
Earwig Releases Provide Accumulative Biological Control of the Woolly Apple Aphid over the Years
 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Treatments
2.3. Assessments
2.4. Statistical Analysis
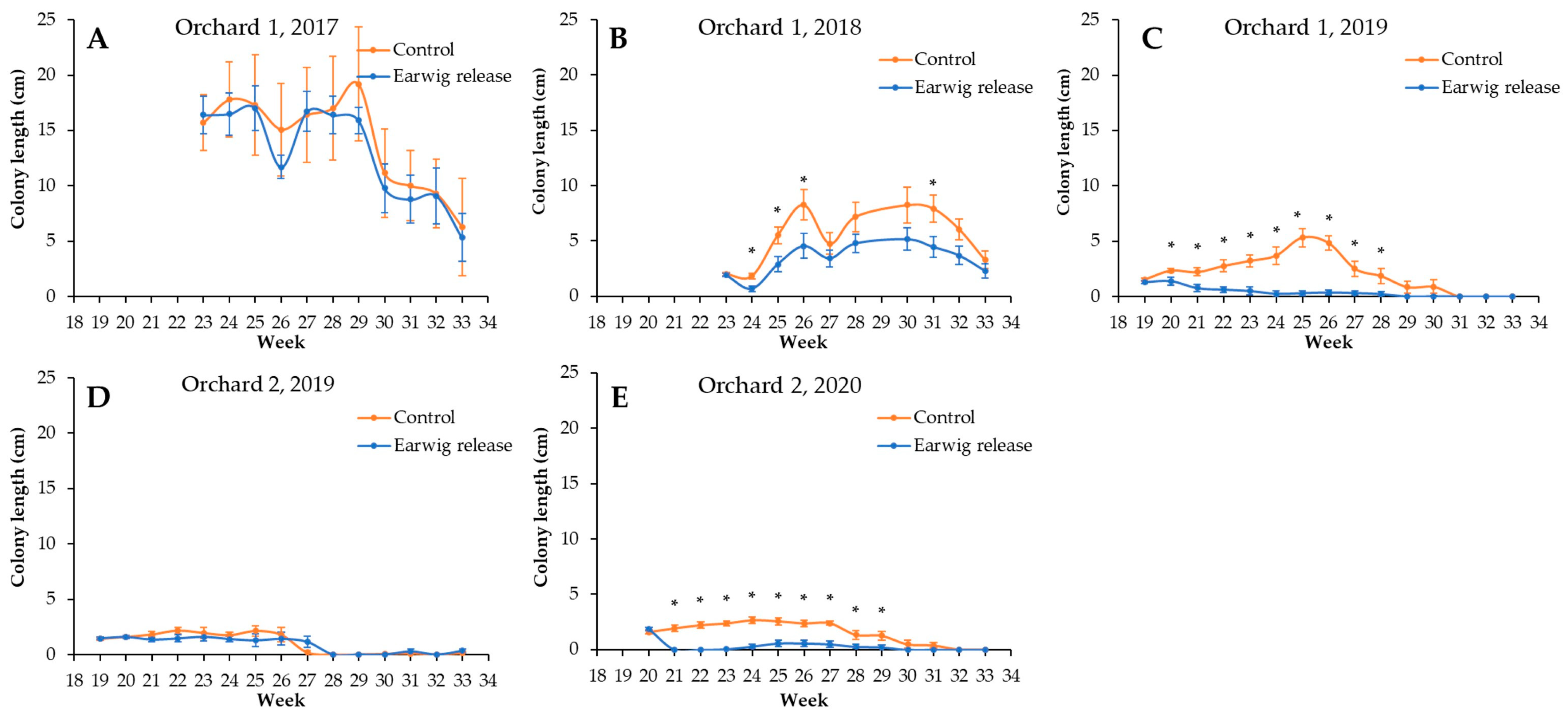
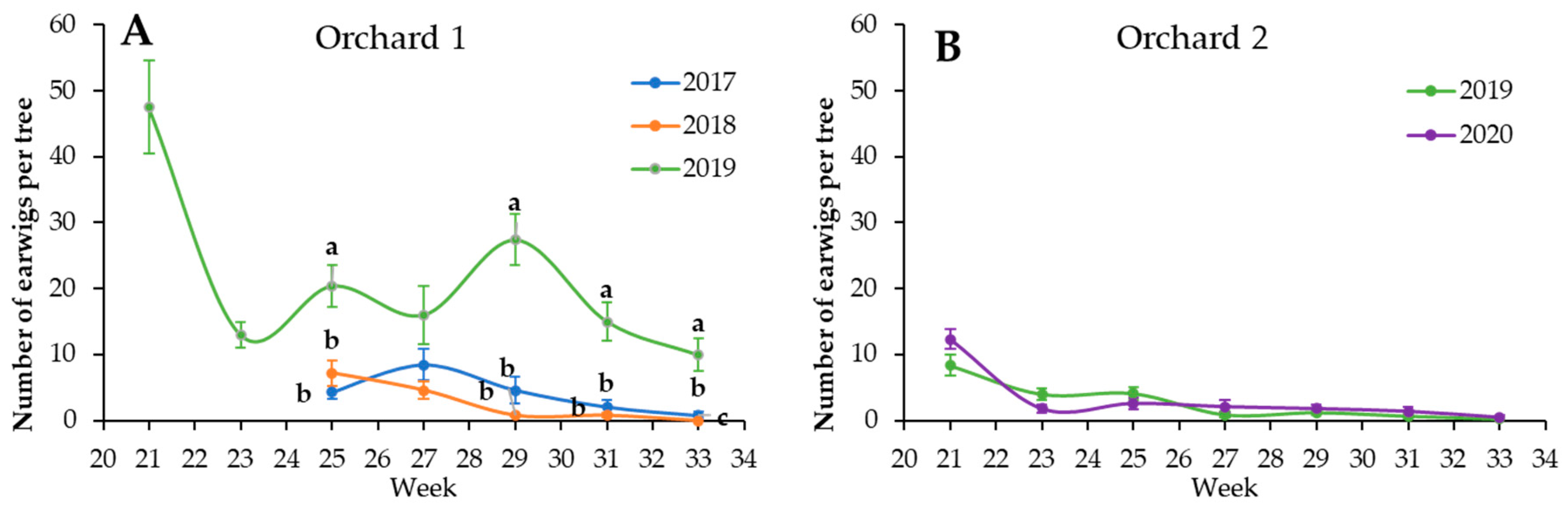
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Cock, M.J.W.; van Lenteren, J.C.; Brodeur, J.; Barratt, B.I.P.; Bigler, F.; Bolckmans, K.; Cônsoli, F.L.; Haas, F.; Mason, P.G.; Parra, J.R.P. Do new Access and Benefit Sharing procedures under the Convention on Biological Diversity threaten the future of biological control? BioControl 2010, 55, 199–218. [Google Scholar] [CrossRef]
- Brodeur, J.; Abram, P.K.; Heimpel, G.E.; Messing, R.H. Trends in biological control: Public interest, international networking and research direction. BioControl 2018, 63, 11–26. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological control using invertebrates and microorganisms: Plenty of new opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef]
- Eilenberg, J.; Hajek, A.; Lomer, C. Suggestions for unifying the terminology in biological control. BioControl 2001, 46, 387–400. [Google Scholar] [CrossRef]
- Lavandero, B.; Ortiz-Martínez, S.; Peñalver-Cruz, A.; Zepeda-Paulo, F. Bases Ecológicas del Control Biológico Derivadas del Control Natural: Factores de Importancia Para el Manejo de Plagas. In Bases Ecológicas Para el Manejo de Plagas; Estay, S.A., Ed.; Ediciones Universidad Católica de Chile: Santiago, Chile, 2021; p. 263. [Google Scholar]
- Heimpel, G.; Mills, N. Biological Control: Ecology and Applications; Cambridge University Press: Cambridge, UK, 2017; p. 380. [Google Scholar]
- Bostanian, N.J.; Goulet, H.; O’Hara, J.; Masner, L.; Racette, G. Towards Insecticide Free Apple Orchards: Flowering Plants to Attract Beneficial Arthropods. Biocontrol Sci. Technol. 2004, 14, 25–37. [Google Scholar] [CrossRef]
- Cahenzli, F.; Sigsgaard, L.; Daniel, C.; Herz, A.; Jamar, L.; Kelderer, M.; Jacobsen, S.K.; Kruczynska, D.; Matray, S.; Porcel, M.; et al. Perennial flower strips for pest control in organic apple orchards—A pan-European study. Agric. Ecosyst. Environ. 2019, 278, 43–53. [Google Scholar] [CrossRef]
- Herz, A.; Cahenzli, F.; Penvern, S.; Pfiffner, L.; Tasin, M.; Sigsgaard, L. Managing Floral Resources in Apple Orchards for Pest Control: Ideas, Experiences and Future Directions. Insects 2019, 10, 247. [Google Scholar] [CrossRef]
- Dedryver, C.-A.; Le Ralec, A.; Fabre, F. The conflicting relationships between aphids and men: A review of aphid damage and control strategies. Comptes Rendus Biol. 2010, 333, 539–553. [Google Scholar] [CrossRef]
- Penvern, S.; Bellon, S.; Fauriel, J.; Sauphanor, B. Peach orchard protection strategies and aphid communities: Towards an integrated agroecosystem approach. Crop Prot. 2010, 29, 1148–1156. [Google Scholar] [CrossRef]
- Barbagallo, S.; Cocuzza, G.E.M.; Cravedi, P.; Komazaki, S. IPM Case Studies: Deciduous Fruit Tree Aphids. In Aphids as Crop Pests, 2nd ed.; VanEmden, H.F., Harrington, R., Eds.; CAB International: Wallingford, UK, 2017; pp. 632–642. [Google Scholar]
- Andrews, K.B.; Powell, K.S. Understanding the Fundamental Interactions between Woolly Apple Aphid and Pome Fruit; Horticulture Australia Ltd.: Sydney, Australia, 2009; p. 65. [Google Scholar]
- Short, B.D.; Bergh, J.C. Feeding and egg distribution studies of Heringia calcarata (Diptera: Syrphidae), a specialized predator of woolly apple aphid (Homoptera: Eriosomatidae) in Virginia apple orchards. J. Econ. Entomol. 2004, 97, 813–819. [Google Scholar] [CrossRef]
- Asante, S.K. Functional Responses of the European Earwig and Two Species of Coccinellids to Densities of Eriosoma lanigerum (Hausmann) (Hemiptera: Aphididae). Aust. J. Entomol. 1995, 34, 105–109. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Cockfield, S.D.; Beers, E.H. Natural Enemies of Woolly Apple Aphid (Hemiptera: Aphididae) in Washington State. Environ. Entomol. 2012, 41, 1364–1371. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.B.; Souza, A.; Belli, E.L. Parasitism on Eriosoma lanigerum (Homoptera: Aphididae) by Aphelinus mali (Hymenoptera: Encyrtidae) on apple orchards, in Fraiburgo county, state of Santa Catarina, Brazil. Rev. Bras. Frutic. 2004, 26, 550–551. [Google Scholar] [CrossRef]
- Quarrell, S.R.; Corkrey, R.; Allen, G.R. Predictive thresholds for forecasting the compatibility of Forficula auricularia and Aphelinus mali as biological control agents against woolly apple aphid in apple orchards. Biocontrol 2017, 62, 243–256. [Google Scholar] [CrossRef]
- Mueller, T.F.; Blommers, L.H.M.; Mols, P.J.M. Earwig (Forficula auricularia) predation on the woolly apple aphid, Eriosoma lanigerum. Entomol. Exp. Appl. 1988, 47, 145–152. [Google Scholar] [CrossRef]
- Nicholas, A.H.; Spooner-Hart, R.N.; Vickers, R.A. Abundance and natural control of the woolly aphid Eriosoma lanigerum in an Australian apple orchard IPM program. BioControl 2005, 50, 271–291. [Google Scholar] [CrossRef]
- Lordan, J.; Alegre, S.; Gatius, F.; Sarasúa, M.J.; Alins, G. Woolly apple aphid Eriosoma lanigerum Hausmann ecology and its relationship with climatic variables and natural enemies in Mediterranean areas. Bull. Entomol. Res. 2015, 105, 60–69. [Google Scholar] [CrossRef]
- Jauset, A.M.; Artigues, M.; Avilla, J.; Sarasúa, M.J. Preliminary study of Forficula sp. (Dermaptera:Forficulidae) as predator of Cacopsylla pyri (Homoptera: Psyllidae), under field conditions in Lleida (Spain). IOBC/WPRS Bull. 2005, 28, 125–128. [Google Scholar]
- Logan, D.P.; Maher, B.J.; Rowe, C.A. Predation of diaspidid scale insects on kiwifruit vines by European earwigs, Forficula auricularia, and steel-blue ladybirds, Halmus chalybeus. Biocontrol 2017, 62, 469–479. [Google Scholar] [CrossRef]
- Englert, C.; Herz, A. Acceptability of Drosophila suzukii as prey for common predators occurring in cherries and berries. J. Appl. Entomol. 2019, 143, 387–396. [Google Scholar] [CrossRef]
- Gabarra, R.; Riudavets, J.; Rodríguez, G.A.; Pujade-Villar, J.; Arnó, J. Prospects for the biological control of Drosophila suzukii. Biocontrol 2015, 60, 331–339. [Google Scholar] [CrossRef]
- Weiss, M.J.; McDonald, G. European earwig, Forficula auricularia L. (Dermaptera: Forficulidae), as a predator of the redlegged earth mite, Halotydeus destructor (Tucker) (Acarina: Penthaleidae). Aust. J. Entomol. 1998, 37, 183–185. [Google Scholar] [CrossRef]
- Carroll, D.P.; Hoyt, S.C. Augmentation of European earwigs (Dermaptera, Forficulidae) for biological control of apple aphid (Homoptera, Aphididae) in an apple orchard. J. Econ. Entomol. 1984, 77, 738–740. [Google Scholar] [CrossRef]
- Piñol, J.; Espadaler, X.; Cañellas, N.; Pérez, N. Effects of the concurrent exclusion of ants and earwigs on aphid abundance in an organic citrus grove. BioControl 2009, 54, 515–527. [Google Scholar] [CrossRef]
- Carroll, D.P.; Walker, J.T.S.; Hoyt, S.C. European earwigs (Dermaptera, Forficulidae) fail to control apple aphids on bearing apple-trees and woolly apple aphids (Homoptera, Aphididae) in apple rootstock stool beds. J. Econ. Entomol. 1985, 78, 972–974. [Google Scholar] [CrossRef]
- Romeu-Dalmau, C.; Piñol, J.; Agustí, N. Detecting aphid predation by earwigs in organic citrus orchards using molecular markers. Bull. Entomol. Res. 2012, 102, 566–572. [Google Scholar] [CrossRef]
- Dib, H.; Jamont, M.; Sauphanor, B.; Capowiez, Y. Predation potency and intraguild interactions between generalist (Forficula auricularia) and specialist (Episyrphus balteatus) predators of the rosy apple aphid (Dysaphis plantaginea). Biol. Control 2011, 59, 90–97. [Google Scholar] [CrossRef]
- Kirstová, M.; Pyszko, P.; Kočárek, P. Factors influencing microhabitat selection and food preference of tree-dwelling earwigs (Dermaptera) in a temperate floodplain forest. Bull. Entomol. Res. 2019, 109, 54–61. [Google Scholar] [CrossRef]
- Crumb, S.; Eide, P.; Bonn, A. The European Earwig. Tech. Bull. USA Dep. Agric. Wash. 1941, 766, 1–76. [Google Scholar]
- Dib, H.; Jamont, M.; Sauphanor, B.; Capowiez, Y. The feasibility and efficacy of early-season releases of a generalist predator (Forficula auricularia L.) to control populations of the RAA (Dysaphis plantaginea Passerini) in Southeastern France. Bull. Entomol. Res. 2016, 106, 233–241. [Google Scholar] [CrossRef]
- Orpet, R.J.; Goldberger, J.R.; Crowder, D.W.; Jones, V.P. Field evidence and grower perceptions on the roles of an omnivore, European earwig, in apple orchards. Biol. Control 2019, 132, 189–198. [Google Scholar] [CrossRef]
- Lordan, J.; Alegre, S.; Moerkens, R.; Sarasúa, M.J.; Alins, G. Phenology and interspecific association of Forficula auricularia and Forficula pubescens in apple orchards. Span. J. Agric. Res. 2015, 13, e1003. [Google Scholar] [CrossRef]
- Moerkens, R.; Leirs, H.; Peusens, G.; Gobin, B. Dispersal of single- and double-brood populations of the European earwig, Forficula auricularia: A mark-recapture experiment. Entomol. Exp. Appl. 2010, 137, 19–27. [Google Scholar] [CrossRef]
- Lamb, R.J.; Wellington, W.G. Life-history and population characteristics of European earwig, Forficula auricularia (Dermaptera: Forficulidae), at Vancouver, British-Columbia. Can. Entomol. 1975, 107, 819–824. [Google Scholar] [CrossRef]
- Moerkens, R.; Leirs, H.; Peusens, G.; Gobin, B. Are populations of European earwigs, Forficula auricularia, density dependent? Entomol. Exp. Appl. 2009, 130, 198–206. [Google Scholar] [CrossRef]
- Shaw, B.; Nagy, C.; Fountain, M.T. Organic Control Strategies for Use in IPM of Invertebrate Pests in Apple and Pear Orchards. Insects 2021, 12, 1106. [Google Scholar] [CrossRef]
- Suckling, D.M.; Burnip, G.M.; Hackett, J.; Daly, J.C. Frass sampling and baiting indicate European earwig (Forficula auricularia) foraging in orchards. J. Appl. Entomol. 2006, 130, 263–267. [Google Scholar] [CrossRef]
- Orpet, R.J.; Crowder, D.W.; Jones, V.P. Woolly Apple Aphid Generalist Predator Feeding Behavior Assessed through Video Observation in an Apple Orchard. J. Insect Behav. 2019, 32, 153–163. [Google Scholar] [CrossRef]
- Burnip, G.M.; Daly, M.J.; Hackett, J.K.; Suckling, D.M. European earwig phenology and effect of understorey management on population estimation. N. Z. Plant Prot. 2002, 55, 390–395. [Google Scholar] [CrossRef]
- Happe, A.-K.; Roquer-Beni, L.; Bosch, J.; Alins, G.; Mody, K. Earwigs and woolly apple aphids in integrated and organic apple orchards: Responses of a generalist predator and a pest prey to local and landscape factors. Agric. Ecosyst. Environ. 2018, 268, 44–51. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Beers, E.H.; Snyder, W.E. Complementary suppression of aphids by predators and parasitoids. Biol. Control 2015, 90, 83–91. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orchard | Year | Control | Earwig Release | DF | F | p |
|---|---|---|---|---|---|---|
| Orchard 1 | 2017 | 942.5 ± 259.8 | 1091.2 ± 115.6 | 1,15 | 0.3211 | 0.5799 |
| Orchard 1 | 2018 | 381.8 ± 52.6 | 231.7 ± 41.5 | 1,19 | 5.0138 | 0.0380 |
| Orchard 1 | 2019 | 276.2 ± 36.7 | 52.9 ± 15.3 | 1,19 | 31.544 | <0.0001 |
| Orchard 2 | 2019 | 126.1 ± 21.6 | 111.4 ± 21.2 | 1,18 | 0.2345 | 0.6344 |
| Orchard 2 | 2020 | 170.4 ± 16.6 | 41.4 ± 12.1 | 1,18 | 40.4831 | <0.0001 |
| Orchard | Year | Control | Earwig Release | DF | F/χ2 | p |
|---|---|---|---|---|---|---|
| Orchard 1 | 2017 | 1165.5 ± 312.1 | 1285.2 ± 294.8 | 1,19 | 0.0777 | 0.7836 |
| Orchard 1 | 2018 | 4555.6 ± 273.1 | 4122.3 ± 119.6 | 1,19 | 2.1124 | 0.1633 |
| Orchard 1 | 2019 | 14,515.2 ± 1672.4 | 20,692.7 ± 2924.9 | 1 | 2.0629 | 0.1509 |
| Orchard 2 | 2019 | 21,261.3 ± 1503.1 | 22,638 ± 1797.6 | 1,18 | 0.3365 | 0.5695 |
| Orchard 2 | 2020 | 7700 ± 725 | 7736.1 ± 648.3 | 1,19 | 0.0014 | 0.9708 |
| Orchard | Year | Week | Control | Earwig Release | DF | F/χ2 | p |
|---|---|---|---|---|---|---|---|
| Orchard 1 | 2017 | 23 | 24.0 ± 6.4 | 22.5 ± 3.8 | 1,19 | 0.0401 | 0.8436 |
| 24 | 23.0 ± 8.5 | 28.0 ± 7.9 | 1,19 | 0.1856 | 0.6717 | ||
| 25 | 15.0 ± 4.1 | 23.0 ± 8.5 | 1 | 0.1206 | 0.7283 | ||
| 26 | 35.5 ± 8.7 | 62.0 ± 8.0 | 1,19 | 5.0230 | 0.0379 | ||
| 27 | 34.5 ± 10.4 | 40.0 ± 10.3 | 1,19 | 0.1416 | 0.7111 | ||
| 28 | 32 ± 12.4 | 32.0 ± 12.4 | 1,19 | 0.0000 | 1.0000 | ||
| 29 | 30.6 ± 8.4 | 33.0 ± 11.0 | 1,18 | 0.0302 | 0.8641 | ||
| 30 | 65.0 ± 12.0 | 83.3 ± 7.7 | 1,14 | 1.8291 | 0.1993 | ||
| 31 | 78.3 ± 5.3 | 84.3 ± 5.1 | 1,12 | 0.6611 | 0.4334 | ||
| 32 | 91.7 ± 4.4 | 95.7 ± 0.7 | 1,12 | 0.9627 | 0.3476 | ||
| 33 | 98.0 ± 1.2 | 96.7 ± 1.1 | 1,10 | 0.6890 | 0.4280 | ||
| 2018 | 23 | 0.0 ± 0.0 | 0.0 ± 0.0 | - | - | - | |
| 24 | 9.5 ± 6.8 | 11.7 ± 5.9 | 1,15 | 0.0478 | 0.8301 | ||
| 25 | 24.0 ± 6.4 | 15.5 ± 4.9 | 1,19 | 1.1092 | 0.3062 | ||
| 26 | 21.5 ± 6.7 | 7.5 ± 3.8 | 1,19 | 3.3189 | 3.3189 | ||
| 27 | 43.5 ± 7.6 | 30.5 ± 9.5 | 1,19 | 1.1415 | 0.2995 | ||
| 28 | 32.5 ± 7.2 | 21.5 ± 6.7 | 1,19 | 1.2568 | 0.2770 | ||
| 30 | 59.0 ± 8.5 | 37.0 ± 9.9 | 1,19 | 2.8564 | 0.1083 | ||
| 31 | 70.5 ± 9.4 | 77.0 ± 8.5 | 1,19 | 0.2644 | 0.6134 | ||
| 32 | 95.0 ± 0.0 | 92.2 ± 2.8 | 1 | 1.1111 | 0.2918 | ||
| 33 | 95.0 ± 0.0 | 91.9 ± 3.1 | 1 | 1.0000 | 0.3173 | ||
| 2019 | 19 | 6.5 ± 2.0 | 2.5 ± 1.1 | 1,19 | 3.0968 | 0.0954 | |
| 20 | 23.5 ± 5.1 | 28.6 ± 9.3 | 1,16 | 0.2653 | 0.6140 | ||
| 21 | 38.0 ± 10.7 | 55.7 ± 16.2 | 1,16 | 0.9065 | 0.3561 | ||
| 22 | 13.9 ± 4.1 | 17.5 ± 14.2 | 1,12 | 0.1084 | 0.7481 | ||
| 23 | 20.0 ± 5.1 | 7.5 ± 2.5 | 1,10 | 1.2355 | 0.2951 | ||
| 24 | 13.5 ± 5.0 | 51.3 ± 48.8 | 1 | 0.1886 | 0.6641 | ||
| 25 | 15.5 ± 4.2 | 28.3 ± 16.4 | 1,12 | 1.3109 | 0.2765 | ||
| 26 | 14.0 ± 3.2 | 51.3 ± 19.4 | 1,13 | 8.7151 | 0.0121 | ||
| 27 | 54.0 ± 11.5 | 47.5 ± 4.8 | 1 | 0.1849 | 0.6672 | ||
| 28 | 68.6 ± 12.4 | 75.0 ± 25.0 | 1,8 | 0.0582 | 0.8163 | ||
| 29 | 66.0 ± 14.7 | 100.0 | 1,5 | 0.8920 | 0.3984 | ||
| 30 | 56.7 ± 21.9 | 50.0 | 1,3 | 0.0233 | 0.8928 | ||
| 31 | 100.0 ± 0.0 | 100.0 | - | - | - | ||
| Orchard 2 | 2019 | 19 | 10.5 ± 2.5 | 13.3 ± 5.2 | 1,18 | 0.2563 | 0.6192 |
| 20 | 28.0 ± 4.6 | 28.3 ± 7.9 | 1,18 | 0.0014 | 0.9707 | ||
| 21 | 17.3 ± 4.3 | 22.2 ± 5.3 | 1,18 | 0.5401 | 0.4724 | ||
| 22 | 25.5 ± 5.3 | 19.3 ± 4.8 | 1,16 | 0.6730 | 0.4248 | ||
| 23 | 31.0 ± 5.8 | 28.6 ± 6.6 | 1,16 | 0.0755 | 0.7873 | ||
| 24 | 49.0 ± 4.3 | 30.6 ± 8.3 | 1 | 2.7056 | 0.1000 | ||
| 25 | 45.0 ± 7.9 | 61.7 ± 11.7 | 1,15 | 1.4983 | 0.2411 | ||
| 26 | 79.5 ± 6.3 | 66.4 ± 10.6 | 1,16 | 1.2857 | 0.2746 | ||
| 27 | 97.9 ± 1.5 | 81.0 ± 6.4 | 1,11 | 9.0793 | 0.0130 | ||
| 28 | 95.0 | 98.8 ± 1.3 | 1,4 | 1.8000 | 0.2722 | ||
| 29 | - | 100.0 | - | - | - | ||
| 30 | 99.0 | - | - | - | |||
| 31 | 95.0 ± 5.0 | 82.5 ± 2.5 | 1,3 | 5.0000 | 0.1548 | ||
| 32 | 95.0 | 100.0 ± 0.0 | - | - | - | ||
| 33 | 30.0 | 75.0 ± 7.6 | 1,3 | 8.6786 | 0.0985 | ||
| 2020 | 20 | 15.0 ± 2.1 | 17.5 ± 2.4 | 1,19 | 0.6164 | 0.4426 | |
| 21 | 29.0 ± 5.5 | 100.0 ± 0.0 | 1 | 15.2226 | <0.0001 | ||
| 22 | 42.5 ± 5.5 | 100.0 | 1,10 | 9.9728 | 0.0116 | ||
| 23 | 43.3 ± 7.3 | 50.0 ± 50.0 | 1,1.04 | 0.0174 | 0.1319 | ||
| 24 | 45.6 ± 6.3 | 37.5 ± 22.5 | 1,3.4833 | 0.1188 | 0.3447 | ||
| 25 | 35.0 ± 7.0 | 52.5 ± 19.3 | 1,12 | 1.1698 | 0.3026 | ||
| 26 | 53.3 ± 8.0 | 91.7 ± 1.7 | 1,11 | 7.1616 | 0.0232 | ||
| 27 | 69.4 ± 6.8 | 85 ± 12.6 | 1,11 | 1.2744 | 0.2853 | ||
| 28 | 76.8 ± 7.5 | 65.0 ± 25.0 | 1,9 | 0.3988 | 0.5453 | ||
| 29 | 87.5 ± 5.3 | 50.0 | 1,6 | 7.1962 | 0.0437 | ||
| 30 | 93.0 ± 5.8 | 100.0 | 1,5 | 0.2402 | 0.6497 | ||
| 31 | 95.0 ± 0.0 | - | - | - | - | ||
| 32 | 100.0 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alins, G.; Lordan, J.; Rodríguez-Gasol, N.; Arnó, J.; Peñalver-Cruz, A. Earwig Releases Provide Accumulative Biological Control of the Woolly Apple Aphid over the Years. Insects 2023, 14, 890. https://doi.org/10.3390/insects14110890
Alins G, Lordan J, Rodríguez-Gasol N, Arnó J, Peñalver-Cruz A. Earwig Releases Provide Accumulative Biological Control of the Woolly Apple Aphid over the Years. Insects. 2023; 14(11):890. https://doi.org/10.3390/insects14110890
Chicago/Turabian StyleAlins, Georgina, Jaume Lordan, Neus Rodríguez-Gasol, Judit Arnó, and Ainara Peñalver-Cruz. 2023. "Earwig Releases Provide Accumulative Biological Control of the Woolly Apple Aphid over the Years" Insects 14, no. 11: 890. https://doi.org/10.3390/insects14110890