

Insect-Mediated Pollination of Strawberries in an Urban Environment

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Biological Material
2.3. Experimental Design
2.4. Pollinator Observations and Pollination Treatments
2.5. Fruit Set and Fruit Quality
2.6. Data Analysis
3. Results
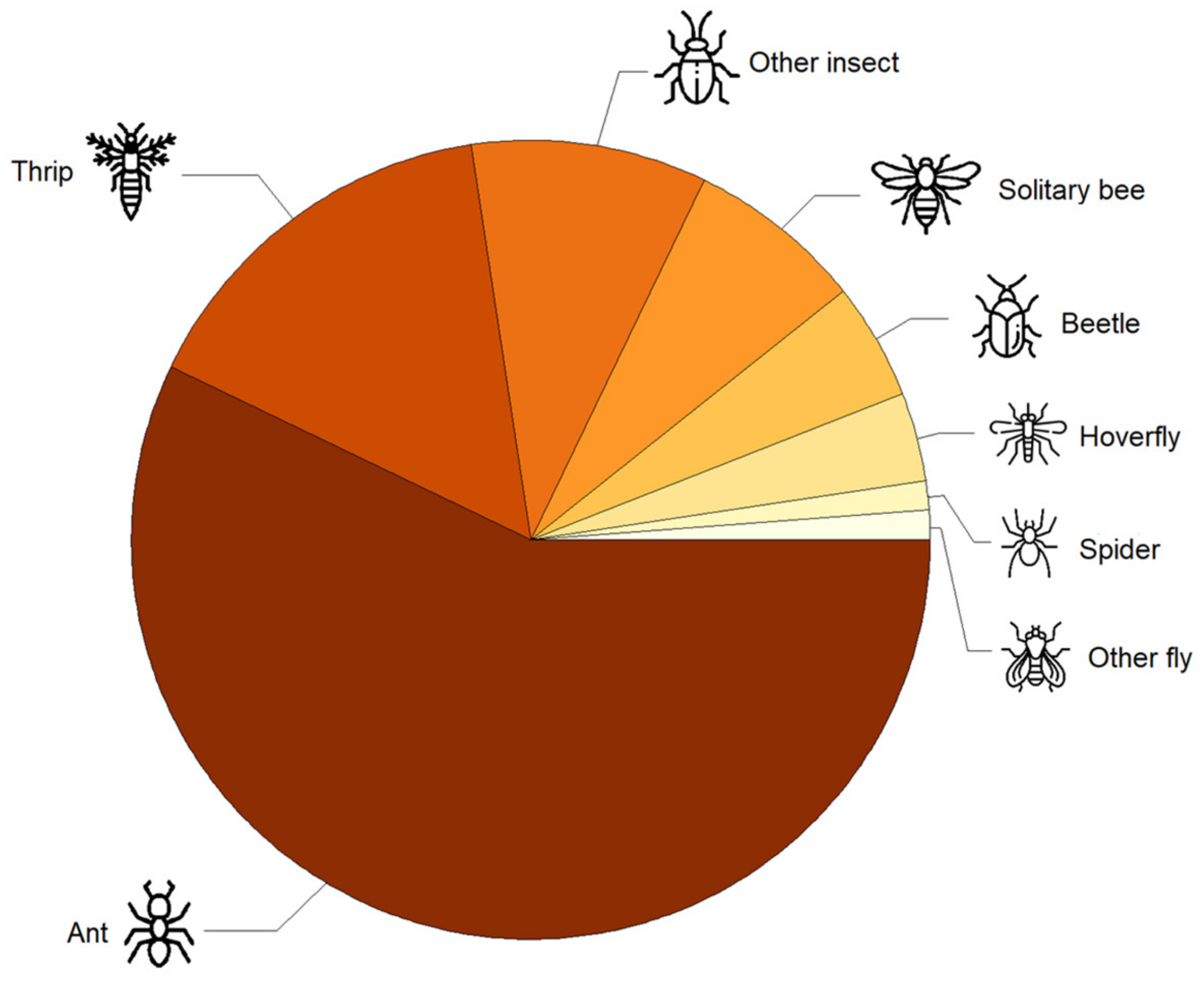
3.1. Flower Visitors
3.2. Fruit Set
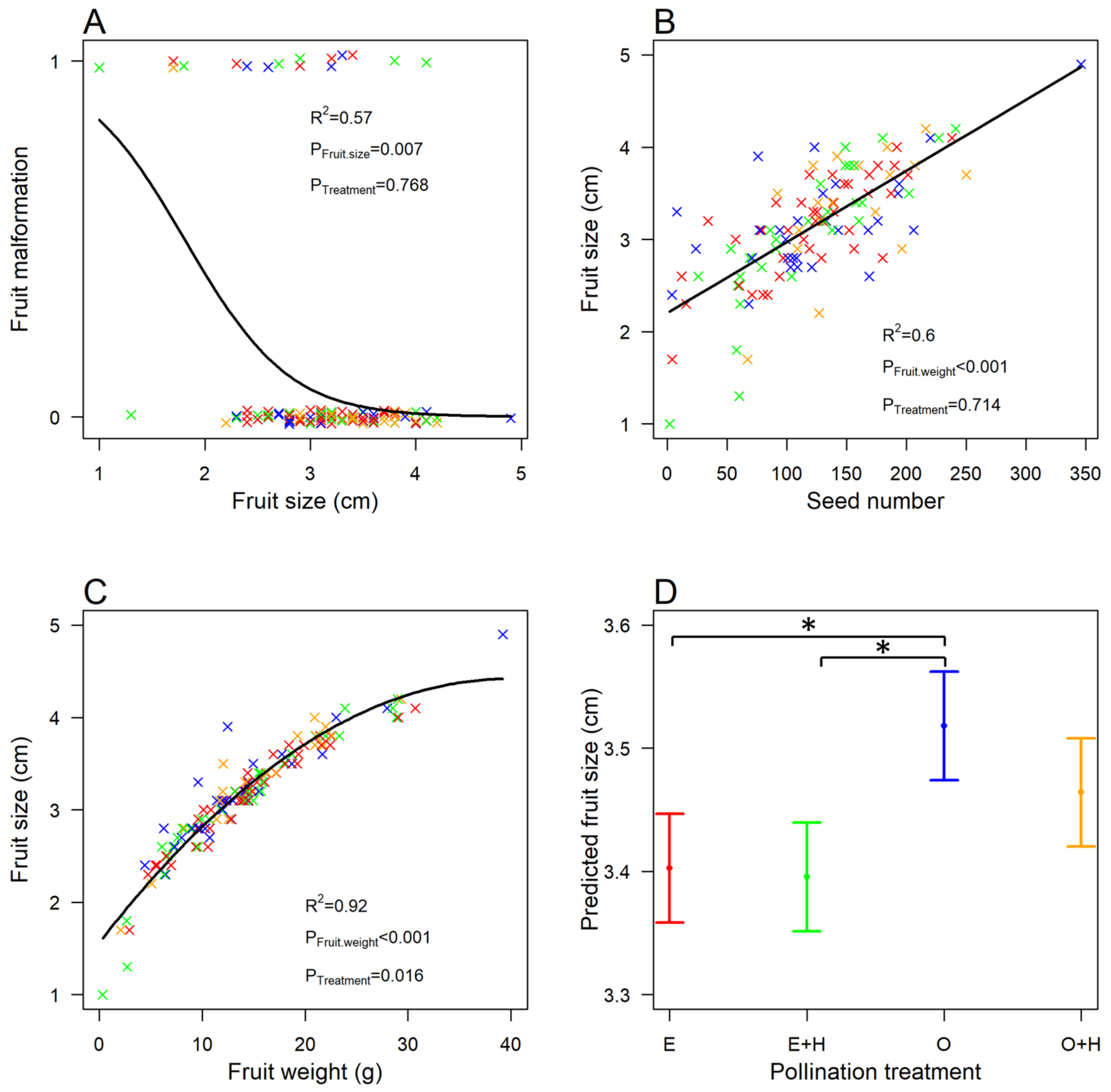
3.3. Fruit Malformation
3.4. Fruit Size
4. Discussion
5. Conclusion and Perspectives
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants Are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Klein, A.-M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of Pollinators in Changing Landscapes for World Crops. Proc. R. Soc. B 2007, 274, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Porto, R.G.; de Almeida, R.F.; Cruz-Neto, O.; Tabarelli, M.; Viana, B.F.; Peres, C.A.; Lopes, A.V. Pollination Ecosystem Services: A Comprehensive Review of Economic Values, Research Funding and Policy Actions. Food Sec. 2020, 12, 1425–1442. [Google Scholar] [CrossRef]
- Pérez-Méndez, N.; Andersson, G.K.S.; Requier, F.; Hipólito, J.; Aizen, M.A.; Morales, C.L.; García, N.; Gennari, G.P.; Garibaldi, L.A. The Economic Cost of Losing Native Pollinator Species for Orchard Production. J. Appl. Ecol. 2020, 57, 599–608. [Google Scholar] [CrossRef]
- Pisanty, G.; Afik, O.; Wajnberg, E.; Mandelik, Y. Watermelon Pollinators Exhibit Complementarity in Both Visitation Rate and Single-Visit Pollination Efficiency. J. Appl. Ecol. 2016, 53, 360–370. [Google Scholar] [CrossRef]
- Sapir, G.; Baras, Z.; Azmon, G.; Goldway, M.; Shafir, S.; Allouche, A.; Stern, E.; Stern, R.A. Synergistic Effects between Bumblebees and Honey Bees in Apple Orchards Increase Cross Pollination, Seed Number and Fruit Size. Sci. Hortic. 2017, 219, 107–117. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Requier, F.; Rollin, O.; Andersson, G.K. Towards an Integrated Species and Habitat Management of Crop Pollination. Curr. Opin. Insect Sci. 2017, 21, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Page, M.L.; Nicholson, C.C.; Brennan, R.M.; Britzman, A.T.; Greer, J.; Hemberger, J.; Kahl, H.; Müller, U.; Peng, Y.; Rosenberger, N.M.; et al. A Meta-analysis of Single Visit Pollination Effectiveness Comparing Honeybees and Other Floral Visitors. Am. J. Bot. 2021, 108, 2196–2207. [Google Scholar] [CrossRef]
- Rader, R.; Bartomeus, I.; Garibaldi, L.A.; Garratt, M.P.D.; Howlett, B.G.; Winfree, R.; Cunningham, S.A.; Mayfield, M.M.; Arthur, A.D.; Andersson, G.K.S.; et al. Non-Bee Insects Are Important Contributors to Global Crop Pollination. Proc. Natl. Acad. Sci. USA 2016, 113, 146–151. [Google Scholar] [CrossRef]
- Dicks, L.V.; Breeze, T.D.; Ngo, H.T.; Senapathi, D.; An, J.; Aizen, M.A.; Basu, P.; Buchori, D.; Galetto, L.; Garibaldi, L.A.; et al. A Global-Scale Expert Assessment of Drivers and Risks Associated with Pollinator Decline. Nat. Ecol. Evol. 2021, 5, 1453–1461. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef] [PubMed]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Ayers, A.C.; Rehan, S.M. Supporting Bees in Cities: How Bees Are Influenced by Local and Landscape Features. Insects 2021, 12, 128. [Google Scholar] [CrossRef] [PubMed]
- Banaszak-Cibicka, W.; Żmihorski, M. Are Cities Hotspots for Bees? Local and Regional Diversity Patterns Lead to Different Conclusions. Urban Ecosyst. 2020, 23, 713–722. [Google Scholar] [CrossRef]
- Fauviau, A.; Baude, M.; Bazin, N.; Fiordaliso, W.; Fisogni, A.; Fortel, L.; Garrigue, J.; Geslin, B.; Goulnik, J.; Guilbaud, L.; et al. A Large-Scale Dataset Reveals Taxonomic and Functional Specificities of Wild Bee Communities in Urban Habitats of Western Europe. Sci. Rep. 2022, 12, 18866. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, A.; Grass, I.; Belavadi, V.V.; Tscharntke, T. How Urbanization Is Driving Pollinator Diversity and Pollination—A Systematic Review. Biol. Conserv. 2020, 241, 108321. [Google Scholar] [CrossRef]
- Zaninotto, V.; Perrard, A.; Babiar, O.; Hansart, A.; Hignard, C.; Dajoz, I. Seasonal Variations of Pollinator Assemblages among Urban and Rural Habitats: A Comparative Approach Using a Standardized Plant Community. Insects 2021, 12, 199. [Google Scholar] [CrossRef]
- Baldock, K.C. Opportunities and Threats for Pollinator Conservation in Global Towns and Cities. Curr. Opin. Insect Sci. 2020, 38, 63–71. [Google Scholar] [CrossRef]
- Hall, D.M.; Camilo, G.R.; Tonietto, R.K.; Ollerton, J.; Ahrné, K.; Arduser, M.; Ascher, J.S.; Baldock, K.C.R.; Fowler, R.; Frankie, G.; et al. The City as a Refuge for Insect Pollinators: Insect Pollinators. Conserv. Biol. 2017, 31, 24–29. [Google Scholar] [CrossRef]
- Harrison, T.; Winfree, R. Urban Drivers of Plant-pollinator Interactions. Funct. Ecol. 2015, 29, 879–888. [Google Scholar] [CrossRef]
- Marquardt, M.; Kienbaum, L.; Kretschmer, L.A.; Penell, A.; Schweikert, K.; Ruttensperger, U.; Rosenkranz, P. Evaluation of the Importance of Ornamental Plants for Pollinators in Urban and Suburban Areas in Stuttgart, Germany. Urban Ecosyst. 2021, 24, 811–825. [Google Scholar] [CrossRef]
- Theodorou, P.; Radzevičiūtė, R.; Lentendu, G.; Kahnt, B.; Husemann, M.; Bleidorn, C.; Settele, J.; Schweiger, O.; Grosse, I.; Wubet, T.; et al. Urban Areas as Hotspots for Bees and Pollination but Not a Panacea for All Insects. Nat. Commun. 2020, 11, 576. [Google Scholar] [CrossRef]
- Fitch, G.; Wilson, C.J.; Glaum, P.; Vaidya, C.; Simao, M.-C.; Jamieson, M.A. Does Urbanization Favour Exotic Bee Species? Implications for the Conservation of Native Bees in Cities. Biol. Lett. 2019, 15, 20190574. [Google Scholar] [CrossRef]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; Stone, G.N.; et al. Where Is the UK’s Pollinator Biodiversity? The Importance of Urban Areas for Flower-Visiting Insects. Proc. R. Soc. B 2015, 282, 20142849. [Google Scholar] [CrossRef] [PubMed]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissière, B.E. Decreasing Abundance, Increasing Diversity and Changing Structure of the Wild Bee Community (Hymenoptera: Anthophila) along an Urbanization Gradient. PLoS ONE 2014, 9, e104679. [Google Scholar] [CrossRef] [PubMed]
- Geslin, B.; Le Féon, V.; Kuhlmann, M.; Vaissière, B.E.; Dajoz, I. The Bee Fauna of Large Parks in Downtown Paris, France. Ann. Soc. Entomol. Fr. 2015, 51, 487–493. [Google Scholar] [CrossRef]
- Anderson, M.; Crubaugh, F.; Greenslit, C.; Hill, E.; Kroth, H.; Stanislawski, E.; Ribbons, R.; Del Toro, I.B.Y.O. Bees: Managing Wild Bee Biodiversity in Urban Greenspaces. PLoS ONE 2023, 18, e0281468. [Google Scholar] [CrossRef]
- Banaszak-Cibicka, W.; Twerd, L.; Fliszkiewicz, M.; Giejdasz, K.; Langowska, A. City Parks vs. Natural Areas—Is It Possible to Preserve a Natural Level of Bee Richness and Abundance in a City Park? Urban Ecosyst. 2018, 21, 599–613. [Google Scholar] [CrossRef]
- Birdshire, K.R.; Carper, A.L.; Briles, C.E. Bee Community Response to Local and Landscape Factors along an Urban-Rural Gradient. Urban Ecosyst. 2020, 23, 689–702. [Google Scholar] [CrossRef]
- Bennett, A.B.; Lovell, S. Landscape and Local Site Variables Differentially Influence Pollinators and Pollination Services in Urban Agricultural Sites. PLoS ONE 2019, 14, e0212034. [Google Scholar] [CrossRef] [PubMed]
- Dietzel, S.; Rojas-Botero, S.; Kollmann, J.; Fischer, C. Enhanced Urban Roadside Vegetation Increases Pollinator Abundance Whereas Landscape Characteristics Drive Pollination. Ecol. Indic. 2023, 147, 109980. [Google Scholar] [CrossRef]
- United Nations. The World’s Cities in 2016. Available online: https://www.un-ilibrary.org/content/books/9789210582766/read (accessed on 21 September 2023).
- McDougall, R.; Kristiansen, P.; Latty, T.; Jones, J.; Rader, R. Pollination Service Delivery Is Complex: Urban Garden Crop Yields Are Best Explained by Local Canopy Cover and Garden Scale Plant Species Richness. J. Appl. Ecol. 2022, 59, 1248–1257. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Matteson, K.C.; Minor, E.S. Diversity of Wild Bees Supports Pollination Services in an Urbanized Landscape. Oecologia 2015, 179, 811–821. [Google Scholar] [CrossRef] [PubMed]
- Cohen, H.; Philpott, S.M.; Liere, H.; Lin, B.B.; Jha, S. The Relationship between Pollinator Community and Pollination Services Is Mediated by Floral Abundance in Urban Landscapes. Urban Ecosyst. 2021, 24, 275–290. [Google Scholar] [CrossRef]
- Brittain, C.; Williams, N.; Kremen, C.; Klein, A.-M. Synergistic Effects of Non-Apis Bees and Honey Bees for Pollination Services. Proc. R. Soc. B 2013, 280, 20122767. [Google Scholar] [CrossRef] [PubMed]
- Greenleaf, S.S.; Kremen, C. Wild Bees Enhance Honey Bees’ Pollination of Hybrid Sunflower. Proc. Natl. Acad. Sci. USA 2006, 103, 13890–13895. [Google Scholar] [CrossRef]
- MacInnis, G.; Normandin, E.; Ziter, C.D. Decline in Wild Bee Species Richness Associated with Honey Bee (Apis mellifera L.) Abundance in an Urban Ecosystem. PeerJ 2023, 11, e14699. [Google Scholar] [CrossRef]
- Ropars, L.; Dajoz, I.; Fontaine, C.; Muratet, A.; Geslin, B. Wild Pollinator Activity Negatively Related to Honey Bee Colony Densities in Urban Context. PLoS ONE 2019, 14, e0222316. [Google Scholar] [CrossRef]
- Page, M.L.; Williams, N.M. Evidence of Exploitative Competition between Honey Bees and Native Bees in Two California Landscapes. J. Anim. Ecol. 2023, 92, 1802–1814. [Google Scholar] [CrossRef]
- Abrol, D.P.; Gorka, A.K.; Ansari, M.J.; Al-Ghamdi, A.; Al-Kahtani, S. Impact of Insect Pollinators on Yield and Fruit Quality of Strawberry. Saudi J. Biol. Sci. 2019, 26, 524–530. [Google Scholar] [CrossRef]
- Bänsch, S.; Tscharntke, T.; Gabriel, D.; Westphal, C. Crop Pollination Services: Complementary Resource Use by Social vs Solitary Bees Facing Crops with Contrasting Flower Supply. J. Appl. Ecol. 2021, 58, 476–485. [Google Scholar] [CrossRef]
- Gaston, A.; Osorio, S.; Denoyes, B.; Rothan, C. Applying the Solanaceae Strategies to Strawberry Crop Improvement. Trends Plant Sci. 2020, 25, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Gil-Ariza, D.J.; Amaya, I.; López-Aranda, J.M.; Sánchez-Sevilla, J.F.; Ángel Botella, M.; Valpuesta, V. Impact of Plant Breeding on the Genetic Diversity of Cultivated Strawberry as Revealed by Expressed Sequence Tag-Derived Simple Sequence Repeat Markers. J. Am. Soc. Hortic. Sci. 2009, 134, 337–347. [Google Scholar] [CrossRef]
- Groupe de Travail des Normes de Qualité des Produits Agricoles Norme CEE-ONU FFV-35 Concernant La Commercialisation et Le Contrôle de La Qualité Commerciale Des Fraises. 2021. Available online: https://unece.org/trade/standards/trade-and-uncefact/agricultural-quality-standards-wp7/fresh-fruit-and-vegetables#c55288 (accessed on 21 September 2023).
- Lado, J.; Vicente, E.; Manzzioni, A.; Ares, G. Application of a Check-All-That-Apply Question for the Evaluation of Strawberry Cultivars from a Breeding Program: A Check-All-That-Apply Question for the Evaluation of Strawberry Cultivars. J. Sci. Food Agric. 2010, 90, 2268–2275. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.M.; Denoyes-Rothan, B.; Lerceteau-Köhler, E. Strawberry. In Genome Mapping and Molecular Breeding in Plants, Volume 4 Fruits and Nuts; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 189–205. [Google Scholar]
- Andersson, G.K.S.; Rundlöf, M.; Smith, H.G. Organic Farming Improves Pollination Success in Strawberries. PLoS ONE 2012, 7, e31599. [Google Scholar] [CrossRef] [PubMed]
- Garratt, M.P.D.; Groot, G.A.; Albrecht, M.; Bosch, J.; Breeze, T.D.; Fountain, M.T.; Klein, A.M.; McKerchar, M.; Park, M.; Paxton, R.J.; et al. Opportunities to Reduce Pollination Deficits and Address Production Shortfalls in an Important Insect-pollinated Crop. Ecol. Appl. 2021, 31, e02445. [Google Scholar] [CrossRef] [PubMed]
- Climate Data for Cities Worldwide. Available online: https://en.climate-data.org/ (accessed on 14 September 2023).
- Gasic, K.; Preece, J.E.; Karp, D. Register of New Fruit and Nut Cultivars List 48. HortScience 2016, 51, 620–652. [Google Scholar] [CrossRef]
- Pierron-Darbonne, A. United States Plant Patent: Strawberry Plant Named “Deluxe”. U.S. Patent 26208P3, 15 December 2015. Available online: https://patents.google.com/patent/USPP26208P3/en (accessed on 21 September 2023).
- Klatt, B.K.; Burmeister, C.; Westphal, C.; Tscharntke, T.; Von Fragstein, M. Flower Volatiles, Crop Varieties and Bee Responses. PLoS ONE 2013, 8, e72724. [Google Scholar] [CrossRef]
- QGIS Development Team. QGIS Geographic Information System. 2020. Available online: http://www.qgis.org (accessed on 21 September 2023).
- Steffan-Dewenter, I.; Kuhn, A. Honeybee Foraging in Differentially Structured Landscapes. Proc. R. Soc. Lond. B 2003, 270, 569–575. [Google Scholar] [CrossRef]
- Fuchs, R.; Müllek, M. Pollination Problems in Styrian Oil Pumpkin Plants: Can Bumblebees Be an Alternative to Honeybees? Phyton 2004, 44, 155–165. [Google Scholar]
- Zebrowska, J. Influence of Pollination Modes on Yield Components in Strawberry (Fragaria × ananassa Duch.). Plant Breed. 1998, 117, 255–260. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022. [Google Scholar]
- Bartomeus, I.; Potts, S.G.; Steffan-Dewenter, I.; Vaissière, B.E.; Woyciechowski, M.; Krewenka, K.M.; Tscheulin, T.; Roberts, S.P.M.; Szentgyörgyi, H.; Westphal, C.; et al. Contribution of Insect Pollinators to Crop Yield and Quality Varies with Agricultural Intensification. PeerJ 2014, 2, e328. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. J. Stat. Soft 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Bartoń, K. MuMIn: Multi-Model Inference. 2023. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 21 September 2023).
- Lüdecke, D. Ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef]
- Pescie, M.A.; Strik, B.C. Thinning before Bloom Affects Fruit Size and Yield of Hardy Kiwifruit. HortScience 2004, 39, 1243–1245. [Google Scholar] [CrossRef]
- Ramsey, M.D.; Daniells, J.W.; Anderson, V.J. Effects of Sigatoka Leaf Spot (Mycosphaerella Musicola Leach) on Fruit Yields, Field Ripening and Greenlife of Bananas in North Queensland. Sci. Hortic. 1990, 41, 305–313. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Los Angeles, CA, USA, 2019. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Castle, D.; Grass, I.; Westphal, C. Fruit Quantity and Quality of Strawberries Benefit from Enhanced Pollinator Abundance at Hedgerows in Agricultural Landscapes. Agric. Ecosyst. Environ. 2019, 275, 14–22. [Google Scholar] [CrossRef]
- Herrmann, J.D.; Beye, H.; De La Broise, C.; Hartlep, H.; Diekötter, T. Positive Effects of the Pollinators Osmia Cornuta (Megachilidae) and Lucilia Sericata (Calliphoridae) on Strawberry Quality. Arthropod-Plant Interact. 2019, 13, 71–77. [Google Scholar] [CrossRef]
- MacInnis, G.; Forrest, J.R.K. Pollination by Wild Bees Yields Larger Strawberries than Pollination by Honey Bees. J. Appl. Ecol. 2019, 56, 824–832. [Google Scholar] [CrossRef]
- Wietzke, A.; Westphal, C.; Gras, P.; Kraft, M.; Pfohl, K.; Karlovsky, P.; Pawelzik, E.; Tscharntke, T.; Smit, I. Insect Pollination as a Key Factor for Strawberry Physiology and Marketable Fruit Quality. Agric. Ecosyt. Environ. 2018, 258, 197–204. [Google Scholar] [CrossRef]
- Scherr, K.D.; Jamieson, M.A. Abiotic and Biotic Drivers of Strawberry Productivity across a Rural-Urban Gradient. Basic Appl. Ecol. 2021, 57, 65–77. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Bartomeus, I.; Bommarco, R.; Klein, A.M.; Cunningham, S.A.; Aizen, M.A.; Boreux, V.; Garratt, M.P.D.; Carvalheiro, L.G.; Kremen, C.; et al. EDITOR’S CHOICE: REVIEW: Trait Matching of Flower Visitors and Crops Predicts Fruit Set Better than Trait Diversity. J. Appl. Ecol. 2015, 52, 1436–1444. [Google Scholar] [CrossRef]
- Bennett, J.M.; Steets, J.A.; Burns, J.H.; Burkle, L.A.; Vamosi, J.C.; Wolowski, M.; Arceo-Gómez, G.; Burd, M.; Durka, W.; Ellis, A.G.; et al. Land Use and Pollinator Dependency Drives Global Patterns of Pollen Limitation in the Anthropocene. Nat. Commun. 2020, 11, 3999. [Google Scholar] [CrossRef] [PubMed]
- Connelly, H.; Poveda, K.; Loeb, G. Landscape Simplification Decreases Wild Bee Pollination Services to Strawberry. Agric. Ecosyst. Environ. 2015, 211, 51–56. [Google Scholar] [CrossRef]
- Ellis, C.R.; Feltham, H.; Park, K.; Hanley, N.; Goulson, D. Seasonal Complementary in Pollinators of Soft-Fruit Crops. Basic Appl. Ecol. 2017, 19, 45–55. [Google Scholar] [CrossRef]
- Requier, F.; Pérez-Méndez, N.; Andersson, G.K.S.; Blareau, E.; Merle, I.; Garibaldi, L.A. Bee and Non-Bee Pollinator Importance for Local Food Security. Trends Ecol. Evol. 2023, 38, 196–205. [Google Scholar] [CrossRef]
- Olsson, R.L.; Brousil, M.R.; Clark, R.E.; Baine, Q.; Crowder, D.W. Interactions between Plants and Pollinators across Urban and Rural Farming Landscapes. Food Webs 2021, 27, e00194. [Google Scholar] [CrossRef]
- Knight, M.E.; Martin, A.P.; Bishop, S.; Osborne, J.L.; Hale, R.J.; Sanderson, R.A.; Goulson, D. An Interspecific Comparison of Foraging Range and Nest Density of Four Bumblebee (Bombus) Species. Mol. Ecol. 2005, 14, 1811–1820. [Google Scholar] [CrossRef]
- Grüter, C.; Ratnieks, F.L.W. Flower Constancy in Insect Pollinators: Adaptive Foraging Behaviour or Cognitive Limitation? Commun. Integr. Biol. 2011, 4, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Philpott, S.M.; Cotton, J.; Bichier, P.; Friedrich, R.L.; Moorhead, L.C.; Uno, S.; Valdez, M. Local and Landscape Drivers of Arthropod Abundance, Richness, and Trophic Composition in Urban Habitats. Urban Ecosyst. 2014, 17, 513–532. [Google Scholar] [CrossRef]
- Albano, S.; Salvado, E.; Borges, P.A.V.; Mexia, A. Floral Visitors, Their Frequency, Activity Rate and Index of Visitation Rate in the Strawberry Fields of Ribatejo, Portugal: Selection of Potential Pollinators. Part 1. Adv. Hortic. Sci. 2009, 24, 238–245. [Google Scholar]
- Marcacci, G.; Devy, S.; Wenzel, A.; Rao, V.S.; Kumar, S.S.; Nölke, N.; Belavadi, V.V.; Tscharntke, T.; Grass, I.; Westphal, C. Direct and Indirect Effects of Urbanization, Pesticides and Wild Insect Pollinators on Mango Yield. J. Appl. Ecol. 2023, 60, 2132–2143. [Google Scholar] [CrossRef]
- Çolak, A.M.; Şahinler, N.; İSlamoğlu, M. The Effect of Honeybee Pollination on Productivity and Quality of Strawberry. Alınteri Zirai Bilimler Dergisi 2017, 32, 87–90. [Google Scholar] [CrossRef]
- Chandler, C.K.; Folta, K.; Dale, A.; Whitaker, V.M.; Herrington, M. Strawberry. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Handbook of Plant Breeding; Springer: Boston, MA, USA, 2012; pp. 305–325. ISBN 978-1-4419-0763-9. [Google Scholar]
- Jamieson, A.R. Strawberry Shape: Phenotypic Variation in Length and Width. Acta Hortic. 2017, 1156, 135–140. [Google Scholar] [CrossRef]
- Tuohimetsä, S.; Hietaranta, T.; Uosukainen, M.; Kukkonen, S.; Karhu, S. Fruit Development in Artificially Self- and Cross-Pollinated Strawberries (Fragaria × ananassa) and Raspberries (Rubus idaeus). Acta Agric. Scand. Sect. Soil Plant Sci. 2014, 64, 408–415. [Google Scholar] [CrossRef]
- Trillo, A.; Herrera, J.M.; Vilà, M. Managed Bumble Bees Increase Flower Visitation but Not Fruit Weight in Polytunnel Strawberry Crops. Basic Appl. Ecol. 2018, 30, 32–40. [Google Scholar] [CrossRef]
- Muola, A.; Weber, D.; Malm, L.E.; Egan, P.A.; Glinwood, R.; Parachnowitsch, A.L.; Stenberg, J.A. Direct and Pollinator-Mediated Effects of Herbivory on Strawberry and the Potential for Improved Resistance. Front. Plant Sci. 2017, 8, 823. [Google Scholar] [CrossRef]
- Ariza, M.T.; Soria, C.; Medina-Mínguez, J.J.; Martínez-Ferri, E. Incidence of Misshapen Fruits in Strawberry Plants Grown under Tunnels Is Affected by Cultivar, Planting Date, Pollination, and Low Temperatures. HortScience 2012, 47, 1569–1573. [Google Scholar] [CrossRef]
- Nicholls, E.; Ely, A.; Birkin, L.; Basu, P.; Goulson, D. The Contribution of Small-Scale Food Production in Urban Areas to the Sustainable Development Goals: A Review and Case Study. Sustain. Sci. 2020, 15, 1585–1599. [Google Scholar] [CrossRef]
- Sircom, J. The Contribution of Epigean Insects to Commercial Cranberry Pollination. J. Acad. Entomol. Soc. 2017, 13, 31–36. [Google Scholar]
- Beattie, A.J.; Turnbull, C.; Knox, R.B.; Williams, E.G. Ant Inhibition of Pollen Function: A Possible Reason Why Ant Pollination Is Rare. Am. J. Bot 1984, 71, 421–426. [Google Scholar] [CrossRef]
- Griffiths-Lee, J.; Nicholls, E.; Goulson, D. Companion Planting to Attract Pollinators Increases the Yield and Quality of Strawberry Fruit in Gardens and Allotments. Ecol. Entomol. 2020, 45, 1025–1034. [Google Scholar] [CrossRef]
- Blareau, E.; Requier, F. Data from: Insect-Mediated Pollination of Strawberries in an Urban Environment. figshare Dataset 2023. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Pollination Treatments | Estimate | Standard Error | p Value |
|---|---|---|---|
| E + H vs. E | −0.007 | 0.042 | 0.998 |
| O vs. E | 0.116 | 0.041 | 0.027 |
| O + H vs. E | 0.062 | 0.048 | 0.570 |
| O vs. E + H | 0.122 | 0.045 | 0.033 |
| O + H vs. E + H | 0.069 | 0.051 | 0.530 |
| O + H vs. O | −0.054 | 0.050 | 0.405 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blareau, E.; Sy, P.; Daoud, K.; Requier, F. Insect-Mediated Pollination of Strawberries in an Urban Environment. Insects 2023, 14, 877. https://doi.org/10.3390/insects14110877
Blareau E, Sy P, Daoud K, Requier F. Insect-Mediated Pollination of Strawberries in an Urban Environment. Insects. 2023; 14(11):877. https://doi.org/10.3390/insects14110877
Chicago/Turabian StyleBlareau, Elsa, Pauline Sy, Karim Daoud, and Fabrice Requier. 2023. "Insect-Mediated Pollination of Strawberries in an Urban Environment" Insects 14, no. 11: 877. https://doi.org/10.3390/insects14110877