
Evaluation of Black Soldier Fly Hermetia illucens as Food for Pink-Spotted Lady Beetle Coleomegilla maculata

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Colonies
2.2. Experimental Design and Diet Treatments, Experiment 1
2.3. Experimental Design and Diet Treatments, Experiment 2
2.4. Statistical Analysis
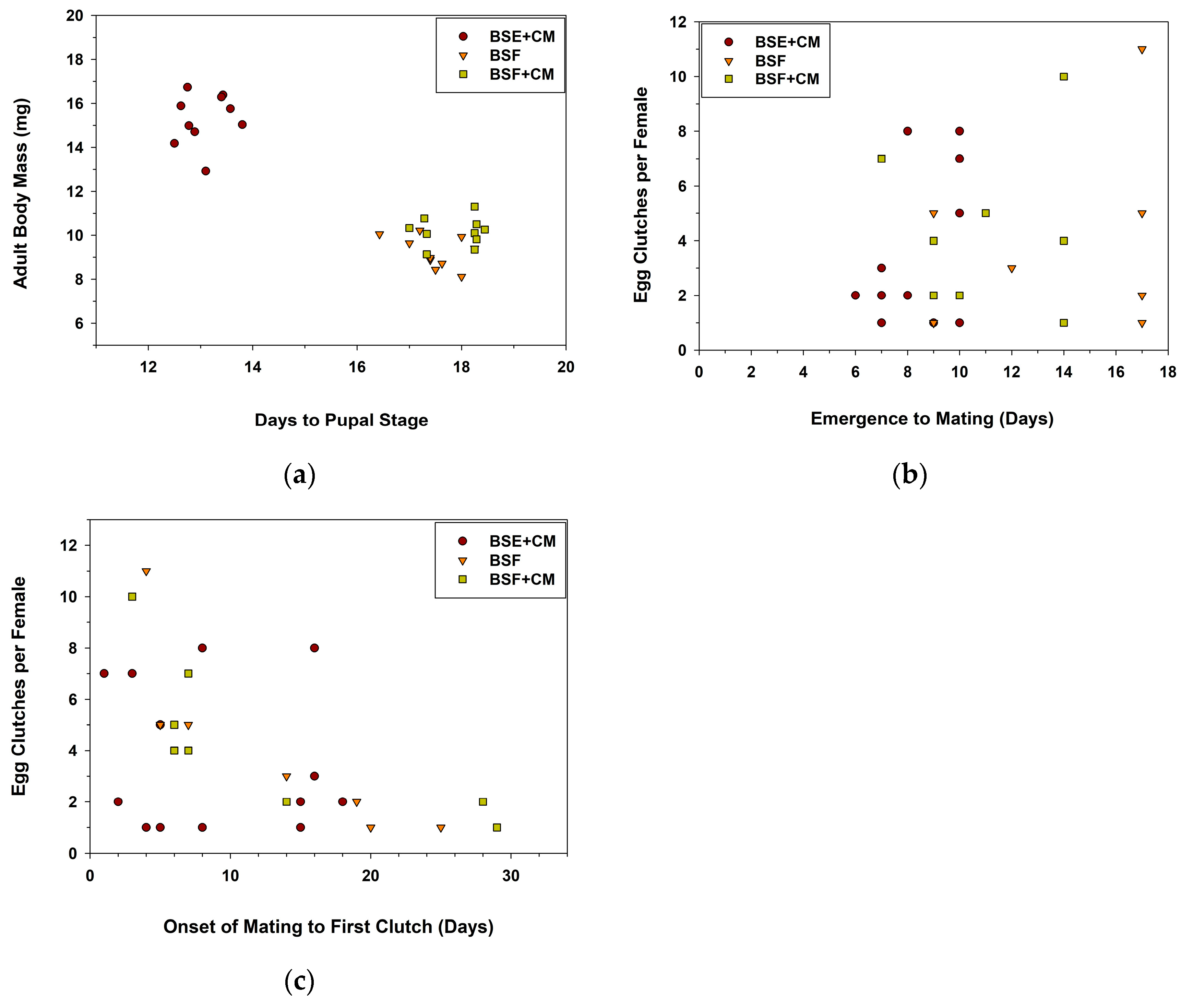
3. Results
3.1. Experiment 1
3.2. Experiment 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hagen, K.S.; Mills, N.J.; Gordh, G.; McMurtry, J.A. Terrestrial arthropod predators of insect and mite pests. In Handbook of Biological Control: Principles and Applications of Biological Control; Bellows, T.S., Fisher, T.W., Eds.; Academic Press: San Diego, CA, USA, 1999; Chapter 16; pp. 383–503. [Google Scholar]
- van Lenteren, J.C.; Hale, A.; Klapwijk, J.N.; van Schelt, J.; Steinberg, S. Guidelines for quality control of commercially produced natural enemies. In Quality Control and Production of Biological Control Agents: Theory and Testing Procedures; van Lenteren, J.C., Ed.; CABI Publishing: Oxon, UK, 2003; Chapter 19; pp. 265–303. [Google Scholar]
- Riddick, E.W. Benefits and limitations of factitious prey and artificial diets on life parameters of predatory beetles, bugs, and lacewings: A mini-review. BioControl 2009, 54, 325–339. [Google Scholar] [CrossRef]
- Sun, Y.X.; Hao, Y.N.; Riddick, E.W.; Liu, T.X. Factitious prey and artificial diets for predatory lady beetles: Current situation, obstacles, and approaches for improvement. Biocontrol Sci. Technol. 2017, 27, 601–619. [Google Scholar] [CrossRef]
- Riddick, E.W. Production of coleopteran predators. In Mass Production of Beneficial Organisms, 2nd ed.; Morales-Ramos, J.A., Rojas, M.G., Shapiro Ilan, D.I., Eds.; Academic Press: London, UK, 2023; Chapter 2; pp. 13–36. [Google Scholar]
- Ricupero, M.; Dai, C.; Siscaro, C.; Russo, A.; Biondi, A.; Zappalá, L. Potential diet regimes for laboratory rearing on the harlequin ladybird. BioControl 2020, 65, 583–592. [Google Scholar] [CrossRef]
- Li, D.; Chen, P.; Liu, J.; Chi, B.; Zhang, X.; Liu, Y. Artificial rearing of the ladybird Propylea japonica on a diet containing Oriental armyworm, Mythimna separata. Entomol. Exp. Appl. 2021, 169, 472–479. [Google Scholar] [CrossRef]
- Montoro, M.; Fine Licht, H.H.; Sigsgaard, L. Nutritional quality of Drosophila melanogaster as factitious prey for rearing the predatory bug Orius majusculus. Insect Sci. 2021, 28, 191–202. [Google Scholar] [CrossRef]
- Vahmani, A.; Shirvani, A.; Rashki, M. Biological parameters of Oenopia conglobata (Linnaeus) (Coleoptera: Coccinellidae) feeding on different diets. Int. J. Trop. Insect Sci. 2022, 42, 2241–2247. [Google Scholar] [CrossRef]
- Ovchinnikov, A.N.; Ovchinnikova, A.A.; Reznik, S.Y.; Belyakova, N.A. Signal and nutritional effects of mixed diets on reproduction of a predatory ladybird, Cheilomenes propinqua. Insects 2023, 14, 587. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhou, Y.; Ran, H.; Li, F. Effects of different hormones as dietary supplements on biological characteristics of Coccinella septempunctata L. J. Appl. Entomol. 2023. [Google Scholar] [CrossRef]
- Tomberlin, J.K.; van Huis, A. Black soldier fly from pest to ‘crown jewel’ of the insects as feed industry: An historical perspective. J. Insects Food Feed 2020, 6, 1–4. [Google Scholar] [CrossRef]
- van Huis, A. Potential of insects as food and feed in assuring food security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- van Huis, A.; Dicke, M.; van Loon, J.J.A. Insects to feed the world. J. Insects Food Feed 2015, 1, 3–5. [Google Scholar] [CrossRef]
- Kaczor, M.; Bulak, P.; Proc-Pietrycha, K.; Kirichenko-Babko, M.; Bieganowski, A. The variety of applications of Hermetia illucens in industrial and agricultural areas—Review. Biology 2023, 12, 25. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.; Taethaisong, N.; Meethip, W.; Surakhunthod, J.; Sinpru, B.; Sroichak, T.; Archa, P.; Thongpea, S.; Paengkoum, S.; Purba, R.A.P.; et al. Nutritional composition of black soldier fly larvae (Hermetia illucens L.) and its potential uses as alternative protein sources in animal diets: A review. Insects 2022, 13, 831. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Dong, Y.; Sun, Q.; Tan, X.; You, C.; Huang, Y.; Zhou, M. Growth and fatty acid composition of black soldier fly Hermetia illucens (Diptera: Stratiomyidae) larvae are influenced by dietary fat sources and levels. Animals 2022, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Mshayisa, V.V.; Van Wyk, J.; Zozo, B. Nutritional, techno-functional and structural properties of black soldier fly (Hermetia illucens) larvae flours and protein concentrates. Foods 2022, 11, 724. [Google Scholar] [CrossRef]
- Hu, Z.; Li, H.; Liu, S.; Xue, R.; Sun, J.; Ji, H. Assessment of black soldier fly (Hermetia illucens) larvae meal as a potential substitute for soybean meal on growth performance and flesh quality of grass carp Ctenopharyngodon idellus. Anim. Nutr. 2023, 14, 425–449. [Google Scholar] [CrossRef]
- Zarantoniello, M.; Chemello, G.; Ratti, S.; Pulido-Rodríguez, L.F.; Daniso, E.; Freddi, L.; Salinetti, P.; Nartea, A.; Bruni, L.; Parisi, G.; et al. Growth and welfare status of giant freshwater prawn (Macrobrachium rosenbergii) post-larvae reared in aquaponic systems and fed diets including enriched black soldier fly (Hermetia illucens) prepupae meal. Animals 2023, 13, 715. [Google Scholar] [CrossRef]
- Riddick, E.W. Insect protein as a partial replacement for fishmeal in the diets of juvenile fish and crustaceans. In Mass Production of Beneficial Organisms; Morales-Ramos, J.A., Rojas, M.G., Shapiro Ilan, D.I., Eds.; Academic Press: London, UK, 2014; Chapter 16; pp. 565–582. [Google Scholar]
- Nguyen, D.T.; Bouguet, V.; Spranghers, T.; Vangansbeke, D.; De Clercq, P. Beneficial effect of supplementing an artificial diet for Amblyseius swirskii with Hermetia illucens haemolymph. J. Appl. Entomol. 2015, 139, 342–351. [Google Scholar] [CrossRef]
- Gordon, R.D. The Coccinellidae (Coleoptera) of America north of Mexico. J. N. Y. Entomol. Soc. 1985, 93, 1–912. [Google Scholar]
- Krafsur, E.S.; Obrycki, J.J. Coleomegilla maculata (Coleoptera: Coccinellidae) is a species complex. Ann. Entomol. Soc. Am. 2000, 93, 1156–1163. [Google Scholar] [CrossRef]
- McAlpine, D.F.; Migneault, R.; Webster, R.P. Coleomegilla maculata lengi Timberlake, 1943 (Coleoptera: Coccinellidae), a native North American lady beetle new to Maritime Canada. J. Acad. Entomol. 2018, 14, 8–10. [Google Scholar]
- Lucas, E.; Labrecque, C.; Coderre, D. Delphastus catalinae and Coleomegilla maculata lengi (Coleoptera: Coccinellidae) as biological control agents of the greenhouse whitefly, Trialeurodes vaporariorum (Homoptera: Aleyrodidae). Pest Manag. Sci. 2004, 60, 1073–1078. [Google Scholar] [CrossRef] [PubMed]
- Hodek, I.; Evans, E.W. Food relationships, In Ecology and Behaviour of Ladybird Beetles (Coccinellidae); Hodek, I., van Emden, H.F., Honěk, A., Eds.; Blackwell Publishing Ltd.: Chichester, UK, 2012; Chapter 8; pp. 141–274. [Google Scholar]
- Schultz, H.; Silva, E.; Aguiar-Menezes, E.L.; Resende, A.L.S.; Rouws, J.R.C.; Silva, A.R.M. Adequacy of Drosophila melanogaster as prey for the development and reproduction of Coleomegilla maculata. BioControl 2019, 64, 43–54. [Google Scholar] [CrossRef]
- Smith, B.C. A technique for rearing coccinellid beetles on dry foods, and influence of various pollens on the development of Coleomegilla maculata lengi Timb. Can. J. Zool. 1960, 38, 1047–1049. [Google Scholar] [CrossRef]
- Lundgren, J.G.; Huber, A.; Wiedenmann, R.N. Quantification of consumption of corn pollen by the predator Coleomegilla maculata (Coleoptera: Coccinellidae) during anthesis in an Illinois cornfield. Agric. For. Entomol. 2005, 7, 53–60. [Google Scholar] [CrossRef]
- Chia, S.Y.; Tanga, C.M.; Osuga, I.M.; Cheseto, X.; Ekesi, S.; Dicke, M.; van Loon, J.J.A. Nutritional composition of black soldier fly larvae feeding on agro-industrial by-products. Entomol. Exp. Appl. 2020, 168, 472–481. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z.; Rojas, M.G. Potential utilization of Artemia franciscana eggs as food for Coleomegilla maculata. BioControl 2014, 59, 575–583. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z. Does a change from whole to powdered food (Artemia franciscana eggs) increase oviposition in the ladybird Coleomegilla maculata? Insects 2015, 6, 815–826. [Google Scholar] [CrossRef]
- Zar, J.H. The fatty acid composition of the ladybird beetle, Coleomegilla maculata (DeGeer) during hibernation. Comp. Biochem. Physiol. 1968, 26, 1127–1129. [Google Scholar] [CrossRef]
- Barlow, J.S. Fatty acids in some insect and spider fats. Can. J. Biochem. 1964, 42, 1365–1374. [Google Scholar] [CrossRef]
- Callow, R.K.; Greenway, A.R.; Griffiths, D.C. Chemistry of the secretion from the cornicles of various species of aphids. J. Insect Physiol. 1973, 19, 737–748. [Google Scholar] [CrossRef]
- Rojas, M.G.; Morales-Ramos, J.A.; Riddick, E.W. Use of Tenebrio molitor (Coleoptera: Tenebrionidae) powder to enhance artificial diet formulation for Coleomegilla maculata (Coleoptera: Coccinellidae). Biol. Control 2016, 100, 70–78. [Google Scholar] [CrossRef]
- Riddick, E.W.; Wu, Z.; Rojas, M.G. Is Tetranychus urticae suitable prey for development and reproduction of naïve Coleomegilla maculata? Insect Sci. 2014, 21, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Bashir, M.O. Effect of Nutrition on Development and Reproduction of Aphidophagous Coccinellids with Special Reference to Olla abdominalis (Say). Ph.D. Thesis, University of California, Berkeley, CA, USA, 1973. [Google Scholar]
- Hagen, K. Nutritional ecology of terrestrial insect predators. In Nutritional Ecology of Insects, Mites, Spiders, and Related Invertebrates; Slansky, F., Jr., Rodriguez, J.G., Eds.; Wiley & Sons: New York, NY, USA, 1987; Chapter 7; pp. 533–577. [Google Scholar]
- Naser El Deen, S.; Spranghers, T.; Baldacchino, F.; Deruytter, D. The effects of the particle size of four different feeds on the larval growth of Tenebrio molitor (Coleoptera: Tenebrionidae). Eur. J. Entomol. 2022, 119, 242–249. [Google Scholar] [CrossRef]
- Michaud, J.P.; Jyoti, J.L. Dietary complementation across life stages in the polyphagous lady beetle Coleomegilla maculata. Entomol. Exp. Appl. 2008, 126, 40–45. [Google Scholar] [CrossRef]
- Oonincx, D.G.A.B.; van Broekhoven, S.; van Huis, A.; van Loon, J.J.A. Feed conversion, survival and development, and composition of four insect species on diets composed of food by-products. PLoS ONE 2015, 10, e0144601. [Google Scholar] [CrossRef]
- St-Hilaire, S.; Sheppard, C.; Tomberlin, J.K.; Irving, S.; Newton, L.; McGuire, M.A.; Mosley, E.E.; Hardy, R.W.; Sealey, W. Fly prepupae as a feedstuff for rainbow trout, Oncorhynchus mykiss. J. World Aquacult. Soc. 2007, 38, 59–67. [Google Scholar] [CrossRef]
- Richardson, A.; Dantas-Lima, J.; Lefranc, M.; Walraven, M. Effect of black soldier fly ingredient on the growth performance and disease resistance of juvenile Pacific white shrimp (Litopenaeus vannamei). Animals 2021, 11, 1450. [Google Scholar] [CrossRef]
- Bommarco, R. Stage sensitivity to food limitation for a generalist arthropod predator, Pterostichus cupreus (Coleoptera: Carabidae). Environ. Entomol. 1998, 27, 863–869. [Google Scholar] [CrossRef]
- Jalali, M.A.; Tirry, L.; De Clercq, P. Food consumption and immature growth of Adalia bipunctata (Coleoptera: Coccinellidae) on a natural prey and a factitious food. Eur. J. Entomol. 2009, 106, 193–198. [Google Scholar] [CrossRef]
- Mercer, N.H.; Obrycki, J.J. Impacts of larval diet on pre-imaginal development, survival and adult size of six species of Coccinellidae (Coleoptera). J. Kansas Entomol. Soc. 2020, 93, 256–261. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| 1 Diet Treatments | First Instars per Arena | Days to Pupal Stage | Pupae | Adults | Adult Body Mass (mg) |
|---|---|---|---|---|---|
| BSE+CM | 10 | 13.08 ± 0.14 b | 8.80 ± 0.39 a | 8.70 ± 0.37 a | 15.29 ± 0.37 a |
| BSF | 10 | 17.48 ± 0.17 a | 5.50 ± 0.43 b | 5.20 ± 0.47 b | 9.24 ± 0.23 c |
| BSF+CM | 10 | 17.87 ± 0.18 a | 6.80 ± 0.55 b | 6.30 ± 0.58 b | 10.16 ± 0.20 b |
| F | -- | 267.63 | 12.93 | 13.99 | 139.78 |
| df | -- | 2, 27 | 2, 27 | 2, 27 | 2, 27 |
| p | -- | <0.001 | <0.001 | <0.001 | <0.001 |
| 1 Diet Treatments | N, Females | Emergence to Mating (Days) | Mating to First Egg Clutch (Days) | Clutches per Female | Eggs per Clutch |
|---|---|---|---|---|---|
| BSE+CM | 13 | 8.38 ± 0.42 b | 8.93 ± 1.71 a | 3.69 ± 0.79 a | 14.80 ± 2.22 a |
| BSF | 7 | 14.0 ± 1.46 a | 13.43 ± 3.12 a | 4.00 ± 1.33 a | 6.35 ± 0.95 b |
| BSF+CM | 8 | 11.0 ± 0.96 ab | 12.50 ± 3.66 a | 4.37 ± 1.05 a | 9.62 ± 1.05 ab |
| F | -- | 10.79 | 0.89 | 0.12 | 5.09 |
| df | -- | 2, 25 | 2, 25 | 2, 25 | 2, 25 |
| p | -- | <0.001 | 0.42 | 0.88 | 0.014 |
| 1 Diet Treatments | First Instars per Arena | Days to Pupal Stage | Pupae | Adults | Adult Body Mass (mg) |
|---|---|---|---|---|---|
| BSE+CM | 10 | 14.01 ± 0.27 b | 9.00 ± 0.33 a | 8.60 ± 0.40 a | 14.59 ± 0.32 a |
| BSF+AD | 10 | 17.23 ± 0.23 a | 6.56 ± 0.44 b | 6.22 ± 0.55 b | 11.17 ± 0.40 b |
| t | -- | 8.91 | 4.46 | 3.56 | 6.71 |
| df | -- | 17 | 17 | 17 | 17 |
| p | -- | <0.001 | <0.001 | 0.002 | <0.001 |
| 1 Diet Treatments | N, Females | Emergence to Mating (Days) | Mating to First Egg Clutch (Days) | Clutches per Female | Eggs per Clutch |
|---|---|---|---|---|---|
| BSE+CM | 12 | 6.33 ± 0.61 b | 8.58 ± 2.12 a | 4.50 ± 0.73 a | 11.05 ± 1.73 a |
| BSF+AD | 9 | 11.33 ± 1.44 a | 13.00 ± 2.07 a | 6.22 ± 1.28 a | 11.23 ± 1.40 a |
| t | -- | 3.51 | 1.45 | 1.24 | 0.08 |
| df | -- | 19 | 19 | 19 | 19 |
| p | -- | 0.002 | 0.16 | 0.23 | 0.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riddick, E.W.; Walker, R.C.; Rojas, M.G.; Morales-Ramos, J.A. Evaluation of Black Soldier Fly Hermetia illucens as Food for Pink-Spotted Lady Beetle Coleomegilla maculata. Insects 2023, 14, 902. https://doi.org/10.3390/insects14120902
Riddick EW, Walker RC, Rojas MG, Morales-Ramos JA. Evaluation of Black Soldier Fly Hermetia illucens as Food for Pink-Spotted Lady Beetle Coleomegilla maculata. Insects. 2023; 14(12):902. https://doi.org/10.3390/insects14120902
Chicago/Turabian StyleRiddick, Eric W., Ryan C. Walker, Maria Guadalupe Rojas, and Juan A. Morales-Ramos. 2023. "Evaluation of Black Soldier Fly Hermetia illucens as Food for Pink-Spotted Lady Beetle Coleomegilla maculata" Insects 14, no. 12: 902. https://doi.org/10.3390/insects14120902