
Efficacy of Entomopathogenic Fungi as Prevention against Early Life Stages of the Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in Laboratory and Greenhouse Trials



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Entomopathogenic Fungi
2.2. Insects
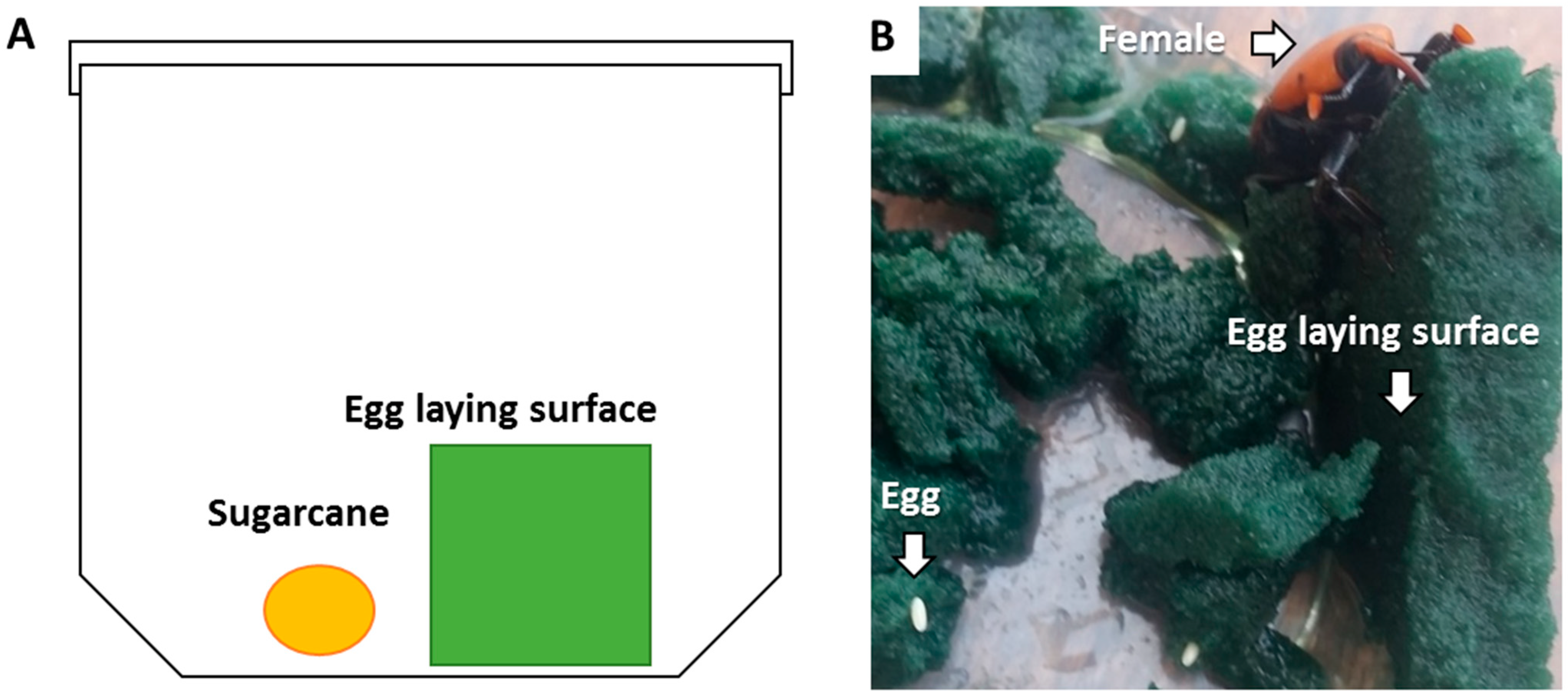
2.3. Laboratory Bioassay
2.4. Greenhouse Experiment
2.5. Data Analysis
3. Results
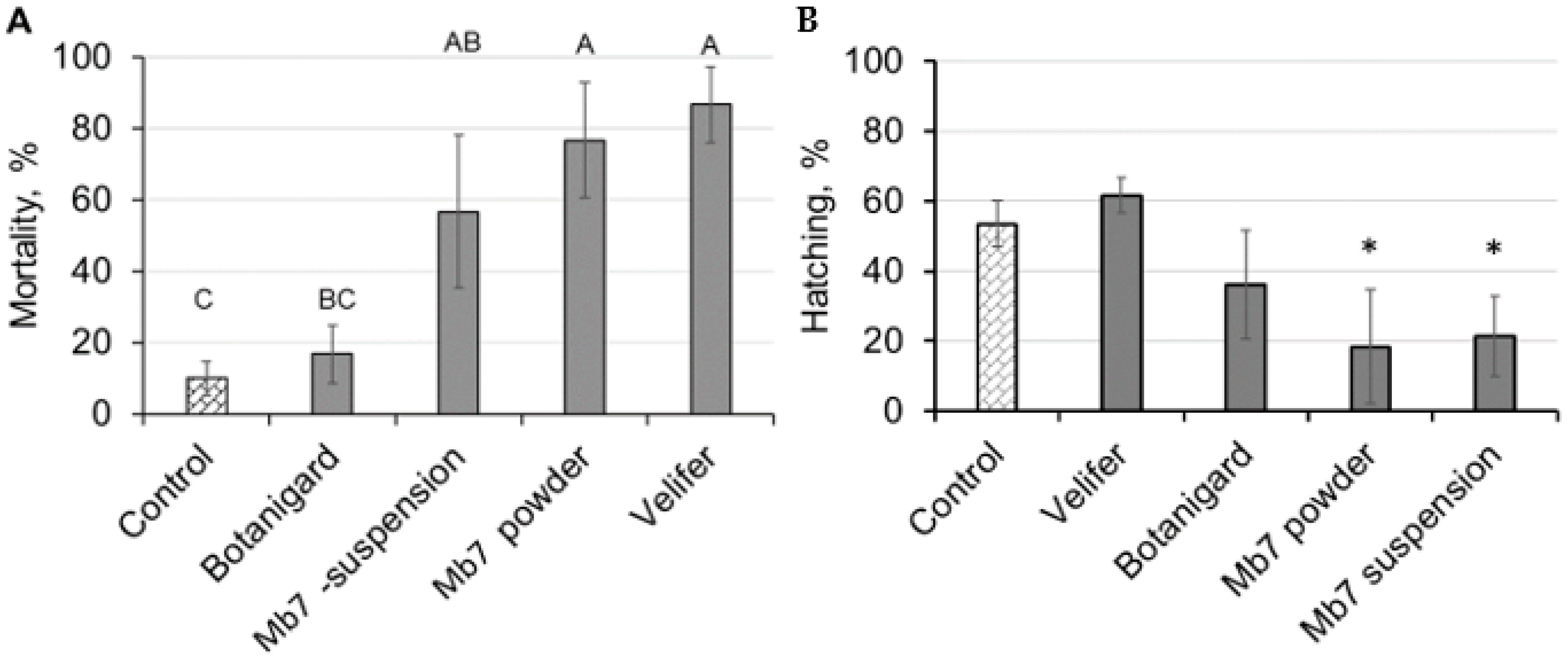
3.1. Laboratory Experiments
3.2. Greenhouse Experiments
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. Red Palm Weevil: Guidelines on Management Practices. 2020. Available online: https://www.fao.org/publications/card/en/c/CA7703EN/ (accessed on 7 October 2023).
- Soroker, V.; Blumberg, D.; Haberman, A.; Hamburger-Rishard, M.; Reneh, S.; Talebaev, S.; Anshelevich, L.; Harari, A.R. Current status of red palm weevil infestation in date palm plantations in Israel. Phytoparasitica 2005, 33, 97–106. [Google Scholar] [CrossRef]
- Blumberg, D. Review: Date palm arthropod pests and their management in Israel. Phytoparasitica 2008, 36, 411–418. [Google Scholar] [CrossRef]
- Llácer, E.; Dembilio, O.; Jacas, J.A. Evaluation of the efficacy of an insecticidal paint based on chlorpyrifos and pyriproxyfen in a microencapsulated formulation against Rhynchophorus ferrugineus (Coleoptera: Curculionidae). J. Econ. Entomol. 2010, 103, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dembilio, Ó.; Llácer, E.; Martínez de Altube, M.d.M.; Jacas, J.A. Field efficacy of imidacloprid and Steinernema carpocapsae in a chitosan formulation against the red palm weevil Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in Phoenix canariensis. Pest Manag. Sci. 2010, 66, 365–370. [Google Scholar] [CrossRef]
- Israeli Extension Service. Management of the Red Palm Weevil, Professional Information. 2017. Available online: https://www.moag.gov.il/shaham/ProfessionalInformation/Pages/meniat_vadbarat_chedkonit_hadekel_june_2017.aspx (accessed on 7 October 2023).
- Mendel, Z.; Voet, H.; Modan, N.; Naor, R.; Ment, D. Seismic sensor-based management of the red palm weevil Rhynchophorus ferrugineus in date palm plantations. Pest Manag. Sci. 2023. [Google Scholar] [CrossRef] [PubMed]
- Mazza, G.; Francardi, V.; Simoni, S.; Benvenuti, C.; Cervo, R.; Faleiro, J.R.; Llácer, E.; Longo, S.; Nannelli, R.; Tarasco, E.; et al. An overview on the natural enemies of Rhynchophorus palm weevils, with focus on R. ferrugineus. Biol. Control 2014, 77, 83–92. [Google Scholar] [CrossRef]
- Ortega-García, L.; Tabone, E.; Beaudoin-Ollivier, L.; Ment, D.; Buradino, M.; Jaques, J.A.; Garrido-Jurado, I.; Dembilio, O.; Moraga, E.Q. Natural enemies of Rhynchophorus ferrugineus and Paysandisia archon. In Handbook of Major Palm Pests; Soroker, V., Colazza, S., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2017; pp. 171–186. [Google Scholar] [CrossRef]
- Nurashikin-Khairuddin, W.; Abdul-Hamid, S.N.A.; Mansor, M.S.; Bharudin, I.; Othman, Z.; Jalinas, J. A review of entomopathogenic nematodes as a biological control agent for red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Insects 2022, 13, 245. [Google Scholar] [CrossRef]
- Santhi, V.S.; Salame, L.; Nakache, Y.; Koltai, H.; Soroker, V.; Glazer, I. Attraction of entomopathogenic nematodes Steinernema carpocapsae and Heterorhabditis bacteriophora to the red palm weevil (Rhynchophorus ferrugineus). Biol. Control 2015, 83, 75–81. [Google Scholar] [CrossRef]
- Wakil, W.; Yasin, M.; Shapiro-Ilan, D. Effects of single and combined applications of entomopathogenic fungi and nematodes against Rhynchophorus ferrugineus (Olivier). Sci. Rep. 2017, 7, 5971. [Google Scholar] [CrossRef]
- Yaacobi, G.; Salame, L.; Glazer, I. Persistence of the entomopathogenic nematode Steinernema carpocapsae on red palm weevil-infested date palm trees in an arid environment. Nematology 2023, 25, 669–675. [Google Scholar] [CrossRef]
- Cito, A.; Mazza, G.; Strangi, A.; Benvenuti, C.; Barzanti, G.P.; Dreassi, E.; Turchetti, T.; Francardi, V.; Roversi, P.F. Characterization and comparison of Metarhizium strains isolated from Rhynchophorus ferrugineus. FEMS Microbiol. Lett. 2014, 355, 108–115. [Google Scholar] [CrossRef] [PubMed]
- Dembilio, Ó.; Quesada-Moraga, E.; Santiago-Álvarez, C.; Jacas, J.A. Potential of an indigenous strain of the entomopathogenic fungus Beauveria bassiana as a biological control agent against the Red Palm Weevil, Rhynchophorus ferrugineus. J. Invertebr. Pathol. 2010, 104, 214–221. [Google Scholar] [CrossRef] [PubMed]
- El-Sufty, R.; Al Bgham, S.; Al-Awash, S.; Shahdad, A.; Al Bathra, A. A trap for auto-dissemination of the entomopathogenic fungus Beauveria bassiana by red palm weevil adults in date palm plantations. Egypt. J. Biol. Pest Control 2011, 21, 271–276. [Google Scholar]
- El-Sufty, R.; Al-Awash, S.A.; Al Amiri, A.M.; Shahdad, A.S.; Al Bathra, A.H.; Musa, S.A. Biological control of red palm weevil, Rhynchophorus ferrugineus (Col.: Curculionidae) by the entomopathogenic fungus Beauveria bassiana in United Arab Emirates. Acta Hortic. 2007, 736, 399–404. [Google Scholar] [CrossRef]
- Gindin, G.; Levski, S.; Glazer, I.; Soroker, V. Evaluation of the entomopathogenic fungi Metarhizium anisopliae and Beauveria bassiana against the red palm weevil Rhynchophorus ferrugineus. Phytoparasitica 2006, 34, 370–379. [Google Scholar] [CrossRef]
- Sabbour, M.M.; Abdel-Raheem, M.A. Evaluations of Isaria fumosorosea isolates against the Red Palm Weevil Rhynchophorus ferrugineus under laboratory and field conditions. Curr. Sci. Int. 2014, 3, 179–185. [Google Scholar] [CrossRef]
- Sabbour, M.M.; Solieman, N.Y. Preliminary investigations into the biological control of Red Palm Weevil Rhynchophorus ferrugineus by using three isolates of the fungus Lecanicillium (Verticillium) lecanii in Egypt. Int. J. Sci. Res. 2014, 3, 2060–2066. [Google Scholar]
- Güerri-Agulló, B.; López-Follana, R.; Asensio, L.; Barranco, P.; Lopez-Llorca, L.V. Use of a solid formulation of Beauveria bassiana for biocontrol of the red palm weevil (Rhynchophorus ferrugineus) (Coleoptera: Dryophthoridae) under field conditions in SE Spain. Fla. Entomol. 2011, 94, 737–747. [Google Scholar] [CrossRef]
- Dembilio, Ó.; Moya, P.; Vacas, S.; Ortega-García, L.; Quesada-Moraga, E.; Jaques, J.A.; Navarro-Llopis, V. Development of an attract-and-infect system to control Rhynchophorus ferrugineus with the entomopathogenic fungus Beauveria bassiana. Pest Manag. Sci. 2018, 74, 1861–1869. [Google Scholar] [CrossRef]
- Matveev, S.; Reingold, V.; Yossef, E.; Levy, N.; Kottakota, C.; Mechrez, G.; Protasov, A.; Belausov, E.; Birnbaum, N.; Davidovitz, M.; et al. The dissemination of Metarhizium brunneum conidia by females of the red palm weevil, Rhynchophorus ferrugineus, from treated surface to laying hole reduces egg hatch and larval survival, suggesting a new mechanism for prevention practic-es. J. Fungi 2023, 9, 458. [Google Scholar] [CrossRef]
- Francardi, V.; Benvenuti, C.; Barzanti, G.P.; Roversi, P.F. Autocontamination trap with entomopathogenic fungi: A possible strategy in the control of Rhynchophorus ferrugineus (Olivier) (Coleoptera Curculionidae). Redia 2013, 96, 57–67. [Google Scholar]
- Najar-Rodríguez, A.J.; Lavidis, N.A.; Mensah, R.K.; Choy, P.T.; Walter, G.H. The toxicological effects of petroleum spray oils on insects—Evidence for an alternative mode of action and possible new control options. Food Chem. Toxicol. 2008, 46, 3003–3014. [Google Scholar] [CrossRef]
- Buteler, M.; Stadler, T.A. Review on the Mode of Action and Current Use of Petroleum Distilled Spray Oils. In Pesticides in the Modern World; Stoycheva, M., Ed.; InTech: Rijeka, Croatia, 2011; pp. 119–136. ISBN 978-953-307-459-7. [Google Scholar]
- Birnbaum, N.; Reingold, V.; Matveev, S.; Kottakota, C.; Davidovitz, M.; Mani, K.A.; Feldbaum, R.; Yaakov, N.; Mechrez, G.; Ment, D. Not Only a Formulation: The Effects of Pickering Emulsion on the Entomopathogenic Action of Metarhizium Brunneum. J. Fungi 2021, 7, 499. [Google Scholar] [CrossRef]
- Ment, D.; Churchill, A.C.L.; Gindin, G.; Belausov, E.; Glazer, I.; Rehner, S.A.; Rot, A.; Donzelli, B.G.G.; Samish, M. Resistant ticks inhibit Metarhizium infection prior to haemocoel invasion by reducing fungal viability on the cuticle surface. Environ. Microbiol. 2012, 14, 1570–1583. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A. The role of epigenetics in host–parasite coevolution: Lessons from the model host insects Galleria mellonella and Tribolium castaneum. Zoology 2016, 119, 273–280. [Google Scholar] [CrossRef]
- Wang, J.; St Lovett, B.; Leger, R.J. The secretome and chemistry of Metarhizium; a genus of entomopathogenic fungi. Fungal Ecol. 2018. [Google Scholar] [CrossRef]
- Reingold, V.; Staropoli, A.; Faigenboim, A.; Maymone, M.; Matveev, S.; Keppanan, R.; Ghanim, M.; Vinale, F.; Ment, D. The SWC4 subunit of the SWR1 chromatin remodeling complex is involved in varying virulence of Metarhizium brunneum isolates offering role of epigenetic regulation of pathogenicity. Virulence 2022, 13, 1252–1269. [Google Scholar] [CrossRef]
- Gindin, G.; Ment, D.; Rot, A.; Glazer, I.; Samish, M. Pathogenicity of Metarhizium anisopliae (Hypocreales: Clavicipitaceae) to tick eggs and the effect of egg cuticular lipids on conidia development. J. Med. Entomol. 2009, 46, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Mondaca, L.L.; Da-Costa, N.; Protasov, A.; Ben-Yehuda, S.; Peisachov, A.; Mendel, Z.; Ment, D. Activity of Metarhizium brunneum and Beauveria bassiana against early developmental stages of the false codling moth. J. Invert. Pathol. 2019, 170, 107312. [Google Scholar] [CrossRef]
- Ricaño, J.; Güerri-Agulló, B.; Serna-Sarriás, M.J.; Rubio-Llorca, G.; Asensio, L.; Barranco, P.; Lopez-Llorca, L.V. Evaluation of the pathogenicity of multiple isolates of Beauveria bassiana (Hypocreales: Clavicipitaceae) on Rhynchophorus ferrugineus (Coleoptera: Dryophthoridae) for the assessment of a solid formulation under simulated field conditions. Fla. Entomol. 2013, 96, 1311–1324. [Google Scholar] [CrossRef]
- Prior, C.; Arura, M. The infectivity of Metarhizium anisopliae to two insect pests of coconuts. J. Invertebr. Pathol. 1985, 45, 187–194. [Google Scholar] [CrossRef]
- Hajjar, M.J.; Ajlan, A.M.; Al-Ahmad, M.H. New approach of Beauveria bassiana to control the red palm weevil (Coleoptera: Curculionidae) by trapping technique. J. Econ. Entomol. 2015, 108, 425–432. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Country of Origin | Source | Trade Name | Formulation |
|---|---|---|---|---|---|
| Beauveria bassiana | PPRI 5339 | South Africa | BASF | Velifer | Emulsified concentrate |
| Beauveria bassiana | GHA | USA | LAM International Corporation | Botanigard | Emulsified concentrate |
| Metarhizium brunneum | Mb7 * | Israel | Sporulated rice produced in the laboratory | None | (1) Dry conidia granules (2) Conidial suspension |
| Treatment | Secretion (%) | Signs of Boring (%) | Cumulative Mortality of Adults | Total Larvae per Palm | Total Pupae per Palm |
|---|---|---|---|---|---|
| Control | 100 | 75 | 6.25 ± 2.5 a | 8.5 ± 3.3 a | 1.5 ± 1.5 a |
| Conidial suspension | 25 | 0 | 10.5 ± 2.9 a | 1 ± 1.9 ab | 3 ± 1.8 a |
| Sporulated rice | 0 | 0 | 10 ± 6.4 a | 0 b | 0 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ment, D.; Levy, N.; Allouche, A.; Davidovitz, M.; Yaacobi, G. Efficacy of Entomopathogenic Fungi as Prevention against Early Life Stages of the Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in Laboratory and Greenhouse Trials. Insects 2023, 14, 918. https://doi.org/10.3390/insects14120918
Ment D, Levy N, Allouche A, Davidovitz M, Yaacobi G. Efficacy of Entomopathogenic Fungi as Prevention against Early Life Stages of the Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in Laboratory and Greenhouse Trials. Insects. 2023; 14(12):918. https://doi.org/10.3390/insects14120918
Chicago/Turabian StyleMent, Dana, Noa Levy, Arnon Allouche, Michael Davidovitz, and Gal Yaacobi. 2023. "Efficacy of Entomopathogenic Fungi as Prevention against Early Life Stages of the Red Palm Weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae) in Laboratory and Greenhouse Trials" Insects 14, no. 12: 918. https://doi.org/10.3390/insects14120918