
Screening and Functional Analyses of Novel Cecropins from Insect Transcriptome


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Screening Cecropin Homologs in 1KITE Dataset and Phylogenetic Analysis of Predicted Cecropin
2.2. cDNA Cloning of Representative Cecropin Homologs
2.3. Sequence Analysis of Predicted Cecropin
2.4. Peptide Synthesis of Representative Cecropin Homologs
2.5. Secondary Structure Analysis of Cecropin with Circular Dichroism (CD)
2.6. Antimicrobial Assays
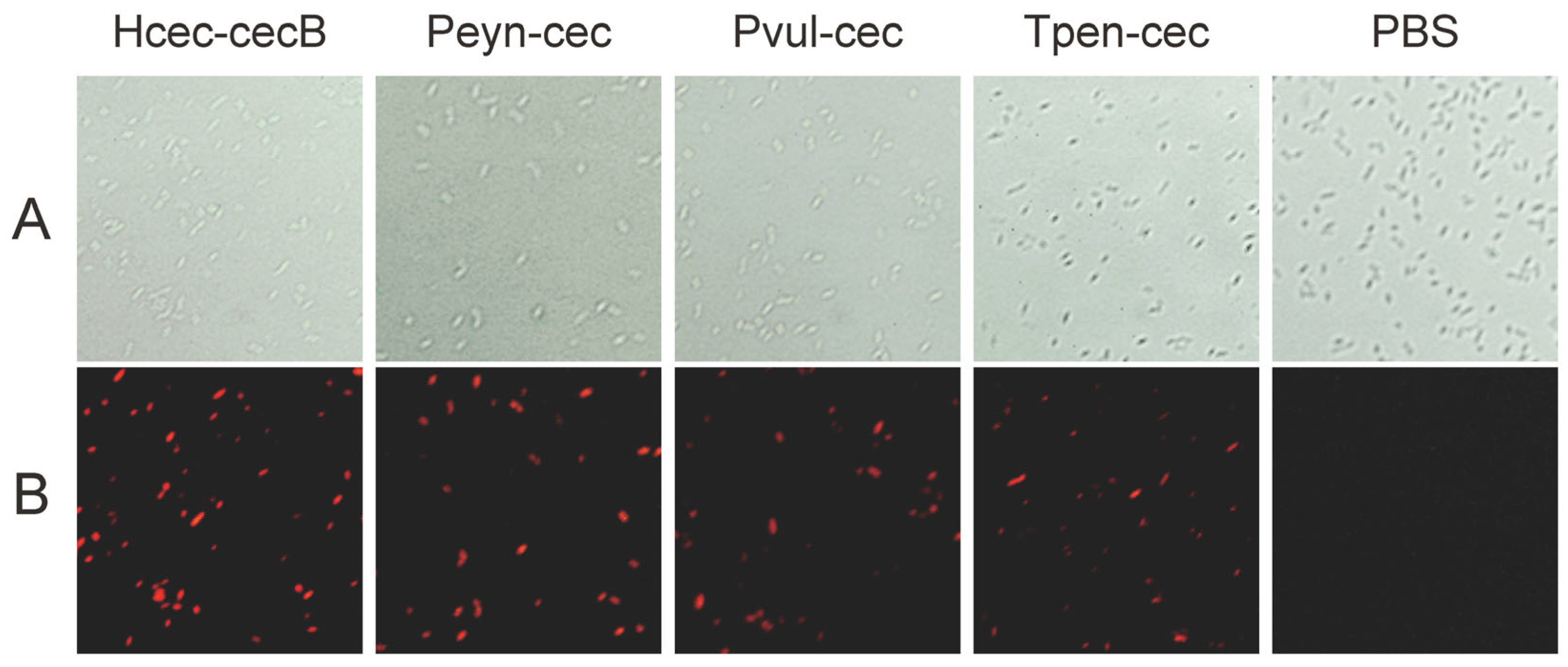
2.7. Permeability of Bacteria Membrane
2.8. Hemolytic Assay
2.9. Statistical Analysis
3. Results
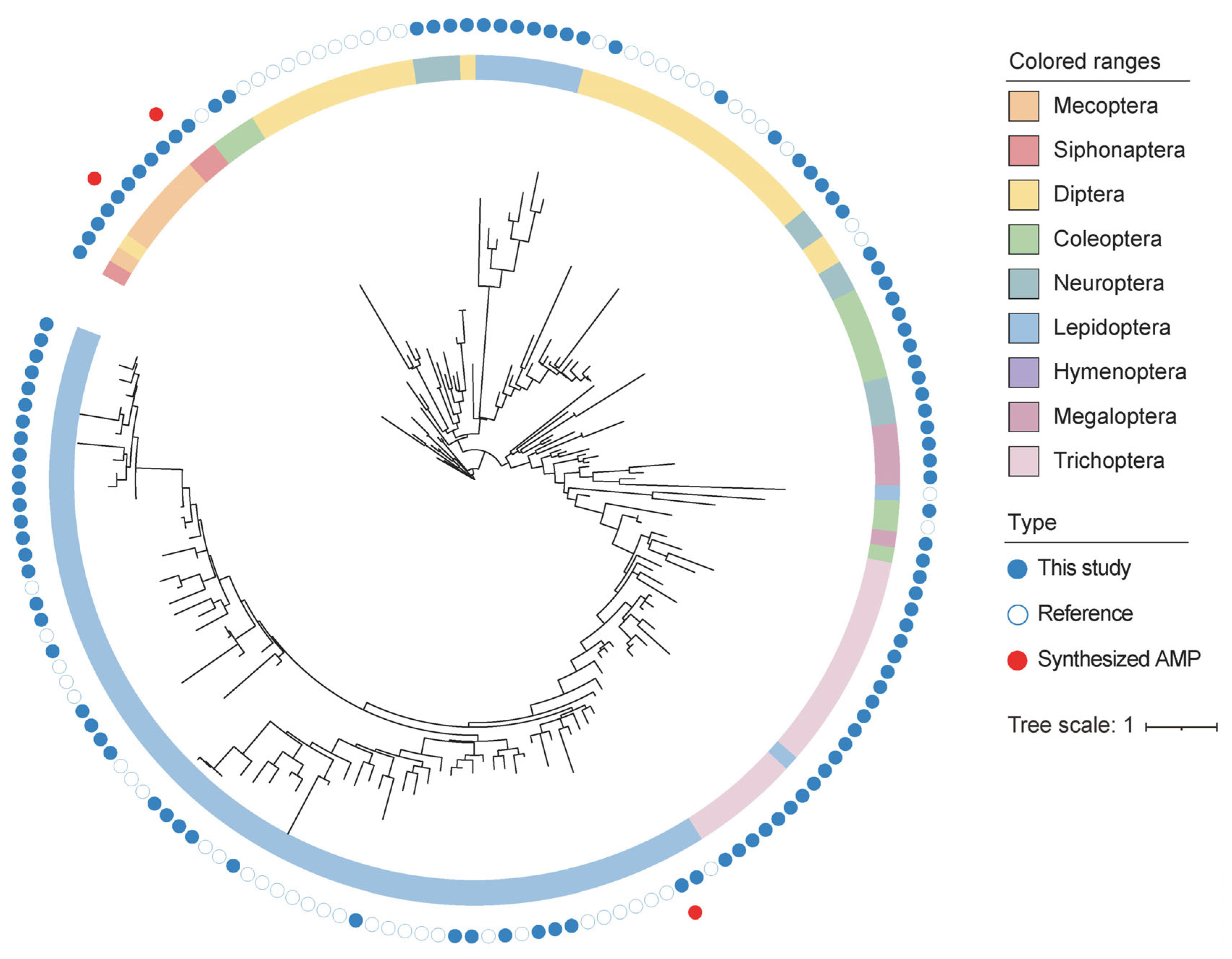
3.1. Identification of Cecropin Genes from 1KITE
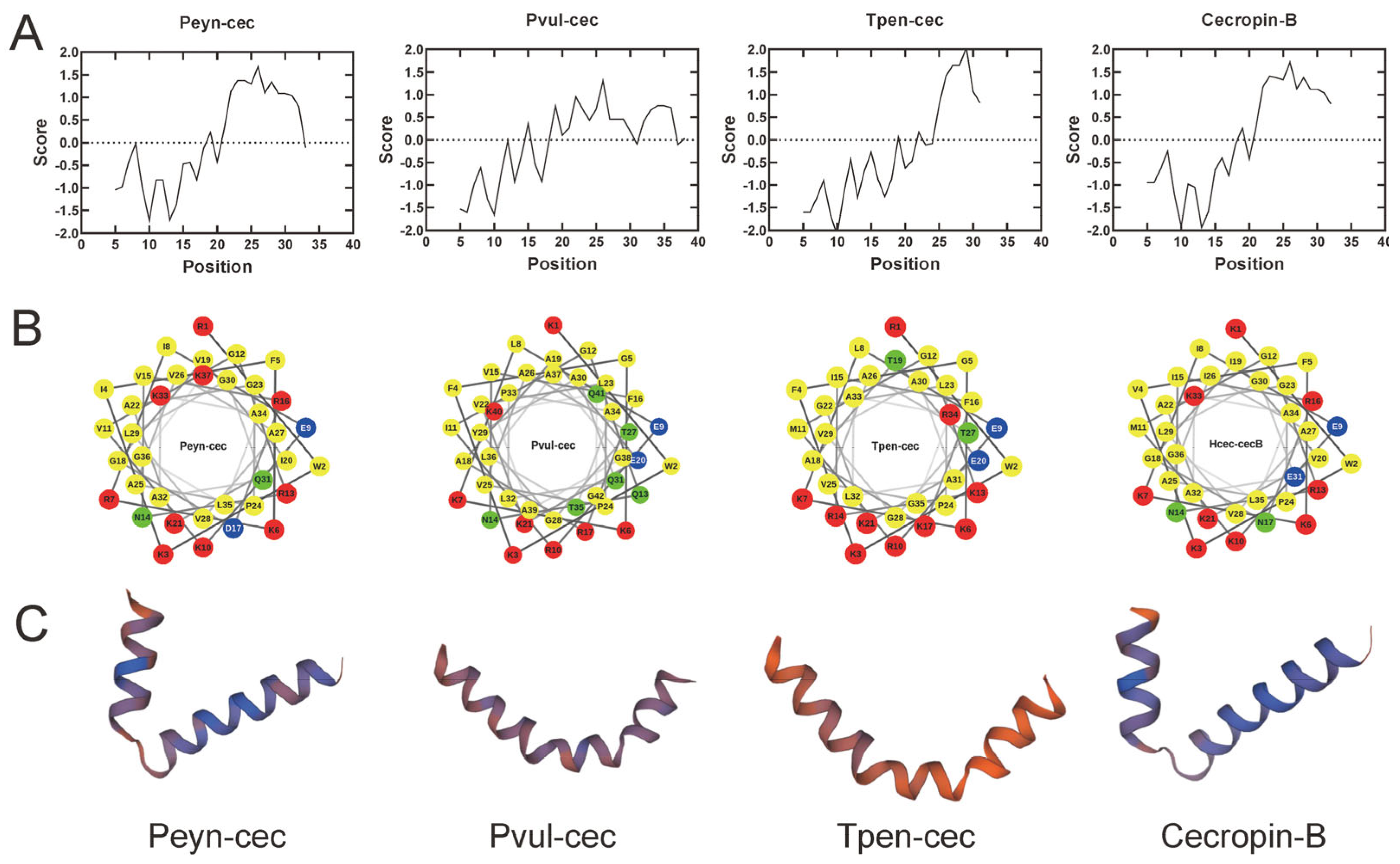
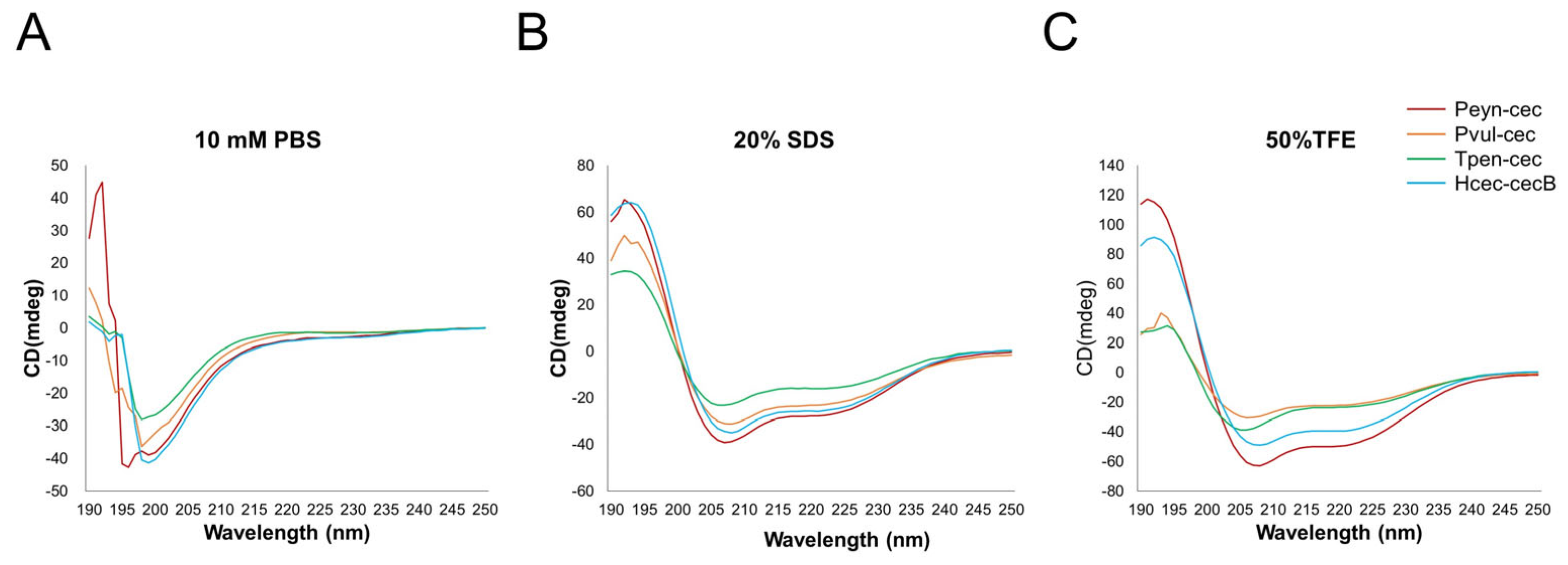
3.2. Cecropin Sequence Validation and Characteristics
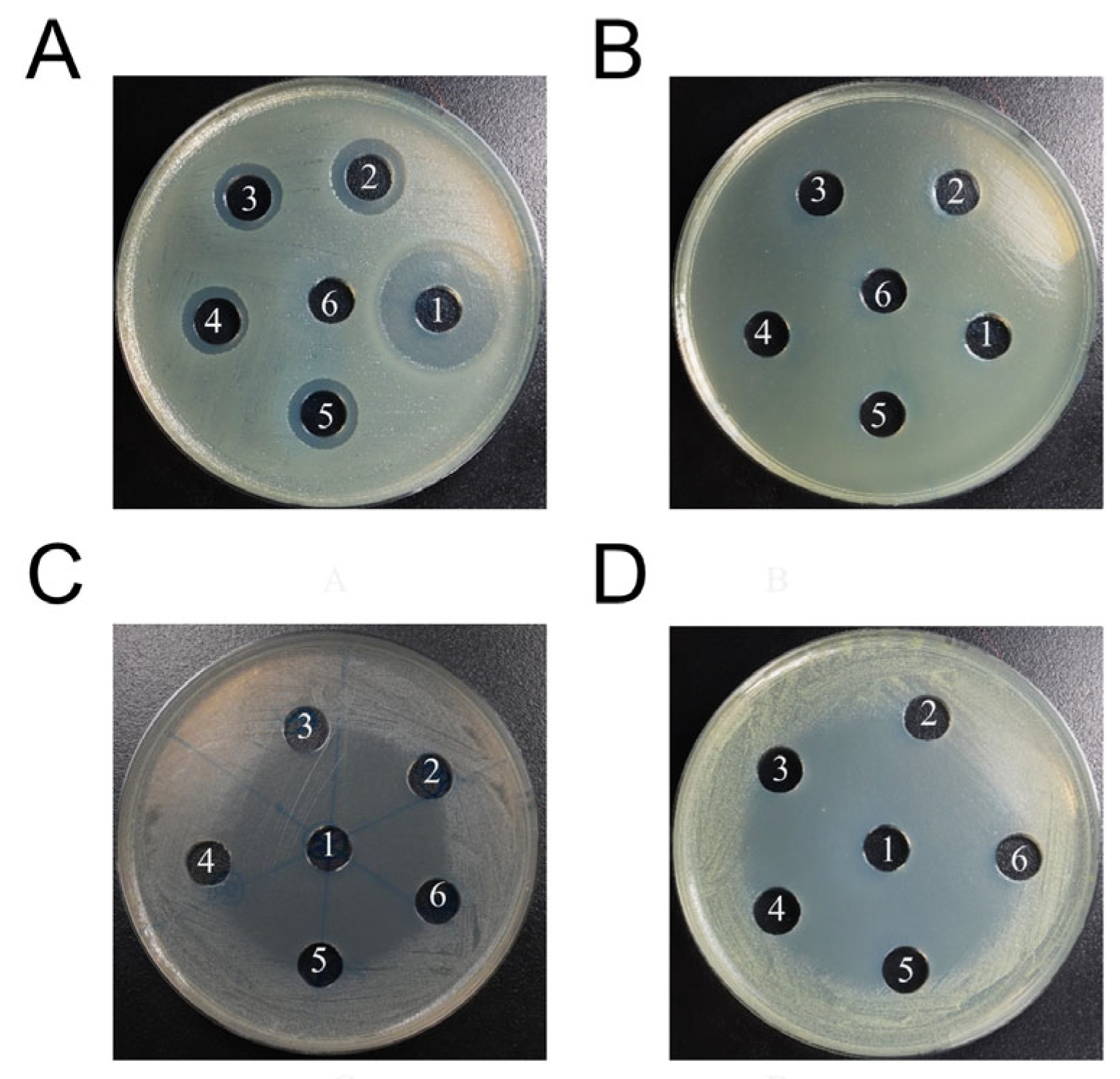
3.3. Antimicrobial Efficiency and Hemolytic Activity of the Novel Cecropins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martinez, J.L.; Baquero, F. Emergence and spread of antibiotic resistance: Setting a parameter space. Ups. J. Med. Sci. 2014, 119, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Stanton, T.B. A call for antibiotic alternatives research. Trends Microbiol. 2013, 21, 111–113. [Google Scholar] [CrossRef] [PubMed]
- von Wintersdorff, C.J.; Penders, J.; van Niekerk, J.M.; Mills, N.D.; Majumder, S.; van Alphen, L.B.; Savelkoul, P.H.; Wolffs, P.F. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal Gene Transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef]
- Boman, H.G. Antibacterial peptides: Key components needed in immunity. Cell 1991, 65, 205–207. [Google Scholar] [CrossRef]
- Cederlund, A.; Gudmundsson, G.H.; Agerberth, B. Antimicrobial peptides important in innate immunity. FEBS J. 2011, 278, 3942–3951. [Google Scholar] [CrossRef]
- Sato, H.; Feix, J.B. Peptide-membrane interactions and mechanisms of membrane destruction by amphipathic alpha-helical antimicrobial peptides. Biochim. Biophys. Acta 2006, 1758, 1245–1256. [Google Scholar] [CrossRef]
- Choi, H.; Hwang, J.S.; Lee, D.G. Identification of a novel antimicrobial peptide, scolopendin 1, derived from centipede Scolopendra subspinipes mutilans and its antifungal mechanism. Insect Mol. Biol. 2014, 23, 788–799. [Google Scholar] [CrossRef]
- Feng, M.; Fei, S.; Xia, J.; Labropoulou, V.; Swevers, L.; Sun, J. Antimicrobial peptides as potential antiviral factors in insect antiviral immune response. Front. Immunol. 2020, 11, 2030. [Google Scholar] [CrossRef]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef]
- Torrent, M.; Pulido, D.; Rivas, L.; Andreu, D. Antimicrobial peptide action on parasites. Curr. Drug Targets 2012, 13, 1138–1147. [Google Scholar] [CrossRef]
- Hancock, R.E.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef]
- Vale, N.; Aguiar, L.; Gomes, P. Antimicrobial peptides: A new class of antimalarial drugs? Front. Pharmacol. 2014, 5, 275. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.W.; Lee, J.H.; Subramaniyam, S.; Yun, E.Y.; Kim, I.; Park, J.; Hwang, J.S. De novo transcriptome analysis and detection of antimicrobial peptides of the American Cockroach Periplaneta americana (Linnaeus). PLoS ONE 2016, 11, e0155304. [Google Scholar] [CrossRef] [PubMed]
- Vilcinskas, A.; Mukherjee, K.; Vogel, H. Expansion of the antimicrobial peptide repertoire in the invasive ladybird Harmonia axyridis. Proc. Biol. Sci. 2013, 280, 20122113. [Google Scholar] [CrossRef]
- Gerardo, N.M.; Altincicek, B.; Anselme, C.; Atamian, H.; Barribeau, S.M.; de Vos, M.; Duncan, E.J.; Evans, J.D.; Gabaldon, T.; Ghanim, M.; et al. Immunity and other defenses in pea aphids, Acyrthosiphon pisum. Genome Biol. 2010, 11, R21. [Google Scholar] [CrossRef] [PubMed]
- Mylonakis, E.; Podsiadlowski, L.; Muhammed, M.; Vilcinskas, A. Diversity, evolution and medical applications of insect antimicrobial peptides. Philos. Trans. R. Soc. Lond B Biol. Sci. 2016, 371, 20150290. [Google Scholar] [CrossRef] [PubMed]
- Boman, H.G.; Nilsson-Faye, I.; Paul, K.; Rasmuson, T., Jr. Insect immunity. I. Characteristics of an inducible cell-free antibacterial reaction in hemolymph of Samia cynthia pupae. Infect. Immun. 1974, 10, 136–145. [Google Scholar] [CrossRef]
- Faye, I.; Pye, A.; Rasmuson, T.; Boman, H.G.; Boman, I.A. Insect immunity 11. Simultaneous induction of antibacterial activity and selection synthesis of some hemolymph proteins in diapausing pupae of Hyalophora cecropia and Samia cynthia. Infect. Immun. 1975, 12, 1426–1438. [Google Scholar] [CrossRef]
- Kim, S.R.; Hong, M.Y.; Park, S.W.; Choi, K.H.; Yun, E.Y.; Goo, T.W.; Kang, S.W.; Suh, H.J.; Kim, I.; Hwang, J.S. Characterization and cDNA cloning of a cecropin-like antimicrobial peptide, papiliocin, from the swallowtail butterfly, Papilio xuthus. Mol. Cells 2010, 29, 419–423. [Google Scholar] [CrossRef] [PubMed]
- Vizioli, J.; Bulet, P.; Charlet, M.; Lowenberger, C.; Blass, C.; Muller, H.M.; Dimopoulos, G.; Hoffmann, J.; Kafatos, F.C.; Richman, A. Cloning and analysis of a cecropin gene from the malaria vector mosquito, Anopheles gambiae. Insect Mol. Biol. 2000, 9, 75–84. [Google Scholar] [CrossRef] [PubMed]
- DeLucca, A.J.; Bland, J.M.; Jacks, T.J.; Grimm, C.; Cleveland, T.E.; Walsh, T.J. Fungicidal activity of cecropin A. Antimicrob. Agents Chemother. 1997, 41, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Engstrom, A.; Bennich, H.; Kapur, R.; Boman, H.G. Insect immunity: Isolation and structure of cecropin D and four minor antibacterial components from Cecropia pupae. Eur. J. Biochem. 1982, 127, 207–217. [Google Scholar] [CrossRef]
- Moore, A.J.; Beazley, W.D.; Bibby, M.C.; Devine, D.A. Antimicrobial activity of cecropins. J. Antimicrob. Chemother. 1996, 37, 1077–1089. [Google Scholar] [CrossRef]
- Dickinson, L.; Russell, V.; Dunn, P.E. A family of bacteria-regulated, cecropin D-like peptides from Manduca sexta. J. Biol. Chem. 1988, 263, 19424–19429. [Google Scholar] [CrossRef]
- Kang, D.; Liu, G.; Gunne, H.; Steiner, H. PCR differential display of immune gene expression in Trichoplusia ni. Insect Biochem. Mol. Biol. 1996, 26, 177–184. [Google Scholar] [CrossRef]
- Saito, A.; Ueda, K.; Imamura, M.; Atsumi, S.; Tabunoki, H.; Miura, N.; Watanabe, A.; Kitami, M.; Sato, R. Purification and cDNA cloning of a cecropin from the longicorn beetle, Acalolepta luxuriosa. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 142, 317–323. [Google Scholar] [CrossRef]
- van Hofsten, P.; Faye, I.; Kockum, K.; Lee, J.Y.; Xanthopoulos, K.G.; Boman, I.A.; Boman, H.G.; Engstrom, A.; Andreu, D.; Merrifield, R.B. Molecular cloning, cDNA sequencing, and chemical synthesis of cecropin B from Hyalophora cecropia. Proc. Natl. Acad. Sci. USA 1985, 82, 2240–2243. [Google Scholar] [CrossRef]
- Waghu, F.H.; Barai, R.S.; Gurung, P.; Idicula-Thomas, S. CAMPR3: A database on sequences, structures and signatures of antimicrobial peptides. Nucleic Acids Res. 2016, 44, D1094–D1097. [Google Scholar] [CrossRef]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. HMMER web server: 2015 update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Kaushal, A.; Gupta, K.; Shah, R.; van Hoek, M.L. Antimicrobial activity of mosquito cecropin peptides against Francisella. Dev. Comp. Immunol. 2016, 63, 171–180. [Google Scholar] [CrossRef]
- Li, Z.; Mao, R.; Teng, D.; Hao, Y.; Chen, H.; Wang, X.; Wang, X.; Yang, N.; Wang, J. Antibacterial and immunomodulatory activities of insect defensins-DLP2 and DLP4 against multidrug-resistant Staphylococcus aureus. Sci. Rep. 2017, 7, 12124. [Google Scholar] [CrossRef]
- Diamond, G.; Beckloff, N.; Weinberg, A.; Kisich, K.O. The roles of antimicrobial peptides in innate host defense. Curr. Pharm. Des. 2009, 15, 2377–2392. [Google Scholar] [CrossRef]
- Amaral, A.C.; Silva, O.N.; Mundim, N.C.; de Carvalho, M.J.; Migliolo, L.; Leite, J.R.; Prates, M.V.; Bocca, A.L.; Franco, O.L.; Felipe, M.S. Predicting antimicrobial peptides from eukaryotic genomes: In silico strategies to develop antibiotics. Peptides 2012, 37, 301–308. [Google Scholar] [CrossRef]
- Huang, K.Y.; Chang, T.H.; Jhong, J.H.; Chi, Y.H.; Li, W.C.; Chan, C.L.; Robert Lai, K.; Lee, T.Y. Identification of natural antimicrobial peptides from bacteria through metagenomic and metatranscriptomic analysis of high-throughput transcriptome data of Taiwanese oolong teas. BMC Syst. Biol. 2017, 11, 131. [Google Scholar] [CrossRef]
- Kim, D.; Soundrarajan, N.; Lee, J.; Cho, H.S.; Choi, M.; Cha, S.Y.; Ahn, B.; Jeon, H.; Le, M.T.; Song, H.; et al. Genomewide analysis of the antimicrobial peptides in Python bivittatus and characterization of cathelicidins with potent antimicrobial activity and low cytotoxicity. Antimicrob. Agents Chemother. 2017, 61, e00530-17. [Google Scholar] [CrossRef]
- Ponnuvel, K.M.; Subhasri, N.; Sirigineedi, S.; Murthy, G.N.; Vijayaprakash, N.B. Molecular evolution of the cecropin multigene family in silkworm Bombyx mori. Bioinformation 2010, 5, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Onsins, S.; Aguade, M. Molecular evolution of the Cecropin multigene family in Drosophila. Functional genes vs. pseudogenes. Genetics 1998, 150, 157–171. [Google Scholar] [CrossRef] [PubMed]
- Moretta, A.; Salvia, R.; Scieuzo, C.; Di Somma, A.; Vogel, H.; Pucci, P.; Sgambato, A.; Wolff, M.; Falabella, P. A bioinformatic study of antimicrobial peptides identified in the Black Soldier Fly (BSF) Hermetia illucens (Diptera: Stratiomyidae). Sci. Rep. 2020, 10, 16875. [Google Scholar] [CrossRef] [PubMed]
- Moretta, A.; Scieuzo, C.; Salvia, R.; Popovic, Z.D.; Sgambato, A.; Falabella, P. Tools in the era of multidrug resistance in bacteria: Applications for new antimicrobial peptides discovery. Curr. Pharm. Des. 2022, 28, 2856–2866. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Yi, Y.; Liang, L.; Shi, Q. High throughput identification of antimicrobial peptides from fish gastrointestinal microbiota. Toxins 2017, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Sutherland, D.; Aninta, S.I.; Louie, N.; Nip, K.M.; Li, C.; Yanai, A.; Coombe, L.; Warren, R.L.; Helbing, C.C.; et al. Mining amphibian and insect transcriptomes for antimicrobial peptide sequences with rAMPage. Antibiotics 2022, 11, 952. [Google Scholar] [CrossRef]
- Yang, X.; Lee, W.H.; Zhang, Y. Extremely abundant antimicrobial peptides existed in the skins of nine kinds of Chinese odorous frogs. J. Proteome Res. 2012, 11, 306–319. [Google Scholar] [CrossRef]
- Christensen, B.; Fink, J.; Merrifield, R.B.; Mauzerall, D. Channel-forming properties of cecropins and related model compounds incorporated into planar lipid membranes. Proc. Natl. Acad. Sci. USA 1988, 85, 5072–5076. [Google Scholar] [CrossRef]
- Lei, M.; Jayaraman, A.; Van Deventer, J.A.; Lee, K. Engineering selectively targeting antimicrobial peptides. Annu. Rev. Biomed. Eng. 2021, 23, 339–357. [Google Scholar] [CrossRef]
- Szymczak, P.; Mozejko, M.; Grzegorzek, T.; Jurczak, R.; Bauer, M.; Neubauer, D.; Sikora, K.; Michalski, M.; Sroka, J.; Setny, P.; et al. Discovering highly potent antimicrobial peptides with deep generative model HydrAMP. Nat. Commun. 2023, 14, 1453. [Google Scholar] [CrossRef]
- Westerhoff, H.V.; Zasloff, M.; Rosner, J.L.; Hendler, R.W.; De Waal, A.; Vaz Gomes, A.; Jongsma, P.M.; Riethorst, A.; Juretic, D. Functional synergism of the magainins PGLa and magainin-2 in Escherichia coli, tumor cells and liposomes. Eur. J. Biochem. 1995, 228, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Gueguen, Y.; Bernard, R.; Julie, F.; Paulina, S.; Delphine, D.G.; Franck, V.; Philippe, B.; Evelyne, B. Oyster hemocytes express a proline-rich peptide displaying synergistic antimicrobial activity with a defensin. Mol. Immunol. 2009, 46, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Chae, J.H.; Kurokawa, K.; So, Y.I.; Hwang, H.O.; Kim, M.S.; Park, J.W.; Jo, Y.H.; Lee, Y.S.; Lee, B.L. Purification and characterization of tenecin 4, a new anti-Gram-negative bacterial peptide, from the beetle Tenebrio molitor. Dev. Comp. Immunol. 2012, 36, 540–546. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers in 5′-3′ Direction |
|---|---|
| Peyn-cec | CAGCATCCGAGAACAAAAGAACT |
| TGCTTCTTAAAATATAATGAAT | |
| Pvul-cec | GGTGATACCGAAAGTTCACAGC |
| AATTCATCCTTGTTTTGCACCTG | |
| Tpen-cec | TCGTACATCTGAAGTGAAGA |
| ATGCCTAATCAACCTCTGG |
| Peptide | MIC (µg/mL) | |||
|---|---|---|---|---|
| Peyn-cec | Pvul-cec | Tpen-cec | Hcec-cecB | |
| Gram-positive bacteria | ||||
| M. luteus ATCC9341 | >128.0 | >128.0 | >128.0 | >128.0 |
| S. aureus ATCC6538 | >128.0 | >128.0 | >128.0 | >128.0 |
| Gram-negative bacteria | ||||
| E. coli ATCC25922 | 8.0 | 8.0 | 8.0 | 8.0 |
| P. aeruginosa CGMCC 1.10712 | 32.0 | 32.0 | 32.0 | 16.0 |
| Peptide | Zone of Inhibition (mm) | ||||
|---|---|---|---|---|---|
| Peyn-cec | Pvul-cec | Tpen-cec | Hcec-cecB | Ampicillin | |
| Gram-positive bacteria | |||||
| M. luteus ATCC9341 | - | - | - | - | 59.49 ± 0.79 |
| S. aureus ATCC6538 | - | - | - | - | 49.57 ± 1.69 |
| Gram-negative bacteria | |||||
| E. coli ATCC25922 | 13.30 ± 0.75 | 15.43 ± 0.56 | 13.46 ± 0.37 | 14.30 ± 0.24 | 25.18 ± 1.10 |
| P. aeruginosa CGMCC 1.10712 | - | - | - | - | <6 |
| Peptides | Hemolysis (%) | |||
|---|---|---|---|---|
| 1 µg/mL | 5 µg/mL | 20 µg/mL | 50 µg/mL | |
| Peyn-cec | 0.07 ± 0.24% | 2.71 ± 0.78% | 5.78 ± 0.58% | 6.37 ± 0.57% |
| Pvul-cec | 2.01 ± 0.76% | 1.59 ± 1.18% | 1.90 ± 0.72% | 2.19 ± 0.92% |
| Tpen-cec | 0.05 ± 0.15% | 0.67 ± 0.48% | 1.32 ± 1.24% | 1.72 ± 1.28% |
| Hcec-cecB | 0.46 ± 0.07% | 0.97 ± 0.77% | 1.25 ± 0.20% | 1.75 ± 0.57% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Tang, M.; Luo, S.; Zhou, X. Screening and Functional Analyses of Novel Cecropins from Insect Transcriptome. Insects 2023, 14, 794. https://doi.org/10.3390/insects14100794
Guo L, Tang M, Luo S, Zhou X. Screening and Functional Analyses of Novel Cecropins from Insect Transcriptome. Insects. 2023; 14(10):794. https://doi.org/10.3390/insects14100794
Chicago/Turabian StyleGuo, Lizhen, Min Tang, Shiqi Luo, and Xin Zhou. 2023. "Screening and Functional Analyses of Novel Cecropins from Insect Transcriptome" Insects 14, no. 10: 794. https://doi.org/10.3390/insects14100794