
Ant Community Is Not Influenced by the Addition of Olive Mill Pomace Compost in Two Different Olive Crop Managements

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Location
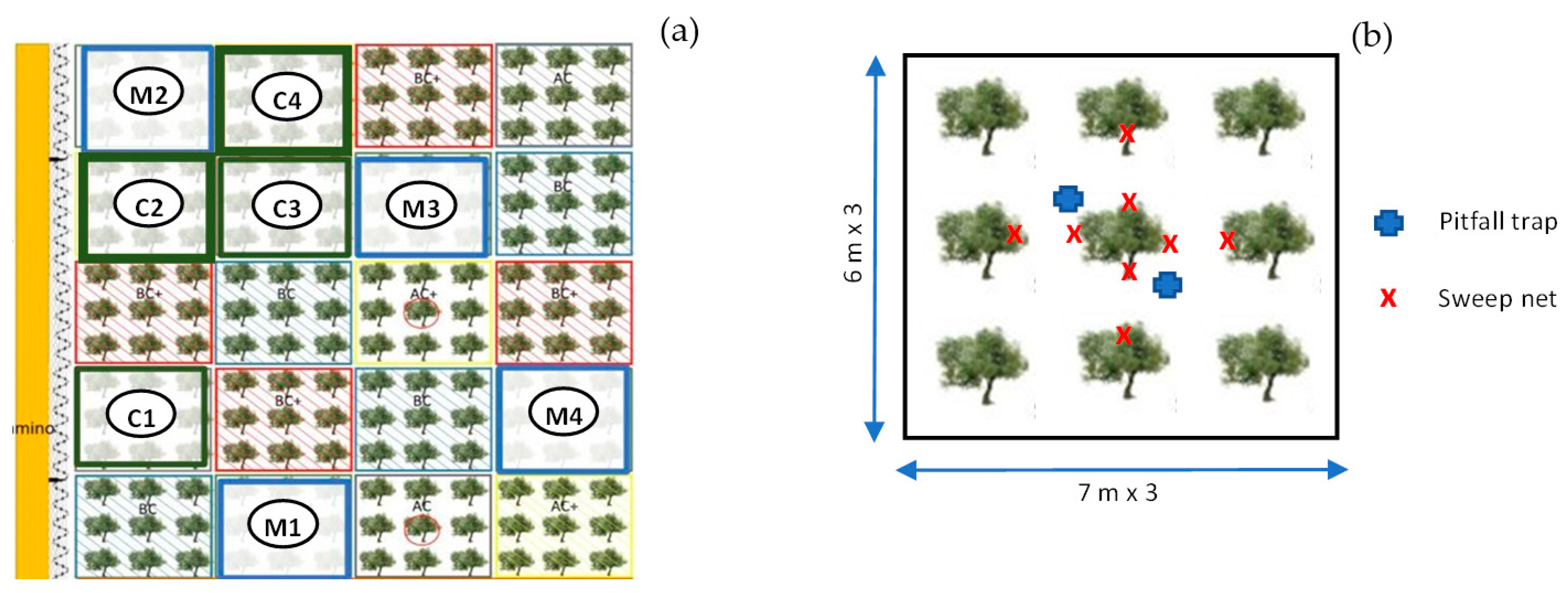
2.2. Experimental Design
2.3. Sampling
2.4. Data Analysis
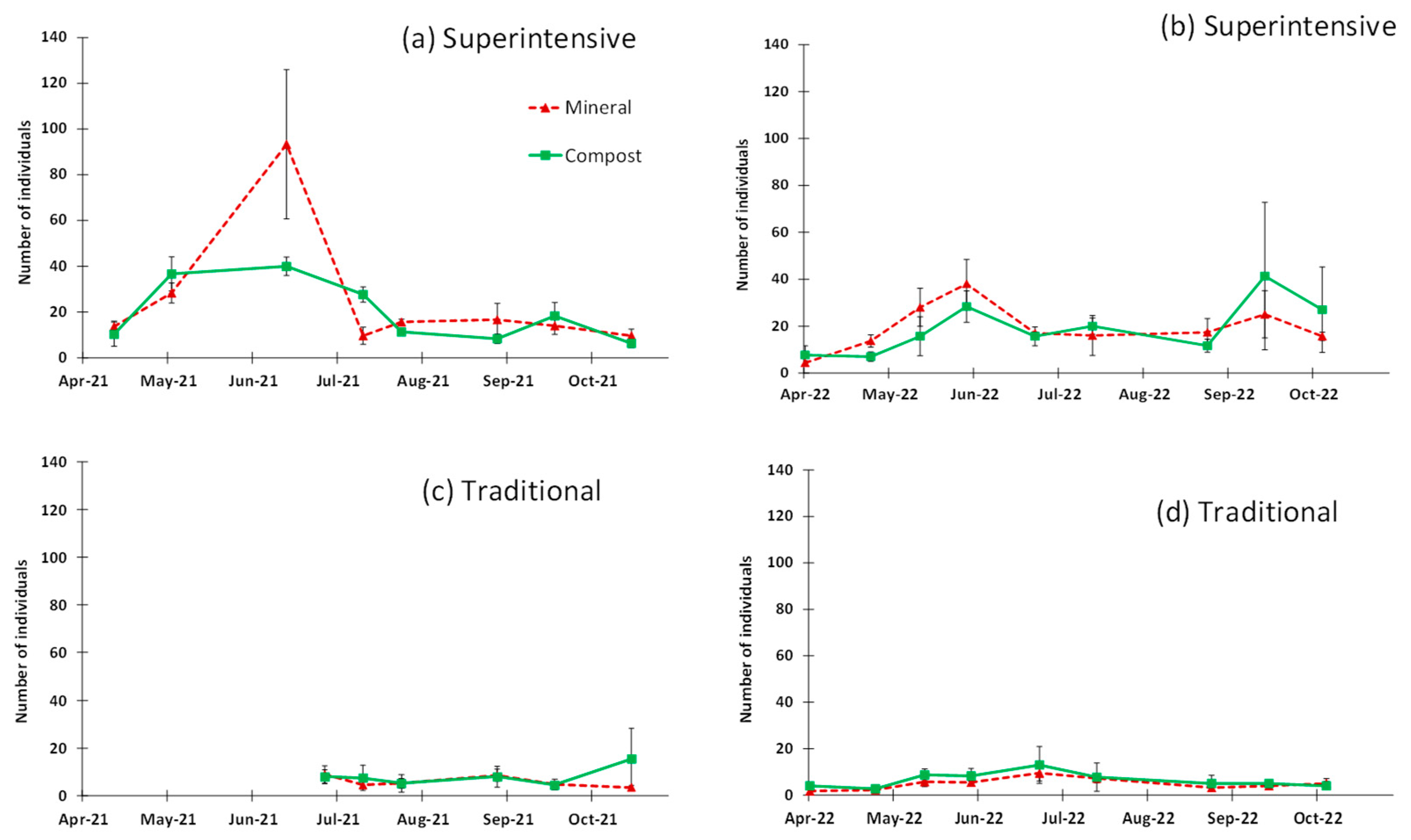
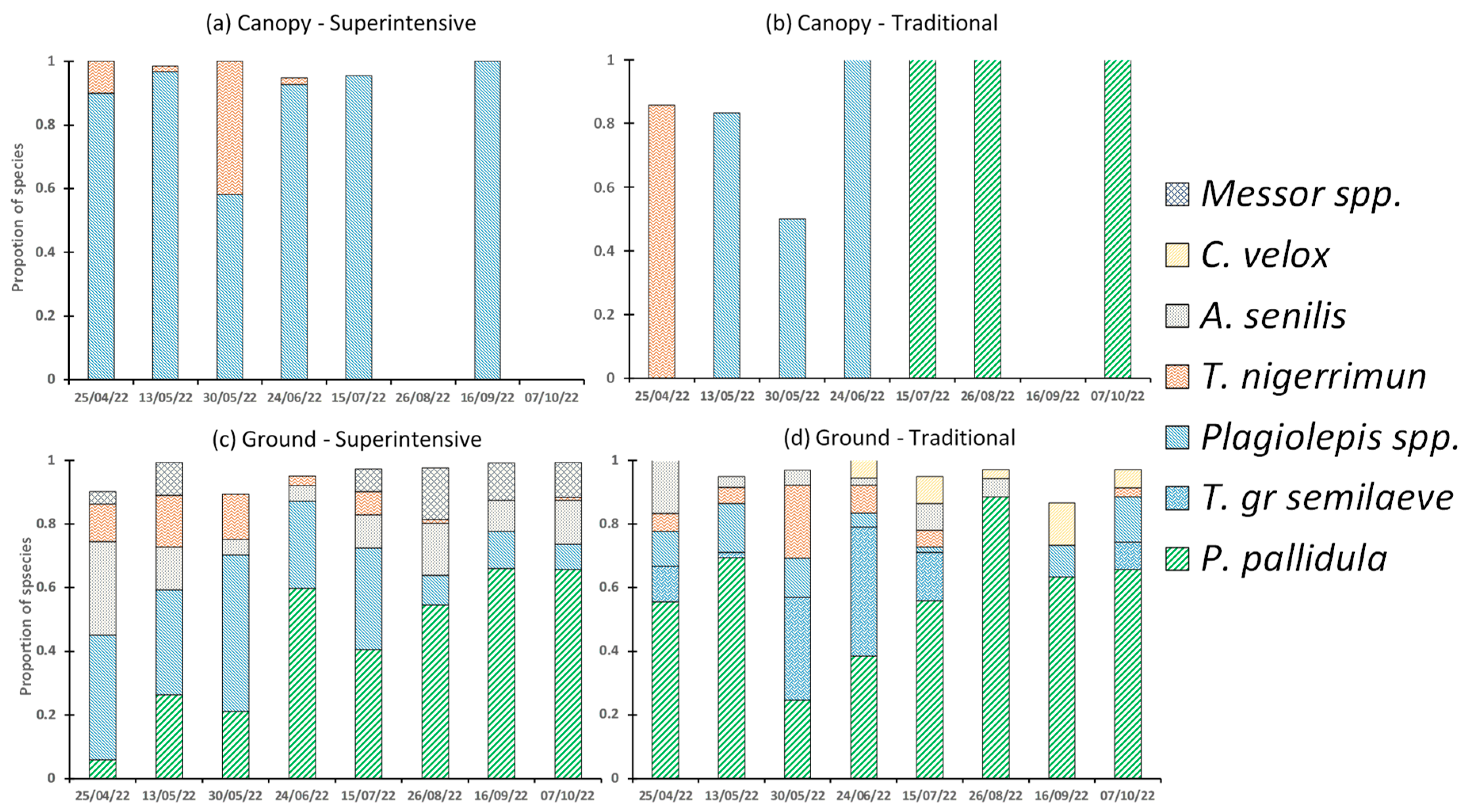
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Superintensive | Traditional | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2021 | 2021 | |||||||||||
| Mineral | Compost | Mineral | Compost | |||||||||
| Foliar | Irrigation | Total | Foliar | Compost | Total | Foliar | Mineral | Total | Foliar | Compost | Total | |
| N | 6.4 | -- | 6.4 | 6.4 | 14.1 | 20.5 | -- | -- | -- | -- | 14.1 | -- |
| P | -- | -- | -- | -- | 93.8 | 93.8 | -- | -- | -- | -- | 93.8 | -- |
| K | 22.4 | -- | 22.4 | 22.4 | 477.0 | 499.4 | -- | -- | -- | -- | 477.0 | -- |
| 2022 | 2022 | |||||||||||
| Mineral | Compost | Mineral | Compost | |||||||||
| Foliar | Irrigation | Total | Foliar | Compost | Total | Foliar | Mineral | Total | Foliar | Compost | Total | |
| N | 6.4 | 119.0 | 125.4 | 6.4 | 14.1 | 20.5 | 8.0 | 72.9 | 80.9 | 8.0 | 14.1 | 22.1 |
| P | -- | 59.5 | 59.5 | -- | 93.8 | 93.8 | -- | 24.3 | 24.3 | -- | 93.8 | 93.8 |
| K | 22.4 | 178.6 | 201.0 | 22.4 | 477.0 | 499.4 | 28.0 | 97.1 | 125.1 | 28.0 | 477.0 | 505.0 |
| SUPERINTENSIVE GROVE | ||
| Date | Product used | Used against |
| 1 January 2021 | Glyphosate 36% | Mono and Dicotyledoneae a |
| MCPA 40% | Dicotyledoneae a | |
| 15 February 2021 | Copper oxychloride 52% | Venturia oleaginea-Other diseases |
| 19 April 2021 | Lambda-cyhalothrin 10% | Prays oleae |
| 19 May 2021 | Deltamethrin 2.5% | Bactrocera oleae-Prays oleae |
| 22 June 2021 | Glyphosate 36% | Mono and Dicotyledoneae a |
| MCPA 40% | Dicotyledoneae a | |
| 23 June 2021 | Phosmet 50% | Bactrocera oleae-Prays oleae-Palpita unionalis |
| 5 November 2021 | Copper oxychloride 52% | Venturia oleaginea-Other diseases |
| Kresoxim methyl 50% | ||
| 16 February 2022 | Copper oxychloride 52% | Venturia oleaginea |
| 20 April 2022 | Glyphosate 36% | Mono and Dicotyledoneae a |
| 26 April 2022 | Deltamethrin 2.5% | Prays oleae |
| 27 May 2022 | Kresoxim methyl 50% | Venturia oleaginea |
| Lambda-cyhalothrin 10% | Prays oleae | |
| 2 June 2022 | Glyphosate 36% | Mono and Dicotyledoneae a |
| 22 June 2022 | Sulphur 80% | Eriophyidae |
| 13 July 2022 | Phosmet 50% | Bactrocera oleae-Prays oleae-Palpita unionalis |
| 4 August 2022 | Phosmet 50% | Bactrocera oleae-Prays oleae-Palpita unionalis |
| 26 October 2022 | Copper oxychloride 52% | Venturia oleaginea |
| TRADITIONAL GROVE | ||
| Date | Product used | Used against |
| 5 November 2021 | Copper oxychloride 52% | Venturia oleaginea |
| 26 April 2022 | Deltamethrin 2.5% | Prays oleae |
| 30 May 2022 | Deltamethrin 2.5% | Prays oleae |
| Difenoconazole 23.5% | Venturia oleaginea | |
| 13 July 2022 | Phosmet 50% | Bactrocera oleae-Prays oleae-Palpita unionalis |
| 4 August 2022 | Phosmet 50% | Bactrocera oleae-Prays oleae-Palpita unionalis |
| 27 October 2022 | Copper oxychloride 52% | Venturia oleaginea |
| 2021 | |||
| Superintensive | Traditional | ||
| Sweep net | Pitfall trap | Sweep net | Pitfall trap |
| 12 April 2021 | 12 April 2021 | ||
| 3 May 2021 | 3 May 2021 | ||
| 14 May 2021 | |||
| 14 June 2021 | 14 June 2021 | 28 June 2021 | 28 June 2021 |
| 12 July 2021 | 12 July 2021 | 12 July 2021 | 12 July 2021 |
| 26 July 2021 | 26 July 2021 | 26 July 2021 | 26 July 2021 |
| 30 August 2021 | 30 August 2021 | 30 August 2021 | 30 August 2021 |
| 20 September 2021 | 20 September 2021 | 20 September 2021 | 20 September 2021 |
| 18 October 2021 | 18 October 2021 | 18 October 2021 | 18 October 2021 |
| 2022 | |||
| Superintensive | Traditional | ||
| Sweep net | Pitfall trap | Sweep net | Pitfall trap |
| 1 April 2022 | 1 April 2022 | 1 April 2022 | 1 April 2022 |
| 25 April 2022 | 25 April 2022 | 25 April 2022 | 25 April 2022 |
| 13 May 2022 | 13 May 2022 | 13 May 2022 | 13 May 2022 |
| 30 May 2022 | 30 May 2022 | 30 May 2022 | 30 May 2022 |
| 24 June 2022 | 24 June 2022 | 24 June 2022 | 24 June 2022 |
| 15 July 2022 | 15 July 2022 | 15 July 2022 | 15 July 2022 |
| 26 August 2022 | 26 August 2022 | 26 August 2022 | 26 August 2022 |
| 16 September 2022 | 16 September 2022 | 16 September 2022 | 16 September 2022 |
| 7 October 2022 | 7 October 2022 | 7 October 2022 | 7 October 2022 |
Appendix B
| SWEEP NET | ||||||||
| Superintensive | Traditional | |||||||
| Shannon | 1.04 | 1.49 | ||||||
| Simpson | 0.59 | 0.73 | ||||||
| Simpson_Inverse | 2.47 | 3.74 | ||||||
| Simpson_Unbias | 0.60 | 0.76 | ||||||
| With sampling dates | ||||||||
| Shannon | 0.34 | (0.16) | 0.29 | (0.11) | ||||
| Simpson | 0.19 | (0.10) | 0.19 | (0.07) | ||||
| Simpson_Inverse | 1.41 | (0.30) | 1.31 | (0.14) | ||||
| Simpson_Unbias | 0.19 | (0.10) | 0.29 | (0.14) | ||||
| PITFALL TRAPS | ||||||||
| Superintensive | Traditional | |||||||
| Mineral | Compost | Mineral | Compost | |||||
| Shannon | 1.75 | 1.69 | 1.63 | 1.34 | ||||
| Simpson | 0.76 | 0.75 | 0.73 | 0.61 | ||||
| Simpson_Inverse | 4.16 | 3.94 0.75 | 3.65 | 2.58 0.61 | ||||
| Simpson_Unbias | 0.76 | 0.73 | ||||||
| With sampling dates | ||||||||
| Shannon | 1.25 | (0.10) | 1.30 | (0.06) | 1.16 | (0.13) | 0.98 | (0.17) |
| Simpson | 0.63 | (0.05) | 0.65 | (0.03) | 0.59 | (0.05) | 0.49 | (0.09) |
| Simpson_Inverse | 2.96 | (0.35) | 3.03 | (0.28) | 2.84 | (0.47) | 2.38 | (0.38) |
| Simpson_Unbias | 0.64 | (0.05) | 0.67 | (0.03) | 0.63 | (0.05) | 0.52 | (0.09) |
| Superintensive | Traditional | |||||||
| Shannon | 1.73 | 1.50 | ||||||
| Simpson | 0.75 | 0.67 | ||||||
| Simpson_Inverse | 4.06 | 3.00 | ||||||
| Simpson_Unbias | 0.75 | 0.67 | ||||||
| With sampling dates | ||||||||
| Shannon | 1.34 | (0.08) | 1.17 | (0.12) | ||||
| Simpson | 0.65 | (0.03) | 0.56 | (0.06) | ||||
| Simpson_Inverse | 3.07 | (0.29) | 2.56 | (0.31) | ||||
| Simpson_Unbias | 0.66 | (0.03) | 0.58 | (0.06) | ||||
| Mineral | Compost | |||||||
| Shannon | 1.83 | 1.69 | ||||||
| Simpson | 0.77 | 0.72 | ||||||
| Simpson_Inverse | 4.30 | 3.59 | ||||||
| Simpson_Unbias | 0.77 | 0.72 | ||||||
| With sampling dates | ||||||||
| Shannon | 1.42 | (0.11) | 1.37 | (0.09) | ||||
| Simpson | 0.67 | (0.05) | 0.65 | (0.04) | ||||
| Simpson_Inverse | 3.40 | (0.42) | 3.18 | (0.41) | ||||
| Simpson_Unbias | 0.67 | (0.05) | 0.66 | (0.04) | ||||
References
- MAPA. Avances Superficie y Producción de Cultivos. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/superficies-producciones-anuales-cultivos/ (accessed on 23 May 2023).
- Morillo, J.A.; Antizar-Ladislao, B.; Monteoliva-Sánchez, M.; Ramos-Cormenzana, A.; Russell, N.J. Bioremediation and biovalorisation of olive-mill wastes. Appl. Microbiol. Biotechnol. 2009, 82, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Mamkagh, A.; AL-Zyoud, F.; Al-Atiyat, R. Olive Mill Pomace Compost as Soil Amendment, and Sources of Biopesticides and Animal Feed: A Review. Int. J. Agric. Biol. 2022, 28, 257–268. [Google Scholar] [CrossRef]
- Murrell, E.G. Can agricultural practices that mitigate or improve crop resilience to climate change also manage crop pests? Curr. Opin. Insect Sci. 2017, 23, 81–88. [Google Scholar] [CrossRef]
- Panettieri, M.; Moreno, B.; de Sosa, L.L.; Benítez, E.; Madejón, E. Soil management and compost amendment are the main drivers of carbon sequestration in rainfed olive trees agroecosystems: An evaluation of chemical and biological markers. Catena 2022, 214, 106258. [Google Scholar] [CrossRef]
- Junta de Andalucia Análisis de la Densidad en las Plantaciones de Olivar en Andalucia. Available online: https://www.juntadeandalucia.es/export/drupaljda/estudios_informes/19/11/Análisisdensidadolivarandaluzv3.pdf (accessed on 16 September 2023).
- Tribaldos Campos, J.; Tribaldos Campos, H. El Olivar en España: Tradicional, Intensivo y Superintensivo. Available online: https://www.agro.basf.es/es/Camposcopio/Secciones/Protección-y-sanidad/olivar-en-espana/ (accessed on 16 September 2023).
- MAPA. Anuarios 1999–2020. Available online: https://www.mapa.gob.es/es/estadistica/temas/publicaciones/anuario-de-estadistica/ (accessed on 24 May 2023).
- Baldovino Sanjuan, A.; Castellanos Gonzalez, L.; de Prado, R.M. Edaphic mesofauna, some studies done: A review. INGE CUC 2022, 18, 197–208. [Google Scholar] [CrossRef]
- Miller, J.J.; Battigelli, J.P.; Beasley, B.W.; Drury, C.F. Response of Soil Mesofauna to Long-Term Application of Feedlot Manure on Irrigated Cropland. J. Environ. Qual. 2017, 46, 185–192. [Google Scholar] [CrossRef]
- Rowen, E.; Tooker, J.F.; Blubaugh, C.K. Managing fertility with animal waste to promote arthropod pest suppression. Biol. Control 2019, 134, 130–140. [Google Scholar] [CrossRef]
- Goncharov, A.A.; Glebova, A.A.; Chernov, T.I.; Karpukhin, M.M.; Kuznetsova, N.A.; Leonov, V.D.; Makarova, O.L.; Maygurova, V.N.; Shesteperov, A.A.; Taranets, I.P.; et al. Detrital subsidy alters the soil invertebrate community and reduces infection of winter wheat seedlings by Fusarium wilt. Appl. Soil Ecol. 2021, 163, 103914. [Google Scholar] [CrossRef]
- Viketoft, M.; Riggi, L.G.A.; Bommarco, R.; Hallin, S.; Taylor, A.R. Type of organic fertilizer rather than organic amendment per se increases abundance of soil biota. PeerJ 2021, 9, e11204. [Google Scholar] [CrossRef]
- Gkisakis, V.D.; Volakakis, N.; Kollaros, D.; Bàrberi, P.; Kabourakis, E.M. Soil arthropod community in the olive agroecosystem: Determined by environment and farming practices in different management systems and agroecological zones. Agric. Ecosyst. Environ. 2016, 218, 178–189. [Google Scholar] [CrossRef]
- Carpio, A.J.; Castro, J.; Tortosa, F.S. Arthropod biodiversity in olive groves under two soil management systems: Presence versus absence of herbaceous cover crop. Agric. For. Entomol. 2018, 21, 58–68. [Google Scholar] [CrossRef]
- Álvarez, H.A.; Morente, M.; Campos, M.; Ruano, F. Maturity of ground covers increases the resilience and presence of natural enemies in the trophic network of olive canopy. Ecosistemas 2019, 28, 92–106. [Google Scholar] [CrossRef]
- Paredes, D.; Karp, D.S.; Chaplin-Kramer, R.; Benítez, E.; Campos, M. Natural habitat increases natural pest control in olive groves: Economic implications. J. Pest Sci. 2019, 92, 1111–1121. [Google Scholar] [CrossRef]
- Rey, P.J.; Manzaneda, A.J.; Valera, F.; Alcántara, J.M.; Tarifa, R.; Isla, J.; Molina-Pardo, J.L.; Calvo, G.; Salido, T.; Gutiérrez, J.E.; et al. Landscape-moderated biodiversity effects of ground herb cover in olive groves: Implications for regional biodiversity conservation. Agric. Ecosyst. Environ. 2019, 277, 61–73. [Google Scholar] [CrossRef]
- Picchi, M.S.; Marchi, S.; Albertini, A.; Petacchi, R. Organic management of olive orchards increases the predation rate of overwintering pupae of Bactrocera oleae (Diptera: Tephritidae). Biol. Control 2017, 108, 9–15. [Google Scholar] [CrossRef]
- Ortega, M.; Sánchez-Ramos, I.; González-Núñez, M.; Pascual, S. Time course study of Bactrocera oleae (Diptera: Tephritidae) pupae predation in soil: The effect of landscape structure and soil condition. Agric. For. Entomol. 2018, 20, 201–207. [Google Scholar] [CrossRef]
- Villa, M.; Santos, S.A.P.; Sousa, J.P.; Ferreira, A.; da Silva, P.M.; Patanita, I.; Ortega, M.; Pascual, S.; Pereira, J.A. Landscape composition and configuration affect the abundance of the olive moth (Prays oleae, Bernard) in olive groves. Agric. Ecosyst. Environ. 2020, 294, 106854. [Google Scholar] [CrossRef]
- Álvarez, H.A.; Jiménez-Muñoz, R.; Morente, M.; Campos, M.; Ruano, F. Ground cover presence in organic olive orchards affects the interaction of natural enemies against Prays oleae, promoting an effective egg predation. Agric. Ecosyst. Environ. 2021, 315, 107441. [Google Scholar] [CrossRef]
- Pascual, S.; Rescia, A.J.; Ondul, B.A.; Paul, C.; Ortega, M. Effects of landscape structure on abundance and family richness of hymenopteran parasitoids in the olive agroecosystem. Agric. Ecosyst. Environ. 2022, 332, 107914. [Google Scholar] [CrossRef]
- Pascual, S.; Ortega, M.; Villa, M. Prays oleae (Bernard), its potential predators and biocontrol depend on the structure of the surrounding landscape. Biol. Control 2022, 176, 105092. [Google Scholar] [CrossRef]
- Vasconcelos, S.; Pina, S.; Jonsson, M.; Heleno, R.; Herrera, J.M.; Moreira, F.; Silva, B.; Melguizo-Ruiz, N.; Daouti, E.; Jiménez-Navarro, G.; et al. Mixed effects of oak woodlands on biocontrol potential and pest pressure in olive orchards under management intensification. J. Pest Sci. 2023, 2023, 1–14. [Google Scholar] [CrossRef]
- Porcel, M.; Cotes, B.; Castro, J.; Campos, M. The effect of resident vegetation cover on abundance and diversity of green lacewings (Neuroptera: Chrysopidae) on olive trees. J. Pest Sci. 2017, 90, 195–206. [Google Scholar] [CrossRef]
- Alcala Herrera, R.; García-Fuentes, A.; Ramos-Font, M.E.; Fernández-Sierra, M.L.; Ruano, F. Vegetation Cover Management and Landscape Plant Species Composition Influence the Chrysopidae Community in the Olive Agroecosystem. Plants 2022, 11, 3255. [Google Scholar] [CrossRef]
- Way, M.; Khoo, K.C. Role Of Ants In Pest-Management. Annu. Rev. Entomol. 1992, 37, 479–503. [Google Scholar] [CrossRef]
- Morris, T.I.; Campos, M. Entomofauna depredadora del suelo del olivar. Zool. Baetica 1999, 10, 149–160. [Google Scholar]
- Santos, S.A.P.; Cabanas, J.E.; Pereira, J.A. Abundance and diversity of soil arthropods in olive grove ecosystem (Portugal): Effect of pitfall trap type. Eur. J. Soil Biol. 2007, 43, 77–83. [Google Scholar] [CrossRef]
- Santos, S.A.P.; Pereira, J.A.; Torres, L.M.; Nogueira, A.J.A. Evaluation of the effects, on canopy arthropods, of two agricultural management systems to control pests in olive groves from north-east of Portugal. Chemosphere 2007, 67, 131–139. [Google Scholar] [CrossRef]
- Ruiz-Torres, M.; Montiel Bueno, A. Introducción al conocimiento de la entomofauna del olivar en la provincia de Jaén. Aspectos cuantitativos (II). Boletín Sanid. Veg. Plagas 2001, 27, 531–560. [Google Scholar]
- Cotes, B.; Campos, M.; Pascual, F.; García, P.A.; Ruano, F. Comparing taxonomic levels of epigeal insects under different farming systems in Andalusian olive agroecosystems. Appl. Soil Ecol. 2010, 44, 228–236. [Google Scholar] [CrossRef]
- Ruiz-Torres, M.; Montiel Bueno, A. Introducción al conocimiento de la Entomofauna del olivar en la provincia de Jaén. Aspectos cualitativos. (I). Boletín Sanid. Veg. Plagas 2000, 26, 129–147. [Google Scholar]
- Morris, T.I.; Symondson, W.O.C.; Kidd, N.A.C.; Jervis, M.A.; Campos, M. Are ants significant predators of the olive moth, Prays oleae? Crop Prot. 1998, 17, 365–366. [Google Scholar] [CrossRef]
- Morris, T.I.; Symondson, W.O.C.; Kidd, N.A.C.; Campos, M. The effect of different ant species on the olive moth, Prays oleae (Bern.), in Spanish olive orchard. J. Appl. Entomol. 2002, 126, 224–230. [Google Scholar] [CrossRef]
- Martínez-Núñez, C.; Rey, P.J.; Salido, T.; Manzaneda, A.J.; Camacho, F.M.; Isla, J. Ant community potential for pest control in olive groves: Management and landscape effects. Agric. Ecosyst. Environ. 2021, 305, 107185. [Google Scholar] [CrossRef]
- Campos, M.; Fernández, L.; Ruano, F.; Cotes, B.; Cárdenas, M.; Castro, J. Short term response of ants to the removal of ground cover in organic olive orchards. Eur. J. Entomol. 2011, 108, 417–423. [Google Scholar] [CrossRef]
- Galindo, A.; Collado-González, J.; Griñán, I.; Corell, M.; Centeno, A.; Martín-Palomo, M.J.; Girón, I.F.; Rodríguez, P.; Cruz, Z.N.; Memmi, H.; et al. Deficit irrigation and emerging fruit crops as a strategy to save water in Mediterranean semiarid agrosystems. Agric. Water Manag. 2018, 202, 311–324. [Google Scholar] [CrossRef]
- Southwood, T.R.E.; Henderson, P.A. Ecological Methods, 3rd ed.; Blackwell Science Ltd.: Malden, MA, USA, 2000; ISBN 0-632-05477-8. [Google Scholar]
- Gonçalves, M.F.; Pereira, J.A. Abundance and Diversity of Soil Arthropods in the Olive Grove Ecosystem. J. Insect Sci. 2012, 12, 20. [Google Scholar] [CrossRef]
- Barrientos, A.J. (Ed.) Curso Práctico de Entomología; Manuals de la Universitat Autònoma de Barcelona; 41; Asociación Española de Entomología; CIBIO; Universidad Autónoma de Barcelona: Alicante and Bellaterra, Spain, 2004; ISBN 8449023831. [Google Scholar]
- Chinery, M. Guía de Campo de los Insectos de España y de Europa; Omega: Barcelona, Spain, 2005; ISBN 84-282-0469-1. [Google Scholar]
- Lebas, C.; Galkowski, C.; Blatrix, R.; Wegnez, P. Guía de Campo de las Hormigas de Europa Occidental; Omega: Barcelona, Spain, 2017; ISBN 9788428216630. [Google Scholar]
- Sharley, D.J.; Hoffmann, A.A.; Thomson, L.J. The effects of soil tillage on beneficial invertebrates within the vineyard. Agric. For. Entomol. 2008, 10, 233–243. [Google Scholar] [CrossRef]
- Álvarez, H.A.; García-García, A.; Sandoval, P.; Martín-Blázquez, R.; Seifert, B.; Tinaut, A.; Ruano, F. Elucidating the trophic role of Tapinoma ibericum (Hymenoptera: Formicidae) as a potential predator of olive pests. J. Appl. Entomol. 2023, 147, 667–675. [Google Scholar] [CrossRef]
- Alvarado, M.; Durán, J.M.; González, M.I.; Montes, F.; Páez, J.; Sánchez, A.; Serrano, A.; Vega, J.M. Plagas y Enfermedades del Olivo; Junta de Andalucía, Consejería de Agricultura y Pesca: Sevilla, Spain, 2002. [Google Scholar]
- Mansour, R.; Suma, P.; Mazzeo, G.; La Pergola, A.; Pappalardo, V.; Grissa Lebdi, K.; Russo, A. Interactions between the ant Tapinoma nigerrimum (Hymenoptera: Formicidae) and the main natural enemies of the vine and citrus mealybugs (Hemiptera: Pseudococcidae). Biocontrol Sci. Technol. 2012, 22, 527–537. [Google Scholar] [CrossRef]
- Detrain, C. Field study on foraging by the polymorphic ant species, Pheidole pallidula. Insectes Soc. 1990, 37, 315–332. [Google Scholar] [CrossRef]
- Serrano, J.M.; Acosta, F.J.; Alvarez, M. Estructura de las comunidades de hormigas en eriales mediterráneos según criterios funcionales. Graellsia 1987, 43, 211–223. [Google Scholar]



| 2021 a | 2022 a | 2021–2022 b | ||||||
|---|---|---|---|---|---|---|---|---|
| Superintensive | Traditional | Superintensive | Traditional | Estimate | SE | p | ||
| Sweep net | Sweep net | |||||||
| Treatment (Tr) | p = 0.437 [F = 0.74; 1, 4] | - c | p = 0.106 [F = 4.3; 1, 4] | p = 1 [F = 0; 1, 6] | Fertilization: Mineral/Compost | 0.31 | 0.38 | 0.415 |
| Sampling date (Sd) | p < 0.01 ** [F = 12.1; 2, 8.2] | - | p < 0.01 ** [F = 15.3; 1.9, 7.4] | p = 0.063 [F = 3.6; 1.9, 11.5] | Grove: Traditional/Superint. | −2.52 | 0.44 | <0.01 ** |
| Tr × Sd | p = 0.335 [F = 1.3; 2, 8.2] | - | p = 0.191 [F = 2.1; 1.9, 7.4] | p = 0.076 [F = 3.3; 1.9, 11.5] | Fertilization × Grove | −0.31 | 0.62 | 0.618 |
| Pitfall | Pitfall | |||||||
| Treatment (Tr) | p = 0.371 [F = 1.01; 1, 4] | p = 0.559 [F = 0.383; 1, 6] | p = 0.99 [F = 0; 1, 4] | p = 0.561 [F = 0.38; 1, 6] | Fertilization: Mineral/Compost | −0.15 | 0.18 | 0.416 |
| Sampling date (Sd) | p = 0.023 * [F = 9.4; 1.3, 5.3] | p = 0.586 [F = 0.42; 1.3, 7.7] | p = 0.243 [F = 1.7; 1.8, 7.2] | p = 0.154 [F = 2.2; 2.1, 12.6] | Grove: Traditional/Superint. | −1.27 | 0.18 | <0.01 ** |
| Tr × Sd | p = 0.150 [F = 2.8; 1.3, 5.3] | p = 0.478 [F = 0.66; 1.3, 7.7] | p = 0.624 [F = 0.47; 1.8, 7.2] | p = 0.868 [F = 0.15; 2.1, 12.6] | Fertilization × Grove | 0.14 | 0.25 | 0.580 |
| Subfamily | Species | Superintensive | Traditional | Total |
|---|---|---|---|---|
| Dolichoderinae | ||||
| Tapinoma nigerrimun | 27 (10.7) | 6 (24.0) (*) | 33 (11.9) | |
| Formicinae | ||||
| Plagiolepis spp. | 219 (86.6) (**) | 11 (44.0) | 230 (82.7) | |
| Plagiolepis pygmaea | 85 (33.6) | 10 (40.0) | 95 (34.2) | |
| Plagiolepis smitzii | 134 (53.0) (**) | 1 (4.0) | 135 (48.6) | |
| Myrmicinae | ||||
| Crematogaster sp. | 7 (2.8) | 2 (8.0) | 9 (3.2) | |
| Pheidole pallidula | 0 | 5 (20.0) (**) | 5 (1.8) | |
| Tetramorium gr semilaeve | 0 | 1 (4.0) | 1 (0.4) | |
| Total numbers | 253 | 25 | 278 | |
| Superint. | Superint. | Traditional | Traditional | Superintensive | Traditional | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Subfamily | Species | Mineral | Compost | Mineral | Compost | Mineral | Compost | TOTAL | ||
| Dolichoderinae | ||||||||||
| Tapinoma nigerrimun | 46 | 28 | 13 | 18 | 74 (7.5) | 31 (7.9) | 59 | 46 | 105 (7.6) | |
| Formicinae | ||||||||||
| Camponotus sp. | 2 | 0 | 2 | 1 | 2 (0.2) | 3 (0.8) | 4 | 1 | 5 (0.4) | |
| Cataglyphis velox | 5 | 1 | 13 | 4 | 6 (0.6) | 17(4.3) | 18 | 5 | 23 (1.7) | |
| Plagiolepis spp. | 151 | 122 | 11 | 21 | 273 (27.7) (**) | 32 (8.2) | 162 | 143 | 305 (22.1) | |
| Plagiolepis pygmaea | 49 | 36 | 11 | 21 | 85 (8.6) | 32 (8.2) | 60 | 57 | 117 (8.5) | |
| Plagiolepis smitzii | 102 | 86 | 0 | 0 | 188 (19.1) (**) | 0 | 102 | 86 | 188 (13.7) | |
| Myrmicinae | ||||||||||
| Aphaenogaster senilis | 51 | 59 | 11 | 6 | 110 (11.2) | 17 (4.3) | 62 | 65 | 127 (9.2) | |
| Crematogaster sp. | 1 | 3 | 0 | 2 | 4 (0.4) | 2 (0.5) | 1 | 5 | 6 (0.4) | |
| Messor spp. | 41 | 30 | 4 | 5 | 71 (7.2) | 9 (2.3) | 45 | 35 | 80 (5.8) | |
| Messor gr bouveri | 11 | 7 | 1 | 5 | 18 (1.8) | 6 (1.5) | 12 | 12 | 24 (1.7) | |
| Messor gr structor | 0 | 0 | 2 | 0 | 0 | 2 (0.5) | 2 | 0 | 2 (0.2) | |
| Messor barbarus | 30 | 23 | 1 | 0 | 53 (5.4) (*) | 1 (0.3) | 31 | 23 | 54 (3.9) | |
| Pheidole pallidula | 221 | 197 | 74 | 134 | 418 (42.4) | 208 (53.1) | 295 | 331 | 626 (45.5) | |
| Tetramorium gr semilaeve | 12 | 15 | 36 | 37 | 27 (2.7) | 73 (18.6) (**) | 48 | 52 | 100 (7.3) | |
| TOTAL NUMBERS | 530 | 455 | 164 | 228 | 985 | 392 | 694 | 683 | 1377 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Zamora, J.E.; Gamero-Monge, J.M.; Pérez-de la Luz, R. Ant Community Is Not Influenced by the Addition of Olive Mill Pomace Compost in Two Different Olive Crop Managements. Insects 2023, 14, 783. https://doi.org/10.3390/insects14100783
González-Zamora JE, Gamero-Monge JM, Pérez-de la Luz R. Ant Community Is Not Influenced by the Addition of Olive Mill Pomace Compost in Two Different Olive Crop Managements. Insects. 2023; 14(10):783. https://doi.org/10.3390/insects14100783
Chicago/Turabian StyleGonzález-Zamora, José E., José M. Gamero-Monge, and Rosa Pérez-de la Luz. 2023. "Ant Community Is Not Influenced by the Addition of Olive Mill Pomace Compost in Two Different Olive Crop Managements" Insects 14, no. 10: 783. https://doi.org/10.3390/insects14100783