
Effects of Different Parts of the Rose Flower on the Development, Fecundity, and Life Parameters of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Source and Foods
2.2. The Group-Rearing of WFT on Different Foods
2.3. Development Duration of WFT
2.4. The Fecundity of WFT
2.5. Data Analysis
3. Results
3.1. Effects of Different Food Types on the Development of WFT in Successive Generations
3.2. Effects of Different Foods on WFT Longevity and Reproduction in Successive Generations
3.3. Effects of Different Foods on WFT Survival in Successive Generations
3.4. Effects of Different Foods on the Population Parameters of Successive Generations of WFT
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Topal, E.; Mărgăoan, R.; Bay, V.; Takma, C.; Yücel, B.; Oskay, D.; Düz, G.; Acar, S.; Kösoğlu, M. The effect of supplementary feeding with different pollens in autumn on colony development under natural environment and in vitro lifespan of honey bees. Insects 2022, 13, 588. [Google Scholar] [CrossRef]
- Vengateswari, G.; Arunthirumeni, M.; Shivaswamy, M.S.; Takma, C.; Yücel, B.; Oskay, D.; Düz, G.; Acar, S.; Kösoğlu, M. Effect of host plants nutrients, antioxidants, and phytochemicals on growth, development, and fecundity of Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae). Int. J. Trop. Insect Sci. 2022, 42, 3161–3173. [Google Scholar] [CrossRef]
- Varikou, K.N.; Tsitsipis, I.; Alexandrakis, V. Effect of diet on development and reproduction of Pezothrips kellyanus (Thysanoptera: Thripidae). Ann. Entomol. Soc. Am. 2010, 103, 66–70. [Google Scholar] [CrossRef]
- da SilvaI, F.; Baldin, E.L.L.; Specht, A.; Roque-Specht, V.F.; Morando, R.; Malaquias, J.V.; Paula-Moraes, S.V. Role of nutritional composition in the development and survival of Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on artificial diet and natural hosts. Bull. Entomol. Res. 2021, 111, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Wen, D.M.; Liu, Y.H.; Ren, L.L.; Lu, P.F.; Luo, Y.Q. The effect of host plant species on the development of Trabala vishnou gigantina Yang (Lepidoptera: Lasiocampidae). Chin. J. Appl. Entomol. 2016, 53, 157–163. [Google Scholar]
- Gao, Z.; Pan, H.S.; Liu, B.; Lu, Y.H.; Liang, G.M. Performance of three Adelphocoris spp. (Hemiptera: Miridae) on flowering and non-flowering cotton and alfalfa. J. Integr. Agric. 2014, 13, 1727–1735. [Google Scholar] [CrossRef]
- Angelella, G.M.; Riley, D.G. Effects of pine pollen supplementation in an onion diet on Frankliniella fusca reproduction. Environ. Entomol. 2010, 39, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.L.; Li, Q.; Qiu, H.Y.; Tang, L.D.; Zeng, D.Q.; Liu, K.; Gao, Y.L. Oviposition, feeding preference, and biological performance of Thrips hawaiiensis on four host plants with and without supplemental foods. Arthropod-Plant Interact. 2019, 13, 441–452. [Google Scholar] [CrossRef]
- Kirk, W.D.J.; Terry, L.I. The spread of the western flower thrips Frankliniella occidentalis (Pergande). Agr. Forest Entomol. 2003, 5, 301–310. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.L.; Kirk, W.D.J.; Hoddle, M.S.; Leiss, K.A.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.B.; Zhang, Z.J.; Wu, Q.J.; Du, Y.Z.; Zhang, H.R.; Yu, Y.; Wang, E.D.; Wang, M.H.; Wang, M.Q.; Tong, X.L.; et al. Research progress of the monitoring, forecast and sustainable management of invasive alien pest Frankliniella occidentalis in China. Chin. J. Appl. Entomol. 2011, 48, 488–496. [Google Scholar]
- Wu, S.Y.; Tang, L.D.; Zhang, X.R.; Xing, Z.L.; Lei, Z.R.; Gao, Y.L. A decade of a thrips invasion in China: Lessons learned. Ecotoxicology 2018, 27, 1032–1038. [Google Scholar] [CrossRef]
- Zhang, B.; Qian, W.Q.; Qiao, X.; Xi, Y.; Wan, F.H. Invasion biology, ecology, and management of Frankliniella occidentalis in China. Arch. Insect Biochem. Physiol. 2019, 102, e21613. [Google Scholar] [CrossRef]
- Morse, J.G.; Hoddle, M.S. Invasion biology of thrips. Annu. Rev. Entomol. 2006, 51, 67–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arthurs, S.P.; Heinz, K.M.; Mitchell, F.L. Comparison of Frankliniella fusca and Frankliniella occidentalis (Thysanoptera: Thripidae) as vectors for a peanut strain of Tomato spotted wilt orthotospovirus. Environ. Entomol. 2018, 47, 623–628. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Guo, J.F.; Reitz, S.R.; Lei, Z.R.; Wu, S.Y. A global invasion by the thrips, Frankliniella occidentalis: Current virus vector status and its management. Insect Sci. 2019, 27, 626–645. [Google Scholar] [CrossRef] [PubMed]
- Kirk, W.D.J. Pollen-feeding and the host specificity and fecundity of flower thrips (Thysanoptera). Ecol. Entomol. 1985, 10, 281–289. [Google Scholar] [CrossRef]
- Ren, X.Y.; Wu, S.Y.; Xing, Z.L.; Gao, Y.L.; Cai, W.Z.; Lei, Z.R. Abundances of thrips on plants in vegetative and flowering stages are related to plant volatiles. J. Appl. Entomol. 2020, 144, 732–742. [Google Scholar] [CrossRef]
- Gerin, C.; Hance, T.H.; Van Impe, G. Impact of flowers on the demography of western flower thrips Frankliniella occidentalis (Thysan, Thripidae). J. Appl. Entomol. 1999, 123, 569–574. [Google Scholar] [CrossRef]
- Hulshof, J.; Ketoja, E.; Vänninen, I. Life history characteristics of Frankliniella occidentalis on cucumber leaves with and without supplemental food. Entomol. Exp. Appl. 2003, 108, 19–32. [Google Scholar] [CrossRef]
- Zhi, J.R.; Fitch, G.K.; Margolies, D.C.; Nechols, J.R. Apple pollen as a supplemental food for the western flower thrips, Frankliniella occidentalis: Response of individuals and populations. Entomol. Exp. Appl. 2005, 117, 185–192. [Google Scholar] [CrossRef]
- Cao, Y.; Li, C.; Yang, H.; Li, J.; Li, S.; Wang, Y.W.; Gao, Y.L. Laboratory and field investigation on the orientation of Frankliniella occidentalis (Thysanoptera: Thripidae) to more suitable host plants driven by volatiles and component analysis of volatiles. Pest Manag. Sci. 2019, 57, 598–606. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhi, J.R.; Zhang, R.Z.; Li, C.; Liu, Y.; Lv, Z.Y.; Gao, Y.L. Different population performances of Frankliniella occidentalis and Thrips hawaiiensis on flowers of two horticultural plants. J. Pest Sci. 2018, 91, 79–91. [Google Scholar] [CrossRef]
- Cao, Y.; Yang, H.; Li, J.; Zhang, G.Z.; Wang, Y.W.; Li, C.; Gao, Y.L. Population development of Frankliniella occidentalis and Thrips hawaiiensis in constant and fluctuating temperatures. J. Appl. Entomol. 2019, 143, 49–57. [Google Scholar] [CrossRef] [Green Version]
- Park, J.D.; Kim, S.G.; Kim, D.I.; Cho, K. Population dynamics of Frankliniella occidentalis on different rose cultivars and flowering stages. J. Asia-Pac. Entomol. 2002, 5, 97–102. [Google Scholar] [CrossRef]
- Sun, M.; Zhi, J.R.; Yao, J.J.; Ma, C. Population dynamics of Frankliniella occidentalis on cut rose. Guizhou Agric. Sci. 2011, 39, 83–86. [Google Scholar]
- Shen, D.R.; Zhang, H.R.; Li, Z.Y.; He, S.Y. Effects of different foods on the growth and development of Frankliniella occidentalis. Plant Prot. 2012, 38, 55–59. [Google Scholar]
- Qi, G.L.; Gu, Z.Y.; Shen, J.W.; Wang, C.; Zeng, L.; Reitz, S.R.; Cao, Y. Rosa chinensis cultivars affect fitness-related characteristics and digestive physiology of the western flower thrips, Frankliniella occidentalis Pergande (Thysanoptera: Thripidae). J. Econ. Entomol. 2022, 115, 1620–1626. [Google Scholar] [CrossRef] [PubMed]
- Watts, J.G. Comparison of the life cycles of Franklimiella tritici (Fitch), F. fusa(Hind) and Thrips Tabaci Lind. (Tysanoptera -Thripidaein) in South Carolina. J. Econ. Entomol. 1934, 27, 1158–1159. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2021. Available online: http://140.120.197.173/Ecology/Download/Twosex-MSChart-B100000.rar (accessed on 3 February 2021).
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2016, 35, 10–21. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef] [PubMed]
- Polat, A.E.; Atlıhan, R.; Okut, H.; Chi, H. Demographic assessment of plant cultivar resistance to insect pests: A case study of the dusky-veined walnut aphid (Hemiptera: Callaphididae) on five walnut cultivars. J. Econ. Entomol. 2015, 108, 378–387. [Google Scholar]
- Cao, Y.; Liu, Y.; Liang, W.Q.; Wang, C.; Li, C. Development of Frankliniella occidentalis on flowers of different horticultural hosts and relationship with flower compounds. Chin. J. Eco-Agric. 2015, 23, 634–641. [Google Scholar]
- Gou, Y.P.; Quandahor, P.; Zhang, Y.Z.; Coulter, J.A.; Liu, C.Z. Host plant nutrient contents influence nutrient contents in Bradysia cellarum and Bradysia impatiens. PLoS ONE 2015, 15, e0226471. [Google Scholar] [CrossRef]
- Sun, M. The Population Dynamics of Frankliniella occidentalis on Cut Rose and Its Control Techniques. Master Thesis, Guizhou University, Guiyang, China, 2011. [Google Scholar]
- Zhi, J.R.; Liu, L.; Hou, X.L.; Xie, W.; Yue, W.B.; Zeng, G. Role of digestive enzymes in the adaptation of Frankliniella occidentalis to preferred and less-preferred host plants. Entomol. Exp. Appl. 2021, 169, 688–700. [Google Scholar] [CrossRef]
- Trichilo, P.J.; Leigh, T.F. Influence of resource quality on the reproductive fitness of flower thrips (Thysanoptera: Thripidae). Ann. Entomol. Soc. Am. 1988, 81, 64–70. [Google Scholar] [CrossRef]
- Zhu, P.Y.; Sheng, X.Q.; Fang, D.H.; Chen, G.H.; Heong, K.L.; Lv, Z.X. Effect of feeding parental adults with plant flowers on growth and predatory capacity of next generation of mired bug Cyrtorhinus lividipennis. Chin. Plant Prot. 2013, 33, 17–21. [Google Scholar]
- Li, Y.R.; Li, Z.; Yang, Y.H.; Yang, Q.P.; Liu, X.X.; Zhang, Q.W. Impact of food diversity on biological parameters of Apolygus lucorum (Hemiptera: Heteroptera: Miridae). Fla. Entomol. 2015, 98, 1188–1192. [Google Scholar] [CrossRef] [Green Version]
- Riley, D.G.; Angelella, G.M.; Mcpherson, R.M. Pine pollen dehiscence relative to thrips population dynamics. Entomol. Exp. Appl. 2011, 138, 223–233. [Google Scholar] [CrossRef]
- Li, Q.; Liu, K.; Fu, B.L.; Zeng, Q.D.; Xia, X.Y.; Qiu, H.Y.; Tang, L.D. Effects of different diets on the biological characteristics of the Thrips hawaiiensis. J. Environ. Entomol. 2018, 40, 136–143. [Google Scholar]
- Wu, F.A.; Zhou, J.X.; Yu, M.D.; Wang, Q.L.; Xu, L.; Lu, C.; Jin, C.J. Statistical inference on the intrinsic rate of increase of the carmine spider mite, Tetranychus cinnabarinus on different mulberry cultivars (Morus L.) under laboratory conditions. Acta Entomol. Sin. 2006, 49, 287–294. [Google Scholar]
- Zhao, M.; Ju, R.T. Effects of temperature on the development and fecundity of experimental population of Rhychophorus ferrugineus. Plant Prot. 2010, 37, 517–521. [Google Scholar]
- Dhall, H.; Jangra, S.; Basavaraj, Y.B.; Ghosh, A. Host plant influences life cycle, reproduction, feeding, and vector competence of Thrips palmi (Thysanoptera: Thripidae), a vector of tospoviruses. Phytoparasitica 2021, 49, 501–512. [Google Scholar] [CrossRef]
- Wäckers, F.L.; Romeis, J.; Van, R.P. Nectar and pollen feeding by insect herbivores and implications for multitrophic interactions. Annu. Rev. Entomol. 2007, 52, 301–323. [Google Scholar] [CrossRef] [Green Version]
- Van, R.P.C.J.; Tanigoshi, L.K. The contribution of extrafloral nectar to survival and reproduction of the predatory mite Iphiseius degenerans on Ricinus communis. Exp. Appl. Acarol. 1999, 23, 281–296. [Google Scholar]
- Zhang, W.N.; Liu, B.; Liang, G.M.; Lu, Y.H. Flowers promote ovarian development and vitellogenin gene expression in Apolygus lucorum (Heteroptera: Miridae). Arthropod-Plant Interact. 2016, 10, 113–119. [Google Scholar] [CrossRef]
- Zhou, D. Effects of Different Foods on Life Table Parameters and Digestive Enzyme Activities of Frankliniella occidentalis (Thysanoptera: Thripidae). Master Thesis, Guizhou University, Guiyang, China, 2021. [Google Scholar]
- van Rijn, P.C.J.; Mollema, C.; Steenhuis-Broers, G.M. Comparative life history studies of Frankliniella occidentalis and Thrips tabaci (Thysanoptera: Thripidae) on cucumber. Bull. Entomol. Res. 1995, 85, 285–297. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
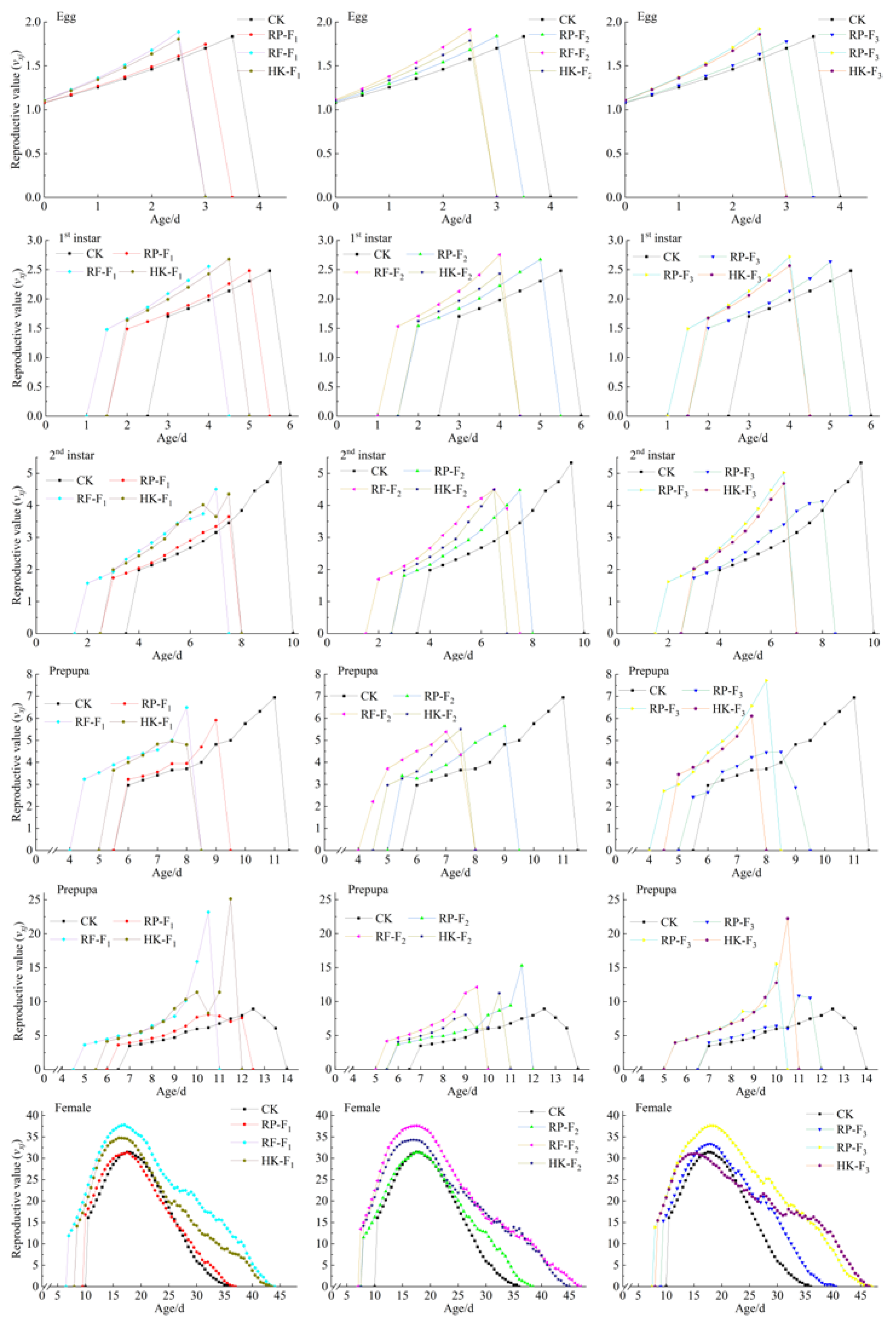{kind=link}
| Stage | Generation | Rose Petals | Rose Flowers | 10% Honey Solution + Kidney Bean Pods |
|---|---|---|---|---|
| Egg (d) | Control | 3.58 ± 0.04 a | 3.58 ± 0.04 a | 3.58 ± 0.04 a |
| F1 | 2.77 ± 0.05 bA | 2.38 ± 0.05 bC | 2.54 ± 0.04 bB | |
| F2 | 2.68 ± 0.06 bA | 2.24 ± 0.05 bC | 2.43 ± 0.04 bB | |
| F3 | 2.71 ± 0.06 bA | 2.27 ± 0.06 bC | 2.46 ± 0.04 bB | |
| 1st instar (d) | Control | 1.52 ± 0.04 ab | 1.52 ± 0.04 ab | 1.52 ± 0.04 ab |
| F1 | 1.57 ± 0.03 aA | 1.21 ± 0.05 bC | 1.37 ± 0.04 bB | |
| F2 | 1.49 ± 0.04 abA | 1.12 ± 0.05 bC | 1.23 ± 0.03 cB | |
| F3 | 1.46 ± 0.04 bA | 1.15 ± 0.05 bB | 1.25 ± 0.03 cB | |
| 2nd instar (d) | Control | 3.30 ± 0.05 a | 3.30 ± 0.05 a | 3.30 ± 0.05 a |
| F1 | 2.77 ± 0.06 bA | 2.52 ± 0.05 bB | 2.60 ± 0.05 bB | |
| F2 | 2.68 ± 0.06 bA | 2.43 ± 0.04 bB | 2.49 ± 0.04 bB | |
| F3 | 2.70 ± 0.06 bA | 2.45 ± 0.05 bB | 2.48 ± 0.05 bB | |
| Prepupa (d) | Control | 1.30 ± 0.05 a | 1.30 ± 0.05 a | 1.30 ± 0.05 a |
| F1 | 1.03 ± 0.05 bA | 0.91 ± 0.05 bA | 0.93 ± 0.04 bA | |
| F2 | 0.98 ± 0.05 bA | 0.87 ± 0.05 bA | 0.89 ± 0.04 bA | |
| F3 | 1.02 ± 0.05 bA | 0.90 ± 0.05 bA | 0.90 ± 0.04 bA | |
| Pupa (d) | Control | 2.92 ± 0.06 a | 2.92 ± 0.06 a | 2.92 ± 0.06 a |
| F1 | 2.72 ± 0.06 bA | 2.39 ± 0.07 bB | 2.51 ± 0.07 bB | |
| F2 | 2.65 ± 0.06 bA | 2.24 ± 0.06 bC | 2.40 ± 0.05 bB | |
| F3 | 2.68 ± 0.07 bA | 2.27 ± 0.05 bC | 2.46 ± 0.05 bB | |
| Immature (d) | Control | 12.64 ± 0.15 a | 12.64 ± 0.15 a | 12.64 ± 0.15 a |
| F1 | 10.87 ± 0.12 bA | 9.39 ± 0.12 bC | 9.92 ± 0.11 bB | |
| F2 | 10.54 ± 0.12 bcA | 8.86 ± 0.09 cC | 9.46 ± 0.11 cB | |
| F3 | 10.54 ± 0.10 cA | 9.09 ± 0.10b cC | 9.55 ± 0.09 cB |
| Fecundity | Generation | Rose Petals | Rose Flowers | 10% Honey Solution + Kidney Bean Pods |
|---|---|---|---|---|
| Longevity of Female (d) | Control | 21.80 ± 0.44 b | 21.80 ± 0.44 b | 21.80 ± 0.44 b |
| F1 | 23.47 ± 0.89 abB | 29.08 ± 1.36 aA | 26.37 ± 1.41 aAB | |
| F2 | 25.78 ± 0.85 aB | 31.16 ± 1.38 aA | 28.92 ± 1.47 aAB | |
| F3 | 25.55 ± 1.29 aB | 31.22 ± 1.40 aA | 29.12 ± 1.74 aAB | |
| Longevity of Male (d) | Control | 11.28 ± 0.16 a | 11.28 ± 0.16 b | 11.28 ± 0.16 b |
| F1 | 11.75 ± 0.47 aB | 13.43 ± 0.19 aA | 12.33 ± 0.46 aAB | |
| F2 | 12.11 ± 0.44 aB | 13.77 ± 0.51 aA | 13.21 ± 0.50 aAB | |
| F3 | 12.04 ± 0.49 aB | 13.71 ± 0.48 aA | 13.11 ± 0.48 aAB | |
| Eggs/female | Control | 80.84 ± 1.40 c | 80.84 ± 1.40 b | 80.84 ± 1.40 b |
| F1 | 86.89 ± 1.89 bC | 155.69 ± 7.00 aA | 124.82 ± 5.47 aB | |
| F2 | 99.08 ± 2.55 aC | 163.17 ± 6.53 aA | 135.80 ± 5.22 aB | |
| F3 | 101.24 ± 5.01 aC | 168.27 ± 7.03 aA | 138.37 ± 7.96 aB |
| Life Table Parameters | Generation | Rose Petals | Rose Flowers | 10% Honey Solution + Kidney Bean Pods |
|---|---|---|---|---|
| Net reproductive rate (R0) | Control | 26.95 ± 4.94 a | 26.95 ± 4.94 b | 26.95 ± 4.94 b |
| F1 | 27.51 ± 5.25 aB | 62.21 ± 10.23 aA | 51.99 ± 8.28 aA | |
| F2 | 37.96 ± 6.29 aB | 67.96 ± 10.73 aA | 49.83 ± 8.67 aAB | |
| F3 | 33.75 ± 6.36 aB | 72.97 ± 11.23 aA | 57.64 ± 9.38 aA | |
| Intrinsic rate of increase (r) | Control | 0.1510 ± 0.0095 a | 0.1510 ± 0.0095 b | 0.1510 ± 0.0095 b |
| F1 | 0.1581 ± 0.0105 aB | 0.2052 ± 0.0100 aA | 0.1964 ± 0.0090 aA | |
| F2 | 0.1724 ± 0.0092 aB | 0.2138 ± 0.0097 aA | 0.1929 ± 0.0103 aAB | |
| F3 | 0.1624 ± 0.0098 aB | 0.2100 ± 0.0091 aA | 0.2051 ± 0.0098 aA | |
| Finite rate of increase (λ) | Control | 1.1630 ± 0.0111 a | 1.1630 ± 0.0111 b | 1.1630 ± 0.0111 b |
| F1 | 1.1714 ± 0.0123 aB | 1.2278 ± 0.0123 aA | 1.2171 ± 0.0110 aA | |
| F2 | 1.1882 ± 0.0110 aB | 1.2385 ± 0.0120 aA | 1.2129 ± 0.0125 aAB | |
| F3 | 1.1764 ± 0.0116 aB | 1.2338 ± 0.0112 aA | 1.2277 ± 0.0120 aA | |
| Mean generation time (T)/d | Control | 21.71 ± 0.33 a | 21.71 ± 0.33 a | 21.71 ± 0.33 a |
| F1 | 20.85 ± 0.33 aA | 20.08 ± 0.39 bA | 20.05 ± 0.26 bA | |
| F2 | 21.02 ± 0.36 aA | 19.68 ± 0.34 bB | 20.19 ± 0.37 bAB | |
| F3 | 21.56 ± 0.37 aA | 20.38 ± 0.36 bB | 19.71 ± 0.36 bB |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.-Y.; Zhou, D.; Zhi, J.-R.; Yue, W.-B.; Li, S.-X. Effects of Different Parts of the Rose Flower on the Development, Fecundity, and Life Parameters of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Insects 2023, 14, 88. https://doi.org/10.3390/insects14010088
Li D-Y, Zhou D, Zhi J-R, Yue W-B, Li S-X. Effects of Different Parts of the Rose Flower on the Development, Fecundity, and Life Parameters of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Insects. 2023; 14(1):88. https://doi.org/10.3390/insects14010088
Chicago/Turabian StyleLi, Ding-Yin, Dan Zhou, Jun-Rui Zhi, Wen-Bo Yue, and Shun-Xin Li. 2023. "Effects of Different Parts of the Rose Flower on the Development, Fecundity, and Life Parameters of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae)" Insects 14, no. 1: 88. https://doi.org/10.3390/insects14010088