavioural history of antlions and lacewings. Nat. Comm. 2018, 9, 3257. [Google Scholar] [CrossRef]
- Haug, C.; Herrera-Flórez, A.F.; Müller, P.; Haug, J.T. Cretaceous chimera—An unusual 100-million-year old neuropteran larva from the “experimental phase” of insect evolution. Palaeodiversity 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Zippel, A.; Baranov, V.A.; Hammel, J.U.; Hörnig, M.K.; Haug, C.; Haug, J.T. The first fossil immature of Elmidae: An unusual riffle beetle larva preserved in Baltic amber. PeerJ 2022, 10, e13025. [Google Scholar] [CrossRef]
- Haug, J.T.; Haug, C. 100 million-year-old straight-jawed lacewing larvae with enormously inflated trunks represent the oldest cases of extreme physogastry in insects. Sci. Rep. 2022, 12, 12760. [Google Scholar] [CrossRef] [PubMed]
- Haug, J.T.; Müller, P.; Haug, C. A 100-million-year old predator: A fossil neuropteran larva with unusually elongated mouthparts. Zool. Lett. 2019, 5, 29. [Google Scholar] [CrossRef]
- Haug, J.T.; Müller, P.; Haug, C. A 100-million-year old slim insectan predator with massive venom-injecting stylets—A new type of neuropteran larva from Burmese amber. Bull. Geosci. 2019, 94, 431–440. [Google Scholar] [CrossRef]
- Haug, J.T.; Baranov, V.; Müller, P.; Haug, C. New extreme morphologies as exemplified by 100 million-year-old lacewing larvae. Sci. Rep. 2021, 11, 20432. [Google Scholar] [CrossRef]
- Pérez-de la Fuente, R.; Delclòs, X.; Penalver, E.; Speranza, M.; Wierzchos, J.; Ascaso, C.; Engel, M.S. Early evolution and ecology of camouflage in insects. Proc. Natl. Acad. Sci. USA 2012, 109, 21414–21419. [Google Scholar] [CrossRef] [PubMed]
- Pérez-de la Fuente, R.; Delclòs, X.; Penalver, E.; Engel, M.S. A defensive behavior and plant-insect interaction in Early Cretaceous amber—The case of the immature lacewing Hallucinochrysa diogenesi. Arthropod Struct. Dev. 2016, 45, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Haug, J.T.; Müller, P.; Haug, C. The ride of the parasite: A 100-million-year old mantis lacewing larva captured while mounting its spider host. Zool. Lett. 2018, 4, 31. [Google Scholar] [CrossRef]
- Hörnig, M.K.; Haug, C.; Müller, P.; Haug, J.T. Not quite social–Possible cases of gregarious behaviour of immatures of various lineages of Insecta preserved in 100-million-year-old amber. Bull. Geosci. 2022, 97, 69–87. [Google Scholar] [CrossRef]
- Wang, B.; Xia, F.; Engel, M.S.; Perrichot, V.; Shi, G.; Zhang, H.; Chen, J.; Jarzembowski, E.A.; Wappler, T.; Rust, J. Debris-carrying camouflage among diverse lineages of Cretaceous insects. Sci. Adv. 2016, 2, e1501918. [Google Scholar] [CrossRef]
- Badano, D.; Fratini, M.; Maugeri, L.; Palermo, F.; Pieroni, N.; Cedola, A.; Haug, J.T.; Weiterschan, T.; Velten, J.; Mei, M.; et al. X-ray microtomography and phylogenomics provide insights into the morphology and evolution of an enigmatic Mesozoic insect larva. Syst. Entomol. 2021, 46, 672–684. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, W.; Winterton, S.L.; Breitkreuz, L.C.; Engel, M.S. Early morphological specialization for insect-spider associations in Mesozoic lacewings. Curr. Biol. 2016, 26, 1590–1594. [Google Scholar] [CrossRef]
- Liu, X.; Shi, G.; Xia, F.; Lu, X.; Wang, B.; Engel, M.S. Liverwort mimesis in a Cretaceous lacewing larva. Curr. Biol. 2018, 28, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.; Liu, H.; Jarzembowski, E.A. High morphological disparity of neuropteran larvae during the Cretaceous revealed by a new large species. Geol. Mag. 2022, 159, 954–962. [Google Scholar] [CrossRef]
- Hayashi, M.; Ohba, S.Y. Mouth morphology of the diving beetle Hyphydrus japonicus (Dytiscidae: Hydroporinae) is specialized for predation on seed shrimps. Biol. J. Linn. Soc. 2018, 125, 315–320. [Google Scholar] [CrossRef]
- Baranov, V.; Haug, C.; Fowler, M.; Kaulfuss, U.; Müller, P.; Haug, J.T. Summary of the fossil record of megalopteran and megalopteran-like larvae, with a report of new specimens. Bull. Geosci. 2022, 97, 89–108. [Google Scholar] [CrossRef]
- Batelka, J.; Prokop, J.; Pohl, H.; Bai, M.; Zhang, W.; Beutel, R.G. Highly specialized Cretaceous beetle parasitoids (Ripiphoridae) identified with optimized visualization of microstructures. Syst. Entomol. 2019, 44, 396–407. [Google Scholar] [CrossRef]
- Batelka, J.; Engel, M.S.; Prokop, J. The complete life cycle of a Cretaceous beetle parasitoid. Curr. Biol. 2021, 31, R118–R119. [Google Scholar] [CrossRef]
- Beutel, R.G.; Zhang, W.W.; Pohl, H.; Wappler, T.; Bai, M. A miniaturized beetle larva in Cretaceous Burmese amber: Reinterpretation of a fossil “strepsipteran triungulin”. Insect Syst. Evol. 2016, 47, 83–91. [Google Scholar] [CrossRef]
- Haug, C.; Haug, G.T.; Zippel, A.; van der Wal, S.; Haug, J.T. The earliest record of fossil solid-wood-borer larvae—Immature beetles in 99 million-year-old Myanmar amber. Palaeoentomology 2021, 4, 390–404. [Google Scholar] [CrossRef]
- Kirejtshuk, A.G.; Azar, D. New taxa of beetles (Insecta, Coleoptera) from Lebanese amber with evolutionary and systematic comments. Alavesia 2008, 2, e46. [Google Scholar]
- Liu, H.; Beutel, R.G.; Makarov, K.V.; Jarzembowski, E.A.; Xiao, C.; Luo, C. The first larval record of Migadopinae (Coleoptera: Adephaga: Carabidae) from mid-Cretaceous Kachin amber, northern Myanmar. Cretac. Res. 2023, 142, 105413. [Google Scholar]
- Poinar, G. Discoscapidae fam. nov. (Hymenoptera: Apoidea), a new family of stem lineage bees with associated beetle triungulins in mid-Cretaceous Burmese amber. Palaeodiversity 2020, 13, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Poinar, G., Jr.; Brown, A. New genera and species of jumping ground bugs (Hemiptera: Schizopteridae) in Dominican and Burmese amber, with a description of a meloid (Coleoptera: Meloidae) triungulin on a Burmese specimen. Ann. Soc. Entomol. France 2014, 50, 372–381. [Google Scholar] [CrossRef]
- Poinar, G., Jr.; Brown, A. Dating the co-evolution between bees and beetle triungulins (Coleoptera: Cleridae) to the mid-Cretaceous. Biosis: Biol. Syst. 2020, 1, 2–7. [Google Scholar] [CrossRef]
- Poinar, G., Jr.; Poinar, R. Ancient hastisetae of Cretaceous carrion beetles (Coleoptera: Dermestidae) in Myanmar amber. Arthropod Struct. Dev. 2016, 45, 642–645. [Google Scholar] [CrossRef]
- Xia, F.; Yang, G.; Zhang, Q.; Shi, G.; Wang, B. Amber: Life Through Time and Space; Science Press: Beijing, China, 2015; p. 197. [Google Scholar]
- Zhang, W.W. Frozen Dimensions. In The Fossil Insects and Other Invertebrates in Amber; Chongqing University Press: Chongqing, China, 2017. [Google Scholar]
- Zhao, X.; Zhao, X.; Jarzembowski, E.A.; Wang, B. The first whirligig beetle larva from mid-Cretaceous Burmese amber (Coleoptera: Adephaga: Gyrinidae). Cretac. Res. 2019, 99, 41–45. [Google Scholar] [CrossRef]
- Zhao, X.; Zhao, X.; Jarzembowski, E.; Tian, Y.; Chen, L. The first record of brachypsectrid larva from mid-Cretaceous Burmese amber (Coleoptera: Polyphaga). Cretac. Res. 2020, 113, 104493. [Google Scholar] [CrossRef]
- Gustafson, G.T.; Michat, M.C.; Balke, M. Burmese amber reveals a new stem lineage of whirligig beetle (Coleoptera: Gyrinidae) based on the larval stage. Zool. J. Linn. Soc. 2020, 189, 1232–1248. [Google Scholar] [CrossRef]
- Baranov, V.A.; Wang, Y.; Gašparič, R.; Wedmann, S.; Haug, J.T. Eco-morphological diversity of larvae of soldier flies and their closest relatives in deep time. PeerJ 2020, 8, e10356. [Google Scholar] [CrossRef]
- Liu, Y.; Hakim, M.; Huang, D. First stratiomyomorphan larvae in the mid-Cretaceous amber from Myanmar (Diptera: Brachycera). Cretac. Res. 2020, 106, 104265. [Google Scholar] [CrossRef]
- Álvarez-Parra, S.; Pérez-de la Fuente, R.; Peñalver, E.; Barrón, E.; Alcalá, L.; Pérez-Cano, J.; Martín-Closas, C.; Trabelsi, K.; Meléndez, N.; López Del Valle, R.; et al. Dinosaur bonebed amber from an original swamp forest soil. eLife 2021, 10, e72477. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.C. In search for the unlikely: Leaf-mining caterpillars (Gracillariidae, Lepidoptera) from Upper Cretaceous and Eocene ambers. Zitteliana 2021, 95, 135–145. [Google Scholar] [CrossRef]
- Gauweiler, J.; Haug, C.; Müller, P.; Haug, J.T. Lepidopteran caterpillars in the Cretaceous: Were they a good food source for early birds? Palaeodiversity 2022, 15, 45–59. [Google Scholar] [CrossRef]
- Grimaldi, D.; Engel, M.S. Evolution of the Insects; Cambridge University Press: Cambridge, UK, 2005; p. 755. [Google Scholar]
- Haug, J.T.; Haug, C. A 100 million-year-old armoured caterpillar supports the early diversification of moths and butterflies. Gondw. Res. 2021, 93, 101–105. [Google Scholar] [CrossRef]
- MacKay, M.R. Lepidoptera in Cretaceous amber. Science 1970, 167, 379–380. [Google Scholar] [CrossRef]
- Engel, M.S. The smallest snakefly (Raphidioptera: Mesoraphidiidae): A new species in Cretaceous amber from Myanmar, with a catalog of fossil snakeflies. Am. Mus. Nov. 2002, 3363, 1–22. [Google Scholar] [CrossRef]
- Haug, J.T.; Müller, P.; Haug, C. A 100 million-year-old snake-fly larva with an unusually large antenna. Bull. Geosci. 2020, 95, 167–177. [Google Scholar] [CrossRef]
- Haug, J.T.; Engel, M.S.; Mendes dos Santos, P.; Haug, G.T.; Müller, P.; Haug, C. Declining morphological diversity in snakefly larvae during last 100 million years. PalZ 2022, 96, 749–780. [Google Scholar] [CrossRef]
- Aspöck, U. Phylogeny of the Neuropterida (Insecta: Holometabola). Zool. Scr. 2002, 31, 51–55. [Google Scholar] [CrossRef]
- Aspöck, U.; Aspöck, H. Phylogenetic relevance of the genital sclerites of Neuropterida (Insecta: Holometabola). Syst. Entomol. 2008, 33, 97–127. [Google Scholar] [CrossRef]
- Aspöck, U.; Haring, E.; Aspöck, H. The phylogeny of the Neuropterida: Long lasting and current controversies and challenges (Insecta: Endopterygota). Arthropod Syst. Phyl. 2012, 70, 119–129. [Google Scholar]
- Bakkes, D.K.; Sole, C.L.; Mansell, M.W. Revision of afrotropical silky lacewings (Neuroptera: Psychopsidae). Zootaxa 2017, 4362, 151–212. [Google Scholar] [CrossRef] [PubMed]
- Froggatt, W.W. Australian Insects; W. Brooks and Co.: Sydney, NSW, Australia, 1907; p. 449. [Google Scholar]
- New, T.R. Planipennia, Lacewings. Handbuch der Zoologie; Vol. 4 (Arthropoda: Insecta), Part 30; Walter de Gruyter: Berlin, Germany, 1989; p. 132. [Google Scholar]
- New, T.R. Neuroptera (lacewings). In The Insects of Australia, 2nd ed.; Naumann, I.D., Ed.; Melbourne University Press: Melbourne, VIC, Australia, 1991; Volume 1, pp. 525–542. [Google Scholar]
- Withycombe, C.L. XV. Some aspects of the biology and morphology of the Neuroptera. With special reference to the immature stages and their possible phylogenetic significance. Trans. R. Entomol. Soc. Lond. 1925, 72, 303–411. [Google Scholar] [CrossRef]
- Tillyard, R.J. Studies in Australian Neuroptera. No. The life-history of Psychopsis elegans (Guérin). Proc. Linn. Soc. N. S. W. 1918, 43, 787–818. [Google Scholar]





































Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassenbach, C.; Buchner, L.; Haug, G.T.; Haug, C.; Haug, J.T. An Expanded View on the Morphological Diversity of Long-Nosed Antlion Larvae Further Supports a Decline of Silky Lacewings in the Past 100 Million Years. Insects 2023, 14, 170. https://doi.org/10.3390/insects14020170
Hassenbach C, Buchner L, Haug GT, Haug C, Haug JT. An Expanded View on the Morphological Diversity of Long-Nosed Antlion Larvae Further Supports a Decline of Silky Lacewings in the Past 100 Million Years. Insects. 2023; 14(2):170. https://doi.org/10.3390/insects14020170
Chicago/Turabian StyleHassenbach, Colin, Laura Buchner, Gideon T. Haug, Carolin Haug, and Joachim T. Haug. 2023. "An Expanded View on the Morphological Diversity of Long-Nosed Antlion Larvae Further Supports a Decline of Silky Lacewings in the Past 100 Million Years" Insects 14, no. 2: 170. https://doi.org/10.3390/insects14020170