
DNA Barcoding of Morphologically Characterized Mosquitoes Belonging to the Genus Mansonia from the Atlantic Forest and Brazilian Savanna

 ,
,  , , and
, , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
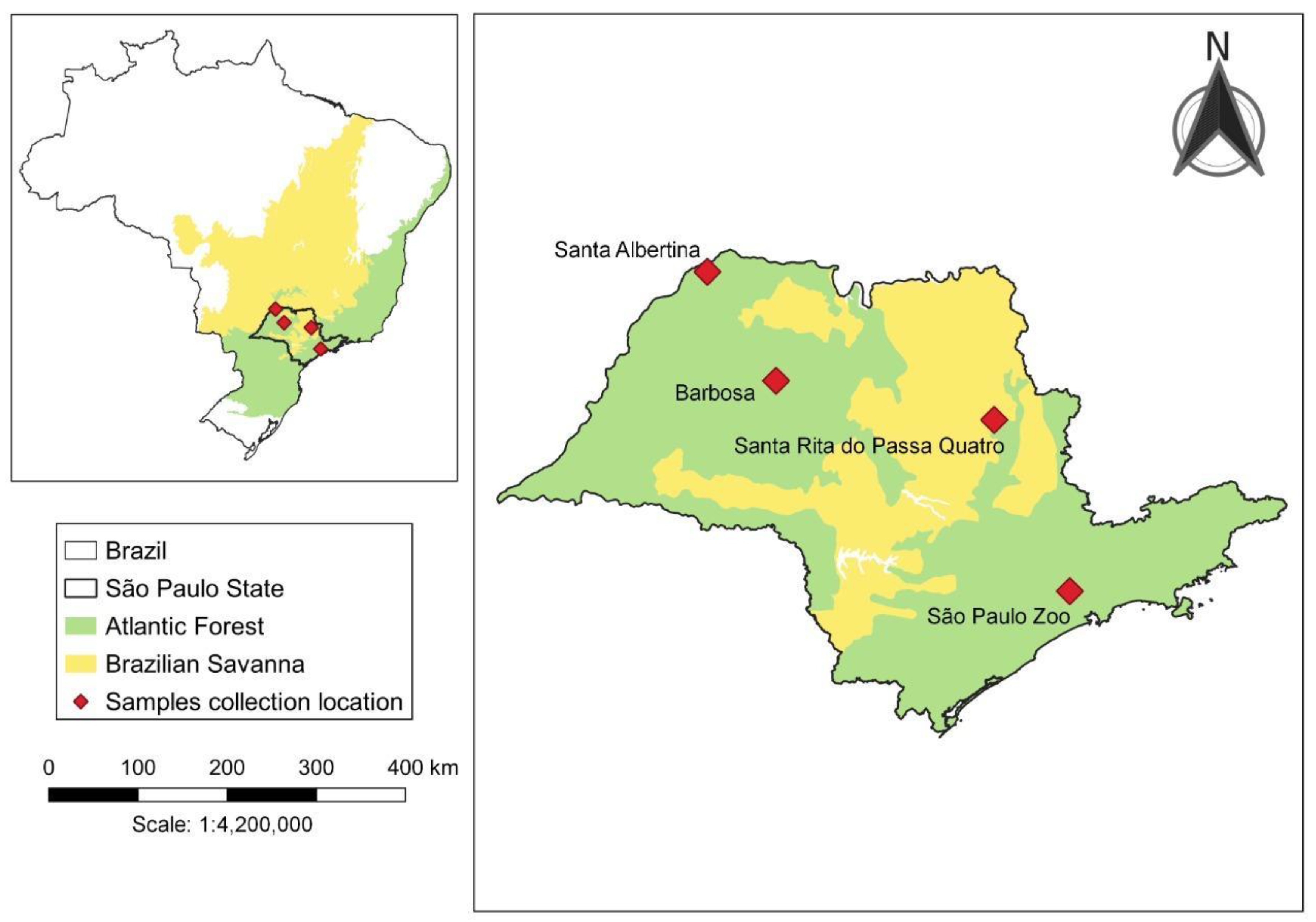
2.1. Mosquito Sampling and Handling
2.2. Genomic DNA Extraction and PCR Amplification of Mitochondrial Gene Fragments
2.3. Sequencing, Alignment, and Sequence Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Collection Year | Period | Method | Strata | Collection Site | Species (According to Taxonomic Keys) |
|---|---|---|---|---|---|---|
| A290E | 2019 | D | Nasci | G | Santa Albertina | Mansonia humeralis |
| A290K | 2019 | D | Nasci | G | Santa Albertina | Mansonia humeralis |
| A290L | 2019 | D | Nasci | G | Santa Albertina | Mansonia humeralis |
| A290M | 2019 | D | Nasci | G | Santa Albertina | Mansonia humeralis |
| A290W | 2019 | D | Nasci | G | Santa Albertina | Mansonia titillans |
| A5725 | 2020 | N | CDC | G | Santa Rita do Passa Quatro | Mansonia (Man.) sp. |
| A613 | 2019 | D | Nasci | G | Barbosa | Mansonia humeralis |
| A613B | 2019 | D | Nasci | G | Barbosa | Mansonia humeralis |
| B173 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia wilsoni |
| B189 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia (Man.) sp. |
| B240 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia (Man.) sp. |
| B245 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia (Man.) sp. |
| B378 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia wilsoni aff |
| B556 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia wilsoni aff |
| B615 | 2020 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| B83 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia (Man.) sp. |
| B926 | 2020 | N | CDC | C | São Paulo Zoo | Mansonia wilsoni aff |
| Zoo044 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo252 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo555 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo634 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo683 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo684 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo685 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo686 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo798 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Zoo799 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia indubitans |
| Zoo800 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| ZooB050 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia pseudotitillans |
| ZooB252 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| ZooB253 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia pseudotitillans |
| ZooB365 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia pseudotitillans |
| ZooB383 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia pseudotitillans |
| ZooB591 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| ZooB592 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia pseudotitillans |
| ZooB797 | 2015 | N | CDC | G | São Paulo Zoo | Mansonia (Man.) sp. |
| Species | Country Source | #GenBank |
|---|---|---|
| Mansonia indubitans | Brazil (Caatinga) | MH118158 |
| Mansonia indubitans | Colombia | MN997669-MN997672 |
| Mansonia flaveola | Puerto Rico | JX260065 |
| Mansonia humeralis | Argentina | MW363430-MW363432 |
| Mansonia titillans | Colombia | KT766533 |
| Mansonia titillans | Colombia | KY859898-KY859902 MN997665-MN997667 |
| Mansonia titillans | Mexico | MN968219, MN968225, MN968231, MN968233, MN968240, MN968241, MN968244, MN968259, MN968266, MN968270, MT999303 |
| Mansonia dyari | Mexico | MN968222, MN968243, MN968246, MN968251, MN968254, MN968264, MN968268, MN968272, MN968273, MN968274 |
| Mansonia dyari | Virgin Islands | MN129182 |
| Mansonia amazonensis | Brazil | MK575483 |
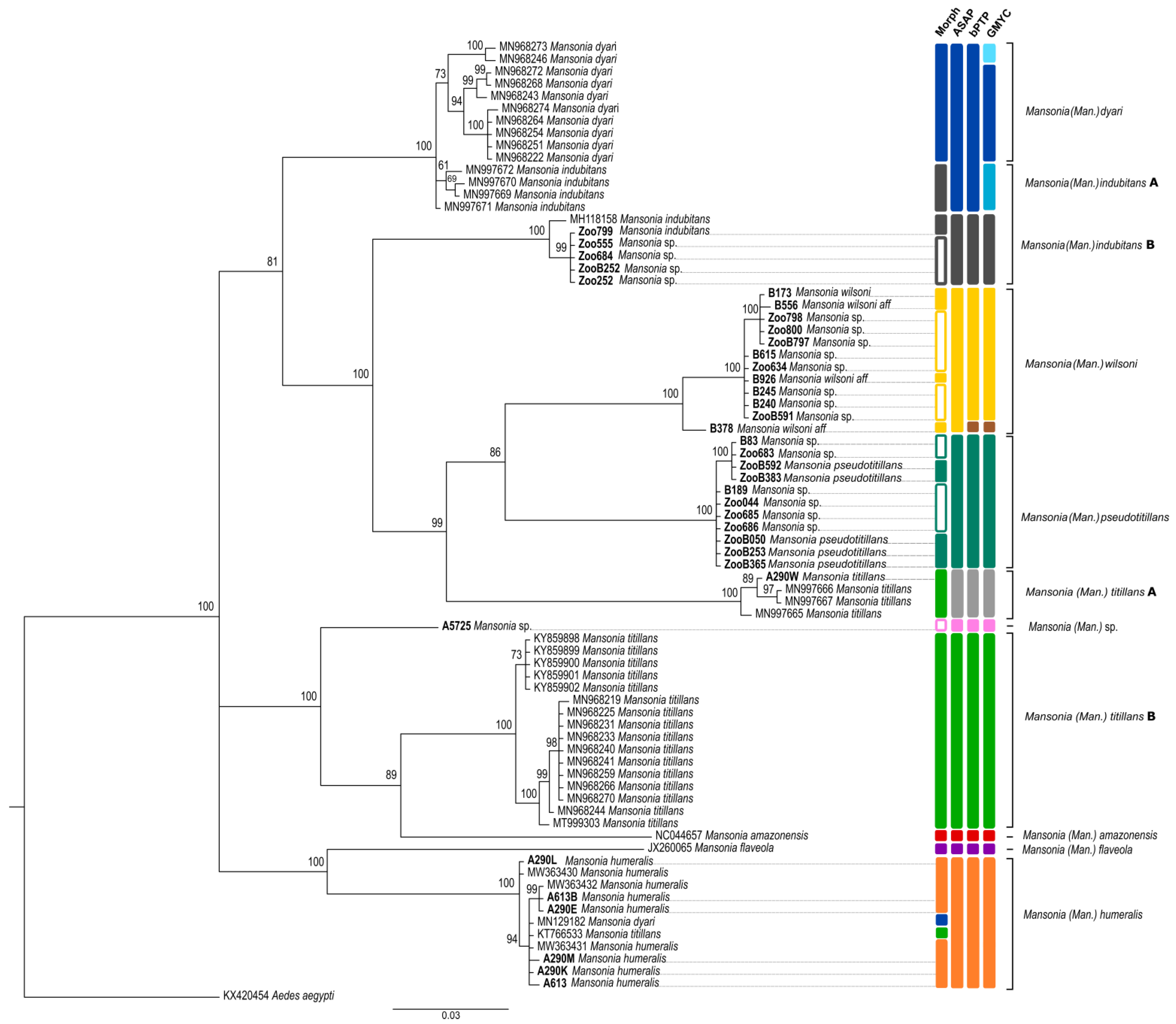
2.4. Species Delimitation
3. Results
3.1. Morphological Assessment
3.1.1. Maxillary Palpus
3.1.2. Spines on the Abdominal Tergite VII and Suprawing Scale
3.2. Sequence Analysis and Species Delimitation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, A.; Chiang, L.-P.; Hapuarachchi, H.C.; Tan, C.-H.; Pang, S.-C.; Lee, R.; Lee, K.-S.; Ng, L.-C.; Lam-Phua, S.-G. DNA Barcoding: Complementing Morphological Identification of Mosquito Species in Singapore. Parasites Vectors 2014, 7, 569. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. Lond. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding Animal Life: Cytochrome c Oxidase Subunit 1 Divergences among Closely Related Species. Proc. R. Soc. Lond. B 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Laurito, M.; de Oliveira, T.M.; Almiron, W.R.; Sallum, M.A.M. COI Barcode versus Morphological Identification of Culex (Culex) (Diptera: Culicidae) Species: A Case Study Using Samples from Argentina and Brazil. Mem. Inst. Oswaldo Cruz 2013, 108, 110–122. [Google Scholar] [CrossRef]
- Laurito, M.; Ayala, A.M.; Arias-Builes, D.L.; Almirón, W.R. Improving the DNA Barcode Library of Mosquito Species with New Identifications and Discoveries in North-Central Argentina. J. Med. Entomol. 2022, 59, 173–183. [Google Scholar] [CrossRef]
- Kirchgatter, K.; de Oliveira Guimarães, L.; Hugo Yañez Trujillano, H.; Rafael Arias, F.; Cáceres, A.; de Castro Duarte, A.; dos Santos Malafronte, R.; Tubaki, R.; Mureb Sallum, M. Phylogeny of Anopheles (Kerteszia) (Diptera: Culicidae) Using Mitochondrial Genes. Insects 2020, 11, 324. [Google Scholar] [CrossRef] [PubMed]
- Zouré, A.A.; Noël, G.; Sombié, A.; Somda, Z.; Badolo, A.; Francis, F. Genetic Analysis and Population Structure of the Anopheles gambiae Complex from Different Ecological Zones of Burkina Faso. Infect. Genet. Evol. 2020, 81, 104261. [Google Scholar] [CrossRef] [PubMed]
- Harbach, R.E.; Kitching, I.J. Phylogeny and Classification of the Culicidae (Diptera). Syst. Entomol. 1998, 23, 327–370. [Google Scholar] [CrossRef]
- Silver, J.B. Mosquito Ecology; Springer: Dordrecht, The Netherlands, 2008; ISBN 978-1-4020-6665-8. [Google Scholar]
- Forattini, O.P. Culicidologia Médica; Publisher of the University of São Paulo: São Paulo, Brazil, 2002; Volume 2. [Google Scholar]
- Ronderos, R.A.; Bachmann, A.O. A proposito del complejo Mansonia (Diptera-Culicidae). Rev. Soc. Entomol. Argent. 1963, 25, 43–51. [Google Scholar]
- Guimarães, J.H. Systematic Database of Diptera of the Americas South of the United States (Family Culicidae); Plêiade/FAPESP: São Paulo, Brazil, 1997. [Google Scholar]
- Barbosa, A.A.; Silva, M.A.N.D.; Sallum, M.A.M. Description and Revalidation of Mansonia (Mansonia) fonsecai (Pinto) (Diptera: Culicidae). Zootaxa 2005, 905, 1–11. [Google Scholar] [CrossRef]
- Barbosa, A.A. Revisão do Subgênero Mansonia Blanchard, 1901 (Diptera, Culicidae) e Estudo Filogenético de Mansoniini. Ph.D. Thesis, Federal University of Paraná, Biological Sciences Sector, Curitiba, Brazil, 2007. [Google Scholar]
- Gaffigan, T.V.; Wilkerson, R.C.; Pecor, J.E.; Stoffer, J.A.; Anderson, T. Systematic Catalog of Culicidae. Walter Reed Biosystematics Unit (WRBU), Division of Entomology, Walter Reed Army Institute of Research (WRAIR), Silver Spring, Maryland, USA. Available online: https://www.mosquitocatalog.org/ (accessed on 3 October 2022).
- Guimarães, L.d.O.; Simões, R.F.; Chagas, C.R.F.; de Menezes, R.M.T.; Silva, F.S.; Monteiro, E.F.; Holcman, M.M.; Bajay, M.M.; Pinter, A.; de Camargo-Neves, V.L.F.; et al. Assessing Diversity, Plasmodium Infection and Blood Meal Sources in Mosquitoes (Diptera: Culicidae) from a Brazilian Zoological Park with Avian Malaria Transmission. Insects 2021, 12, 215. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Jones, J.W.; Sardelis, M.R.; Dohm, D.J.; Coleman, R.E.; Watts, D.M.; Fernandez, R.; Calampa, C.; Klein, T.A. Vector Competence of Peruvian Mosquitoes (Diptera: Culicidae) for Epizootic and Enzootic Strains of Venezuelan Equine Encephalomyelitis Virus. J. Med. Entomol. 2000, 37, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Unlu, I.; Kramer, W.L.; Roy, A.F.; Foil, L.D. Detection of West Nile Virus RNA in Mosquitoes and Identification of Mosquito Blood Meals Collected at Alligator Farms in Louisiana. J. Med. Entomol. 2010, 47, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Méndez, W.; Liria, J.; Navarro, J.-C.; García, C.Z.; Freier, J.E.; Salas, R.; Weaver, S.C.; Barrera, R. Spatial Dispersion of Adult Mosquitoes (Diptera: Culicidae) in a Sylvatic Focus of Venezuelan Equine Encephalitis Virus. J. Med. Entomol. 2001, 38, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Sudia, W.D.; McLEAN, R.G.; Newhouse, V.F.; Johnston, J.G.; Miller, D.L.; Trevino, H.; Bowen, G.S.; Sather, G. Epidemic Venezuelan Equine Encephalitis in North America in 1971: Vertebrate Field Studies. Am. J. Epidemiol. 1975, 101, 36–50. [Google Scholar] [CrossRef]
- Aitken, T.H.G. Habits of Some Mosquito Hosts of VEE (Mucambo) Virus from Northeastern South America, Including Trinidad. In Venezuelan encephalitis; Science Publication 243; Pan American Health Organization: Washington, DC, USA, 1972; pp. 254–256. [Google Scholar]
- Turell, M.J. Vector Competence of Three Venezuelan Mosquitoes (Diptera: Culicidae) for an Epizootic IC Strain of Venezuelan Equine Encephalitis Virus: Table 1. J. Med. Entomol. 1999, 36, 407–409. [Google Scholar] [CrossRef]
- Hoyos-López, R.; Soto, S.U.; Rúa-Uribe, G.; Gallego-Gómez, J.C. Molecular Identification of Saint Louis Encephalitis Virus Genotype IV in Colombia. Mem. Inst. Oswaldo Cruz 2015, 110, 719–725. [Google Scholar] [CrossRef] [Green Version]
- Beranek, M.D.; Gallardo, R.; Almirón, W.R.; Contigiani, M.S. First Detection of Mansonia titillans (Diptera: Culicidae) Infected with St. Louis Encephalitis Virus (Flaviviridae: Flavivirus) and Bunyamwera Serogroup (Peribunyaviridae: Orthobunyavirus) in Argentina. J. Vector Ecol. 2018, 43, 340–343. [Google Scholar] [CrossRef] [Green Version]
- Maes, P.; Alkhovsky, S.V.; Bào, Y.; Beer, M.; Birkhead, M.; Briese, T.; Buchmeier, M.J.; Calisher, C.H.; Charrel, R.N.; Choi, I.R.; et al. Taxonomy of the Family Arenaviridae and the order Bunyavirales: Update 2018. Arch. Virol. 2018, 163, 2295–2310. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, F.C.; Rodrigues, R.A.; Sato, Y.; Borges, M.A.Z.; Braga, É.M. Searching for Putative Avian Malaria Vectors in a Seasonally Dry Tropical Forest in Brazil. Parasites Vectors 2016, 9, 587. [Google Scholar] [CrossRef] [Green Version]
- Sudia, W.D.; Chamberlain, R.W. Battery-Operated Light Trap, an Improved Model. By W. D. Sudia and R. W. Chamberlain, 1962. J. Am. Mosq. Control Assoc. 1988, 4, 536–538. [Google Scholar]
- Nasci, R.S. A Lightweight Battery-Powered Aspirator for Collecting Resting Mosquitoes in the Field. Mosq. News 1981, 41, 808–811. [Google Scholar]
- Consoli, R.A.G.B.; Lourenço-de-Oliveira, R. Principais Mosquitos de Importância Sanitária No Brasil; Cadernos de Saúde Pública; FIOCRUZ: Rio de Janeiro, Brazil, 1994. [Google Scholar]
- Assumpção, I.C.D. Chave de identificação pictórica para o subgênero Mansonia Blanchard, 1901 (Diptera, Culicidae) da região neotropical. In Biological Sciences Sector; Federal University of Paraná: Curitiba, Brazil, 2009; Volume 41, p. 34. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Ruiz, F.; Linton, Y.-M.; Ponsonby, D.J.; Conn, J.E.; Herrera, M.; Quiñones, M.L.; Vélez, I.D.; Wilkerson, R.C. Molecular Comparison of Topotypic Specimens Confirms Anopheles (Nyssorhynchus) dunhami Causey (Diptera: Culicidae) in the Colombian Amazon. Mem. Inst. Oswaldo Cruz 2010, 105, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J. The CLUSTAL_X Windows Interface: Flexible Strategies for Multiple Sequence Alignment Aided by Quality Analysis Tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Reinert, J.E. Revised List of Abbreviations for Genera and Subgenera of Culicidae (Diptera) and Notes on Generic and Subgeneric Changes. J. Am. Mosq. Control Assoc. 2001, 17, 51–55. [Google Scholar]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree: Tree Figure Drawing Tool. Version 1.4.0. Institute of Evolutionary Biology, University of Edinburgh. 2006–2012. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 22 May 2019).
- Kimura, M. A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Fontaneto, D.; Hortal, J. At Least Some Protist Species Are Not Ubiquitous. Mol. Ecol. 2013, 22, 5053–5055. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble Species by Automatic Partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, T.; Barraclough, T.G. Delimiting Species Using Single-Locus Data and the Generalized Mixed Yule Coalescent Approach: A Revised Method and Evaluation on Simulated Data Sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forattini, O.P. Entomologia Médica. Culicini: Haemagogus, Mansonia, Culiseta, Sabethini, Toxorhynchitini, Arboviroses, Filariose Bancroftiana. Genética; Publisher of the University of São Paulo: São Paulo, Brazil, 1965; Volume 3. [Google Scholar]
- Belkin, J.N.; Heinemann, S.J.; Page, W.A. Mosquito studies (Diptera: Culicidae) XXI. The Culicidae of Jamaica. Contrib. Am. Entomol. Inst. 1970, 6, 1–458. [Google Scholar]
- Scarpassa, V.M.; Batista, E.T.; Ferreira, V.C.; Santos Neto, V.A.; Roque, R.A.; Tadei, W.P.; Ferreira, F.A.S.; Costa, F.M. DNA Barcoding Suggests New Species for the Mansonia Subgenus (Mansonia, Mansoniini, Culicidae, Diptera) in the Area Surrounding the Jirau Hydroelectric Dam, Porto Velho Municipality, Rondônia State, Brazil. Acta Trop. 2022, 233, 106574. [Google Scholar] [CrossRef]
- Barbosa, A.A.; Da Silva, M.A.N.; Sallum, M.A.M. Mansonia (Mansonia) iguassuensis Sp. Nov. (Diptera: Culicidae) from Brasil. Zootaxa 2007, 1527, 45–52. [Google Scholar] [CrossRef]
- LANE, C.J. On a Collection of Culicinae (Diptera: Culicidae) from Brazil. Mosq. Syst. 1992, 24, 16–22. [Google Scholar]





| Species | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Ma. wilsoni | 0.002 | ||||||||||||
| 2 | Ma. humeralis | 0.161 | 0.003 | |||||||||||
| 3 | Ma. titillans A | 0.148 | 0.158 | 0.005 | ||||||||||
| 4 | Mansonia sp. A5725 | 0.148 | 0.126 | 0.142 | ||||||||||
| 5 | Ma. pseudotitillans | 0.112 | 0.149 | 0.132 | 0.153 | 0.002 | ||||||||
| 6 | Ma. wilsoni aff B378 | 0.021 | 0.149 | 0.135 | 0.138 | 0.103 | ||||||||
| 7 | Ma. flaveola | 0.160 | 0.136 | 0.183 | 0.149 | 0.151 | 0.155 | |||||||
| 8 | Aedes aegypti | 0.152 | 0.165 | 0.189 | 0.147 | 0.172 | 0.150 | 0.174 | ||||||
| 9 | Ma. titillans B | 0.143 | 0.140 | 0.134 | 0.073 | 0.142 | 0.134 | 0.151 | 0.164 | 0.005 | ||||
| 10 | Ma. indubitans B | 0.124 | 0.134 | 0.133 | 0.120 | 0.112 | 0.113 | 0.155 | 0.155 | 0.126 | 0.003 | |||
| 11 | Ma. dyari | 0.124 | 0.139 | 0.123 | 0.108 | 0.118 | 0.111 | 0.144 | 0.137 | 0.107 | 0.110 | 0.010 | ||
| 12 | Ma. indubitans A | 0.124 | 0.133 | 0.120 | 0.105 | 0.119 | 0.111 | 0.148 | 0.139 | 0.108 | 0.109 | 0.013 | 0.004 | |
| 13 | Ma. amazonensis | 0.140 | 0.154 | 0.148 | 0.104 | 0.139 | 0.133 | 0.162 | 0.162 | 0.096 | 0.128 | 0.127 | 0.122 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirchgatter, K.; Guimarães, L.d.O.; Monteiro, E.F.; Helfstein, V.C.; Telles-de-Deus, J.; Menezes, R.M.T.d.; Reginato, S.L.; Chagas, C.R.F.; de Camargo-Neves, V.L.F. DNA Barcoding of Morphologically Characterized Mosquitoes Belonging to the Genus Mansonia from the Atlantic Forest and Brazilian Savanna. Insects 2023, 14, 109. https://doi.org/10.3390/insects14020109
Kirchgatter K, Guimarães LdO, Monteiro EF, Helfstein VC, Telles-de-Deus J, Menezes RMTd, Reginato SL, Chagas CRF, de Camargo-Neves VLF. DNA Barcoding of Morphologically Characterized Mosquitoes Belonging to the Genus Mansonia from the Atlantic Forest and Brazilian Savanna. Insects. 2023; 14(2):109. https://doi.org/10.3390/insects14020109
Chicago/Turabian StyleKirchgatter, Karin, Lilian de Oliveira Guimarães, Eliana Ferreira Monteiro, Vanessa Christe Helfstein, Juliana Telles-de-Deus, Regiane Maria Tironi de Menezes, Simone Liuchetta Reginato, Carolina Romeiro Fernandes Chagas, and Vera Lucia Fonseca de Camargo-Neves. 2023. "DNA Barcoding of Morphologically Characterized Mosquitoes Belonging to the Genus Mansonia from the Atlantic Forest and Brazilian Savanna" Insects 14, no. 2: 109. https://doi.org/10.3390/insects14020109