
Presence of Spodoptera frugiperda Multiple Nucleopolyhedrovirus (SfMNPV) Occlusion Bodies in Maize Field Soils of Mesoamerica
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Colonies and Reference Virus Isolate
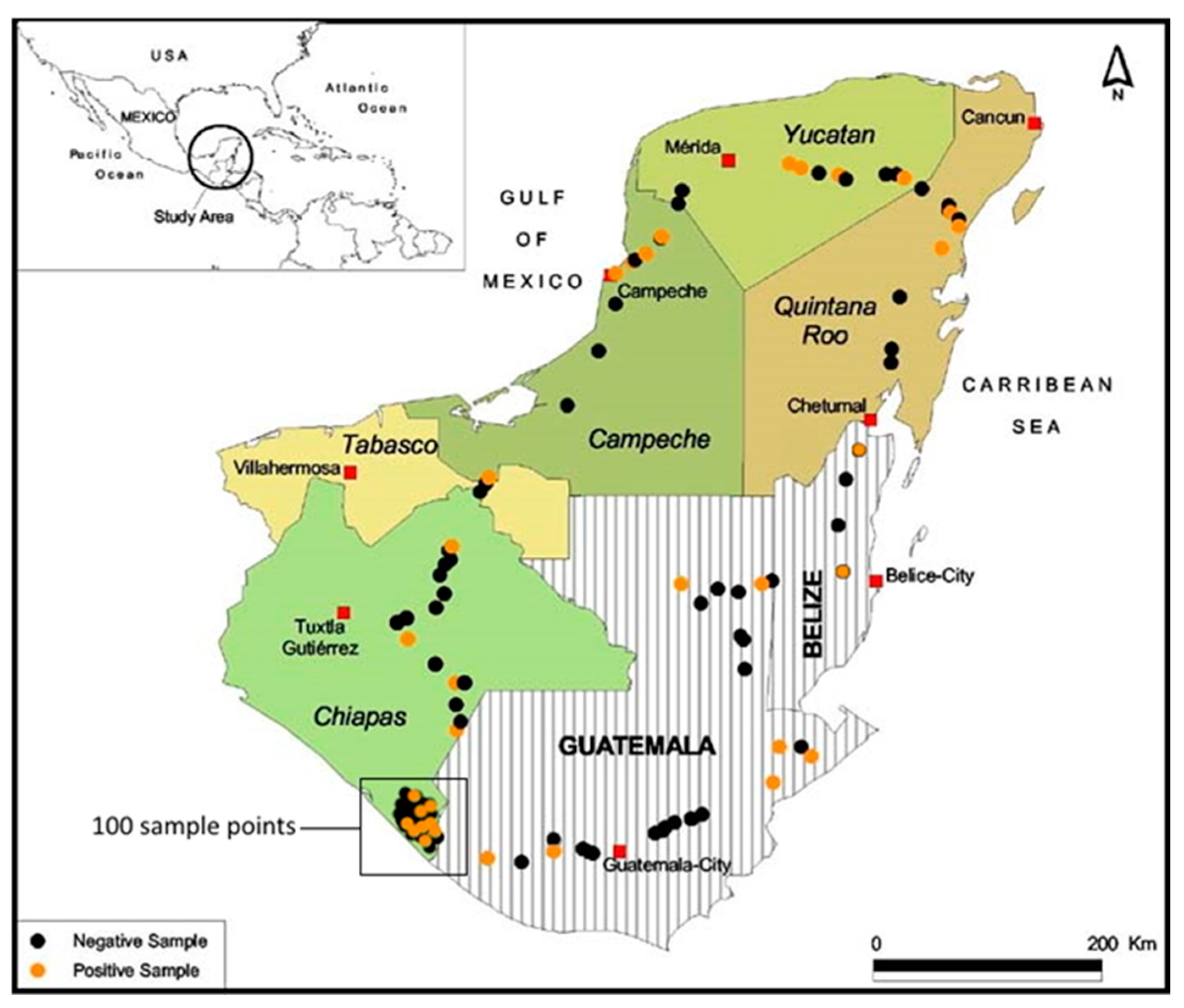
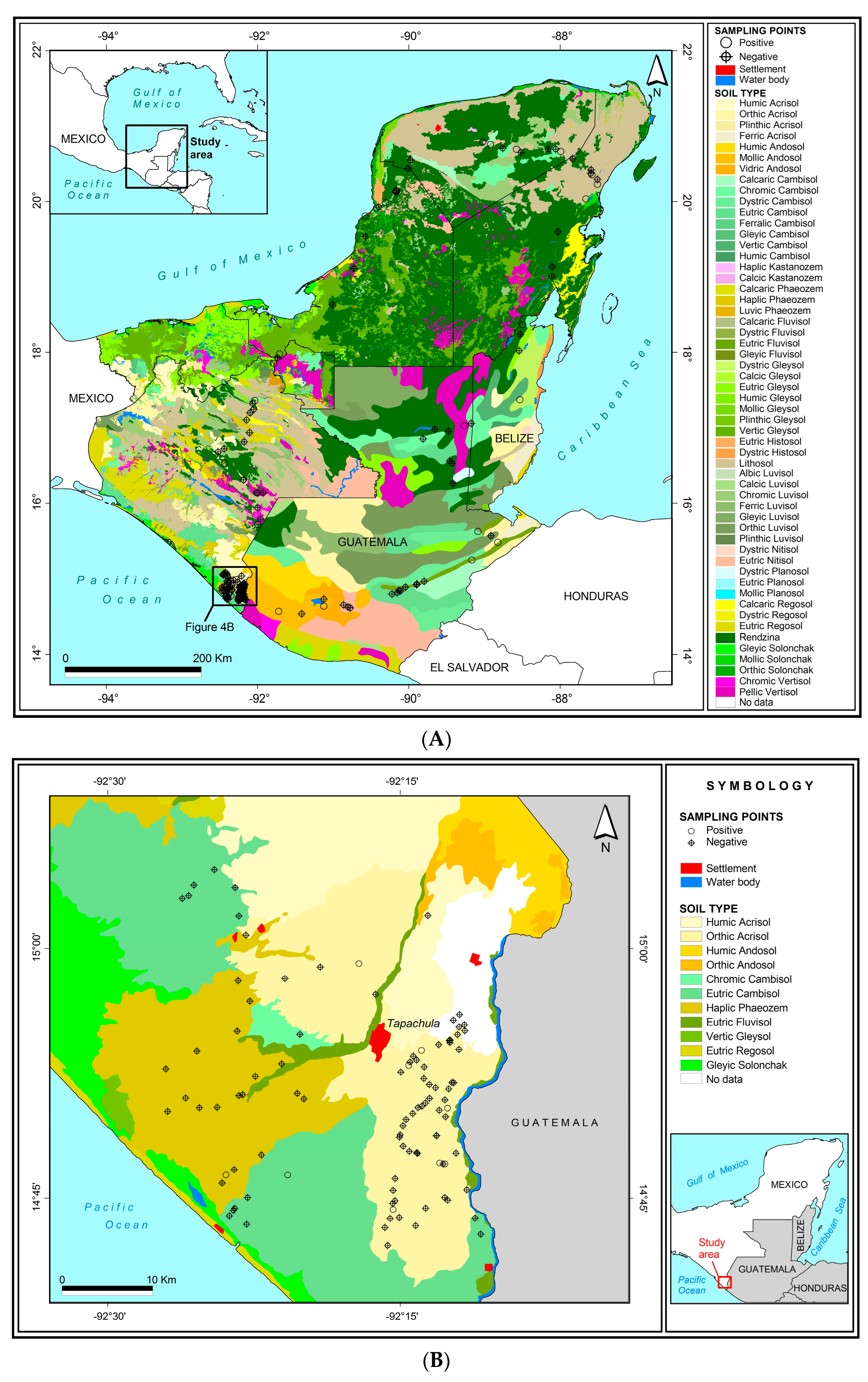
2.2. Soil Sampling
2.3. Bioassay of Soil Samples
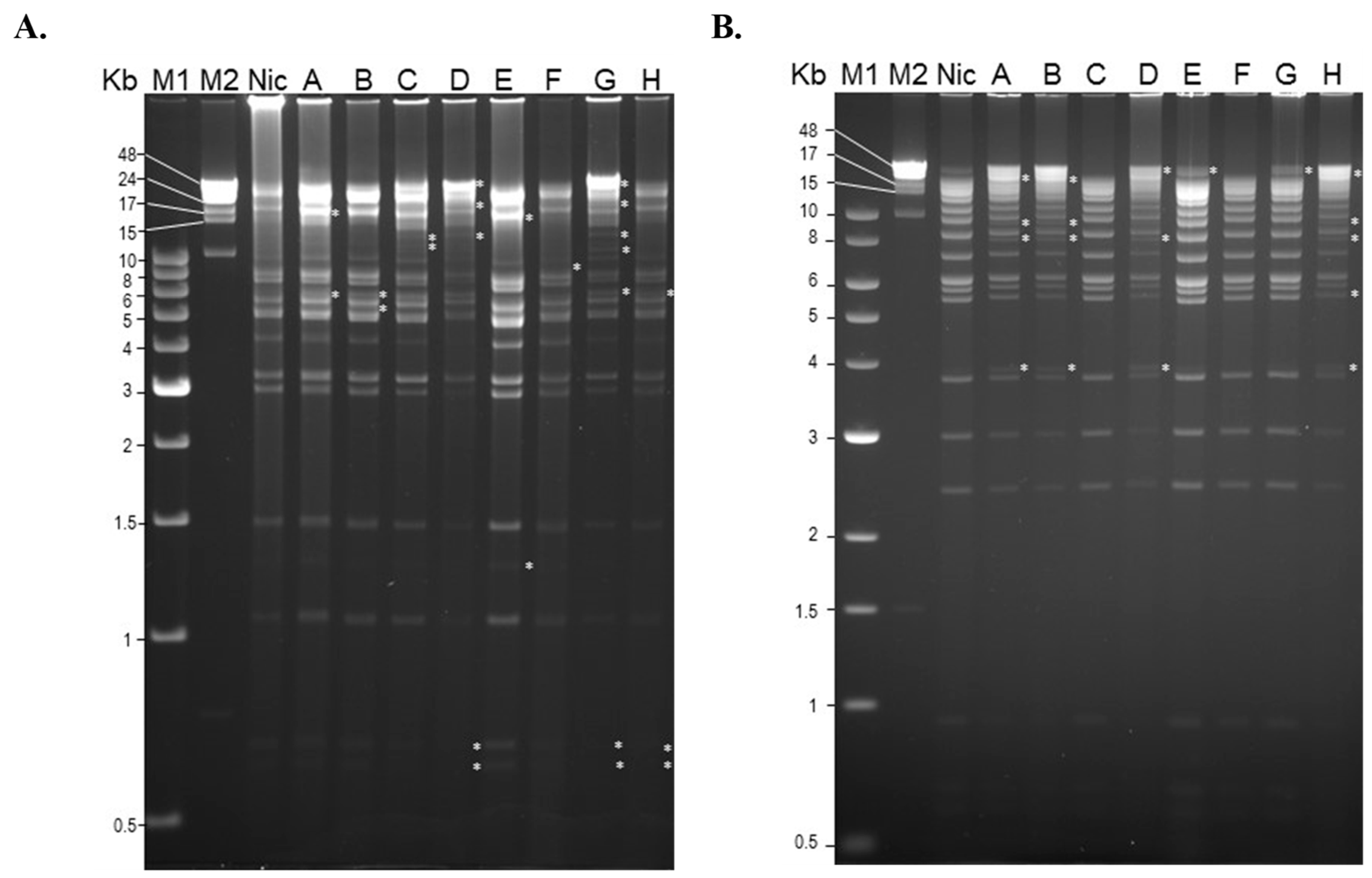
2.4. Restriction Endonuclease Analysis of Soil Isolates
2.5. Relationship between Soil OBs and Environmental Factors
3. Results
3.1. Calibration of the Soil Bioassay
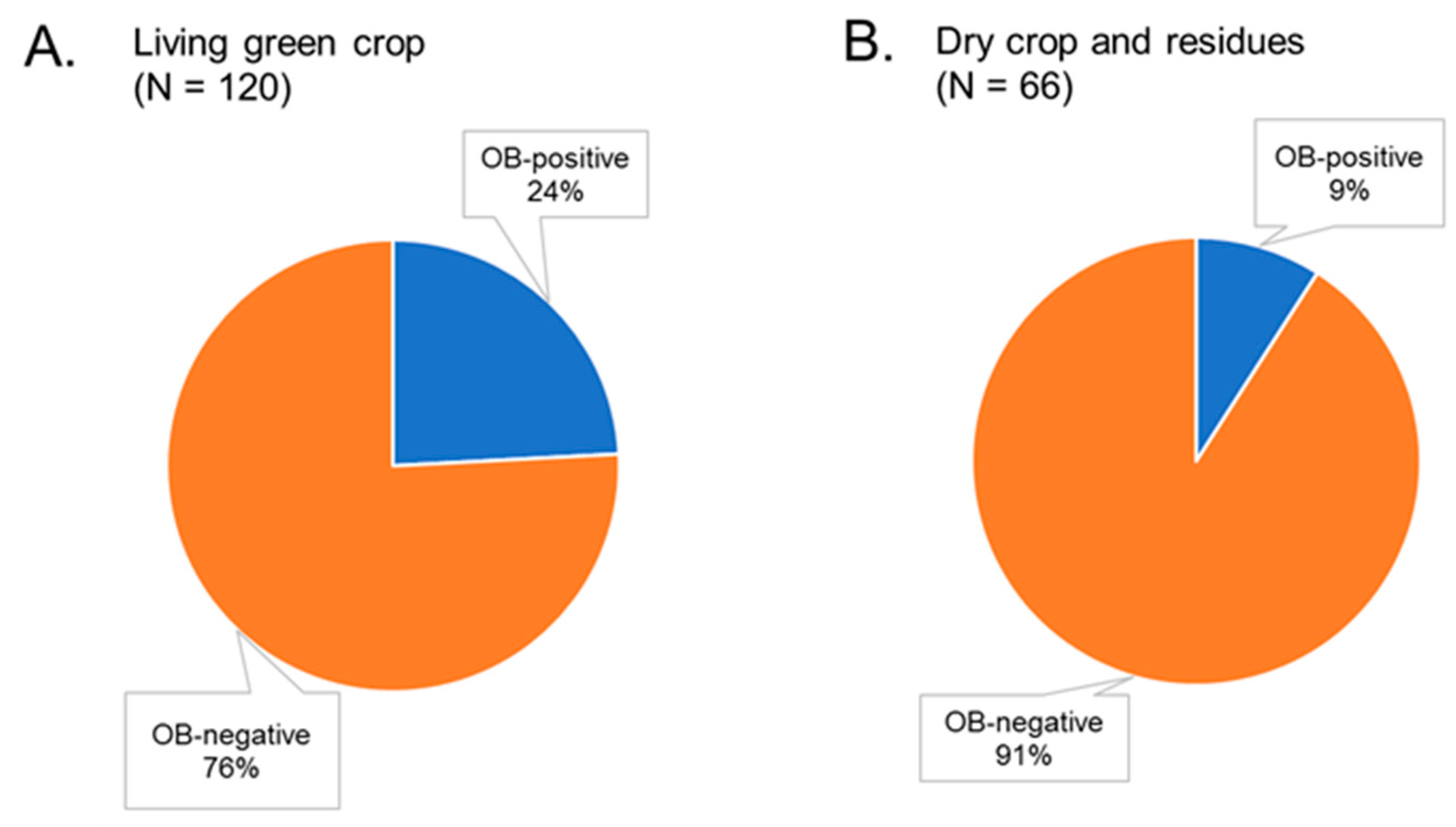
3.2. Bioassay of Soil Samples from Maize Fields
3.3. Restriction Endonuclease Analysis of Soil Isolates
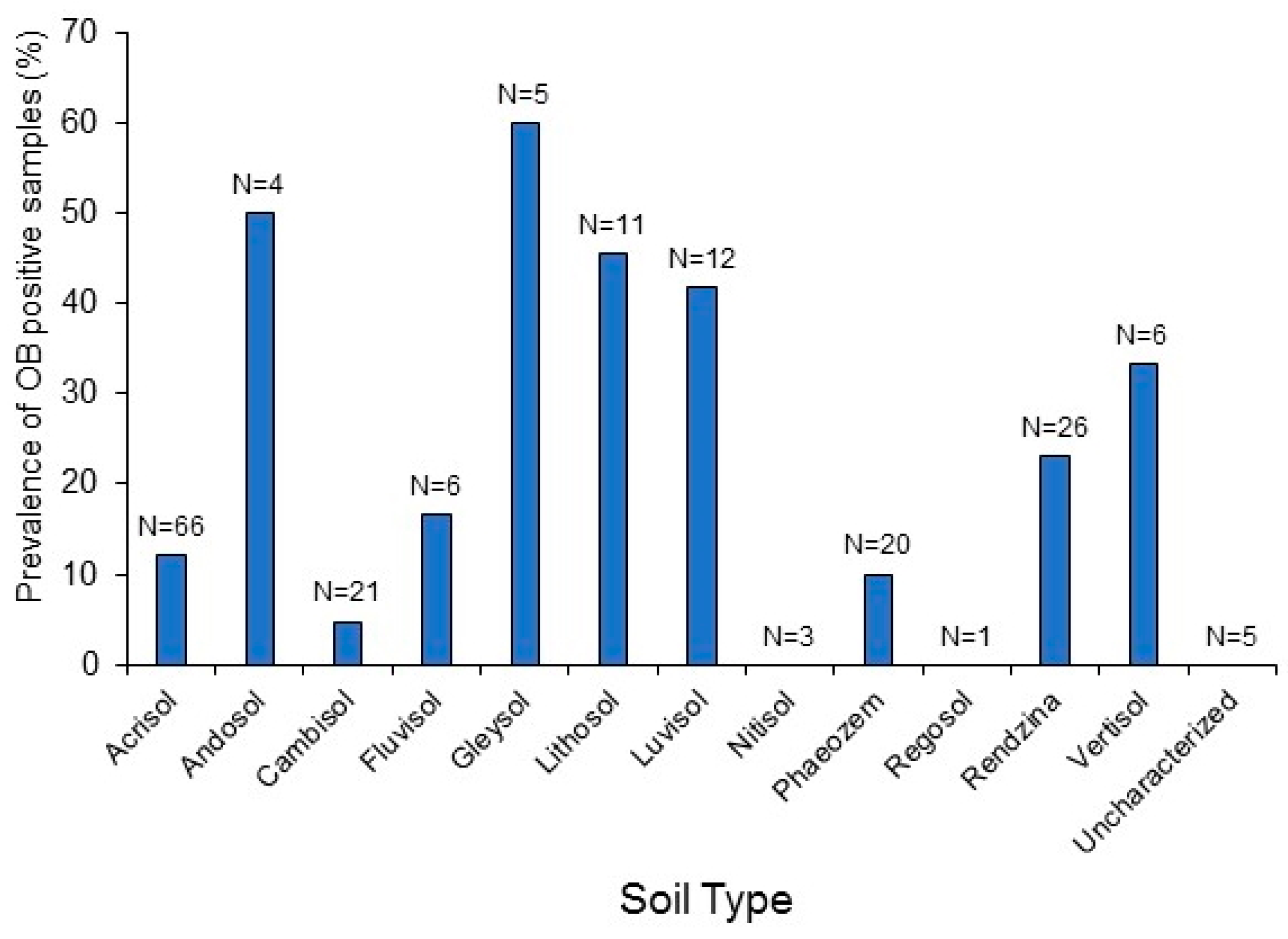
3.4. Relationship between Soil OBs and Environmental Factors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- D’Amico, V.; Elkinton, J.S. Rainfall effects on transmission of gypsy moth (Lepidoptera: Lymantriidae) nuclear polyhedrosis virus. Environ. Entomol. 1995, 24, 1144–1149. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Richter, A.R. Effect of agricultural operations and precipitation on vertical distribution of a nuclear polyhedrosis virus in soil. Biol. Control 1996, 6, 324–329. [Google Scholar] [CrossRef]
- Hochberg, M. The potential role of pathogens in biological control. Nature 1989, 337, 262–265. [Google Scholar] [CrossRef]
- Williams, T. Viruses. In Ecology of Invertebrate Diseases; Hajek, A.E., Shapiro-Ilan, D.I., Eds.; Wiley: Chichester, UK, 2018; pp. 215–285. [Google Scholar]
- Fuxa, J.R. Ecology of insect nucleopolyhedroviruses. Agric. Ecosyst. Environ. 2004, 103, 27–43. [Google Scholar] [CrossRef]
- Ignoffo, C.M.; Hostetter, D.L.; Sikorowski, P.P.; Sutter, G.; Brooks, W.M. Inactivation of representative species of entomopathogenic viruses, a bacterium, fungus, and protozoan by an ultraviolet light source. Environ. Entomol. 1977, 6, 411–415. [Google Scholar] [CrossRef]
- Olofsson, E. Dispersal of the nuclear polyhedrosis virus of Neodiprion sertifer from soil to pine foliage with dust. Entomol. Exp. Appl. 1988, 46, 181–186. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Matter, M.M.; Abdel-Rahman, A.; Micinski, S.; Richter, A.R.; Flexner, J.L. Persistence and distribution of wild-type and recombinant nucleopolyhedroviruses in soil. Microb. Ecol. 2001, 41, 222–231. [Google Scholar] [CrossRef]
- Fuxa, J.R. Threshold concentrations of nucleopolyhedrovirus in soil to initiate infections in Heliothis virescens on cotton plants. Microb. Ecol. 2008, 55, 530–539. [Google Scholar] [CrossRef]
- Lee, Y.; Fuxa, J.R. Transport of wild-type and recombinant nucleopolyhedroviruses by scavenging and predatory arthropods. Microb. Ecol. 2000, 39, 301–313. [Google Scholar]
- Evans, H.F.; Bishop, J.M.; Page, E.A. Methods for the quantitative assessment of nuclear-polyhedrosis virus in soil. J. Invertebr. Pathol. 1980, 35, 1–8. [Google Scholar] [CrossRef]
- de Moraes, R.R.; Maruniak, J.E.; Funderburk, J.E. Methods for detection of Anticarsia gemmatalis nucleopolyhedrovirus DNA in soil. Appl. Environ. Microbiol. 1999, 65, 2307–2311. [Google Scholar] [CrossRef]
- England, L.S.; Trevors, J.T.; Holmes, S.B. Extraction and detection of baculoviral DNA from lake water, detritus and forest litter. J. Appl. Microbiol. 2001, 90, 630–636. [Google Scholar] [CrossRef]
- Ebling, P.M.; Holmes, S.B. A refined method for the detection of baculovirus occlusion bodies in forest terrestrial and aquatic habitats. Pest Manag. Sci. 2002, 58, 1216–1222. [Google Scholar] [CrossRef]
- Richards, A.R.; Christian, P.D. A rapid bioassay screen for quantifying nucleopolyhedroviruses (Baculoviridae) in the environment. J. Virol. Meth. 1999, 82, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Rios-Velasco, C.; Gallegos-Morales, G.; Del Rincón-Castro, M.C.; Cerna-Chávez, E.; Sánchez-Peña, S.R.; Siller, M.C. Insecticidal activity of native isolates of Spodoptera frugiperda multiple nucleopolyhedrovirus from soil samples in Mexico. Fl. Entomol. 2011, 94, 716–718. [Google Scholar] [CrossRef]
- Murillo, R.; Elvira, S.; Munoz, D.; Williams, T.; Caballero, P. Genetic and phenotypic variability in Spodoptera exigua nucleopolyhedrovirus isolates from greenhouse soils in southern Spain. Biol. Control 2006, 38, 157–165. [Google Scholar] [CrossRef]
- Murillo, R.; Munoz, D.; Ruíz-Portero, M.C.; Alcázar, M.D.; Belda, J.E.; Williams, T.; Caballero, P. Abundance and genetic structure of nucleopolyhedrovirus populations in greenhouse substrate reservoirs. Biol. Control 2007, 42, 216–225. [Google Scholar] [CrossRef]
- Infante-Rodríguez, D.A.; Berber, J.J.; Mercado, G.; Valenzuela-González, J.; Muñoz, D.; Williams, T. Earthworm mediated dispersal of baculovirus occlusion bodies: Experimental evidence from a model system. Biol. Control 2016, 100, 18–24. [Google Scholar] [CrossRef]
- Hernández-Melchor, L.; Ramírez-Santiago, J.J.; Mercado, G.; Williams, T. Vertical dispersal of nucleopolyhedrovirus occlusion bodies in soil by the earthworm Amynthas gracilus: A field-based estimation. Biocontr. Sci. Technol. 2020, 30, 602–608. [Google Scholar] [CrossRef]
- Lei, C.; Yang, J.; Wang, J.; Hu, J.; Sun, X. Molecular and biological characterization of Spodoptera frugiperda multiple nucleopolyhedrovirus field isolate and genotypes from China. Insects 2020, 11, 777. [Google Scholar] [CrossRef] [PubMed]
- Firake, D.M.; Behere, G.T. Natural mortality of invasive fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) in maize agroecosystems of northeast India. Biol. Control 2020, 148, 104303. [Google Scholar] [CrossRef]
- Ginting, S.; Zarkani, A.; Wibowo, R.H.; Sipriyadi. New invasive pest, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) attacking corns in Bengkulu, Indonesia. Serangga 2020, 25, 105–117. [Google Scholar]
- Valicente, F.H.; Barreto, M.R. Levantamento dos inimigos naturais da lagarta do cartucho do milho, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae), na região de Cascavel, PR. An. Soc. Entomol. Brasil 1999, 28, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Molina-Ochoa, J.; Lezama-Gutierrez, R.; Gonzalez-Ramirez, M.; Lopez-Edwards, M.; Rodriguez-Vega, M.A.; Arceo-Palacios, F. Pathogens and parasitic nematodes associated with populations of fall armyworm (Lepidoptera: Noctuidae) larvae in Mexico. Fl. Entomol. 2003, 86, 244–253. [Google Scholar] [CrossRef]
- García-Gutiérrez, C.; Escobedo-Bonilla, C.M.; López, M.A. Infectivity of a Sinaloa native isolate of multicapsid nuclear polyhedrosis virus (SfMNPV) against fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Southwest. Entomol. 2013, 38, 597–604. [Google Scholar] [CrossRef]
- Fuxa, J.R. Prevalence of viral infections in populations of fall armyworm, Spodoptera frugiperda, in southeastern Louisiana. Environ. Entomol. 1982, 11, 239–242. [Google Scholar] [CrossRef]
- Estrada-Vírgen, O.; Campos, J.C.; Bermudez, A.R.; Velasco, C.R.; Cazola, C.C.; Aquino, N.I.; Cancino, E.R. Parasitoides y entomopatógenos nativos asociados al gusano cogollero Spodoptera frugiperda (Lepidoptera: Noctuidae) en Nayarit, México. Southwest. Entomol. 2013, 38, 339–344. [Google Scholar] [CrossRef]
- Kenis, M.; Benelli, G.; Biondi, A.; Calatayud, P.A.; Day, R.; Desneux, N.; Harrison, R.D.; Kriticos, D.; Rwomushana, I.; van den Berg, J.; et al. Invasiveness, biology, ecology, and management of the fall armyworm, Spodoptera frugiperda. Entomol. Gen. 2023. [Google Scholar] [CrossRef]
- García-Banderas, D.; Tamayo-Mejía, F.; Pineda, S.; de la Rosa, J.I.F.; Lasa, R.; Chavarrieta-Yáñez, J.M.; Gervasio-Rosas, E.; Zamora-Avilés, N.; Morales, S.I.; Ramos-Ortiz, S.; et al. Biological characterization of two Spodoptera frugiperda nucleopolyhedrovirus isolates from Mexico and evaluation of one isolate in a small-scale field trial. Biol. Control 2020, 149, 104316. [Google Scholar] [CrossRef]
- Mihm, J.A. Técnicas Eficientes para la Crianza Masiva e Infestacion de Insectos, en la Selection de las Plantas Hospedantes para Resistencia al Gusano Cogollero, Spodoptera frugiperda; Centro Internacional de Mejoramiento de Maiz y Trigo (CIMMYT): El Batán, Mexico, 1984; p. 16. [Google Scholar]
- Martínez, A.M.; Williams, T.; López-Ferber, M.; Caballero, P. Optical brighteners do not influence covert baculovirus infection of Spodoptera frugiperda. Appl. Environ. Microbiol. 2005, 71, 1668–1670. [Google Scholar] [CrossRef] [Green Version]
- Simón, O.; Palma, L.; Williams, T.; López-Ferber, M.; Caballero, P. Analysis of a naturally-occurring deletion mutant of Spodoptera frugiperda multiple nucleopolyhedrovirus reveals sf58 as a new per os infectivity factor of lepidopteran-infecting baculoviruses. J. Invertebr. Pathol. 2012, 109, 117–126. [Google Scholar] [CrossRef] [Green Version]
- Velasco, E.A.; Molina-Ruíz, C.S.; Gómez-Díaz, J.S.; Williams, T. Properties of nucleopolyhedrovirus occlusion bodies from living and virus-killed larvae of Spodoptera frugiperda (Lepidoptera: Noctuidae). Biol. Control 2022, 174, 105008. [Google Scholar] [CrossRef]
- Escribano, A.; Williams, T.; Goulson, D.; Cave, R.D.; Chapman, J.W.; Caballero, P. Selection of a nucleopolyhedrovirus for control of Spodoptera frugiperda (Lepidoptera: Noctuidae): Structural, genetic and biological comparison of four isolates from the Americas. J. Econ. Entomol. 1999, 92, 1079–1085. [Google Scholar] [CrossRef] [PubMed]
- Simón, O.; Williams, T.; López-Ferber, M.; Taulemesse, J.M.; Caballero, P. Population genetic structure determines the speed of kill and occlusion body production in Spodoptera frugiperda multiple nucleopolyhedrovirus. Biol. Control 2008, 44, 321–330. [Google Scholar] [CrossRef]
- Simón, O.; Palma, L.; Beperet, I.; Muñoz, D.; López-Ferber, M.; Caballero, P.; Williams, T. Sequence comparison between three geographically distinct Spodoptera frugiperda multiple nucleopolyhedrovirus isolates: Detecting positively selected genes. J. Invertebr. Pathol. 2011, 107, 33–42. [Google Scholar] [CrossRef]
- Hunter-Fujita, F.R.; Entwistle, P.F.; Evans, H.F.; Crook, N.E. Insect Viruses and Pest Management; Wiley: Chichester, UK, 1998; p. 620. [Google Scholar]
- Crawley, M.J. GLIM for Ecologists; Blackwell Science: Oxford, UK, 1993; p. 392. [Google Scholar]
- Instituto Nacional de Investigaciones Forestales y Agropecuarias (INIFAP). Comisión Nacional para el Conocimiento y Uso de la Biodiversidad (CONABIO), (1995). Edafología. Scale 1:250000 and 1:1000000. Mexico. Available online: https://idegeo.geoint.mx/layers/geonode%3Aedafologia/layer_info_metadata (accessed on 22 October 2022).
- Food and Agriculture Organization. FAO/UNESCO Digital Soil Map of the World. Available online: https://data.apps.fao.org/map/catalog/static/api/records/446ed430-8383-11db-b9b2-000d939bc5d8 (accessed on 21 October 2022).
- ArcGis for Windows, ver. 10.7. Available online: https://enterprise.arcgis.com/en/get-started/10.7/windows/what-is-arcgis-enterprise-.htm (accessed on 21 October 2022).
- Boogaard, B.; Evers, F.; van Lent, J.W.; van Oers, M.M. The baculovirus Ac108 protein is a per os infectivity factor and a component of the ODV entry complex. J. Gen. Virol. 2019, 100, 669–678. [Google Scholar] [CrossRef]
- Ramírez-Arias, F.G.; Lasa, R.; Murillo, R.; Navarro-de-la-Fuente, L.; Mercado, G.; Williams, T. Post-mortem incubation influences occlusion body production in nucleopolyhedrovirus-infected larvae of Spodoptera frugiperda. Biol. Control 2019, 135, 33–40. [Google Scholar] [CrossRef]
- Roome, R.E.; Daoust, R.A. Survival of the nuclear polyhedrosis virus of Heliothis armigera on crops and in soil in Botswana. J. Invertebr. Pathol. 1976, 27, 7–12. [Google Scholar] [CrossRef]
- Jaques, R.P. Occurrence and accumulation of viruses of Trichoplusia ni in treated field plots. J. Invertebr. Pathol. 1974, 23, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Podgwaite, J.D.; Shields, K.S.; Zerillo, R.T.; Bruen, R.B. Environmental persistence of the nucleopolyhedrosis virus of the gypsy moth, Lymantria dispar. Environ. Entomol. 1979, 8, 528–536. [Google Scholar] [CrossRef] [Green Version]
- Hukuhara, T. Further studies on the distribution of a nuclear-polyhedrosis virus of the fall webworm, Hyphantria cunea, in soil. J. Invertebr. Pathol. 1973, 22, 345–350. [Google Scholar] [CrossRef]
- Christian, P.D.; Richards, A.R.; Williams, T. Differential adsorption of occluded and nonoccluded insect-pathogenic viruses to soil-forming minerals. Appl. Environ. Microbiol. 2006, 72, 4648–4652. [Google Scholar] [CrossRef] [PubMed]
- Rios-Velasco, C.; Gallegos-Morales, G.; Berlanga-Reyes, D.; Cambero-Campos, J.; Romo-Chacón, A. Mortality and production of occlusion bodies in Spodoptera frugiperda larvae (Lepidoptera: Noctuidae) treated with nucleopolyhedrovirus. Fl. Entomol. 2012, 95, 752–757. [Google Scholar] [CrossRef]
- Jaques, R. Stability of viruses in the environment. In Viral Insecticides for Biological Control; Maramorosch, K., Sherman, K.E., Eds.; Academic Press: Orlando, FL, USA, 1985; pp. 285–360. [Google Scholar]
- Gómez-Bonilla, Y.; López-Ferber, M.; Caballero, P.; Léry, X.; Muñoz, D. Costa Rican soils contain highly insecticidal granulovirus strains against Phthorimaea operculella and Tecia solanivora. J. Appl. Entomol. 2012, 136, 530–538. [Google Scholar] [CrossRef]
- Clavijo, G.; Williams, T.; Muñoz, D.; Caballero, P.; López-Ferber, M. Mixed genotype transmission bodies and virions contribute to the maintenance of diversity in an insect virus. Proc. R. Soc. B Biol. Sci. 2010, 277, 943–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simón, O.; Williams, T.; López-Ferber, M.; Caballero, P. Genetic structure of a Spodoptera frugiperda nucleopolyhedrovirus population: High prevalence of deletion genotypes. Appl. Environ. Microbiol. 2004, 70, 5579–5588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baillie, V.L.; Bouwer, G. High levels of genetic variation within Helicoverpa armigera nucleopolyhedrovirus populations in individual host insects. Arch. Virol. 2012, 157, 2281–2289. [Google Scholar] [CrossRef] [PubMed]
- Cory, J.S.; Green, B.M.; Paul, R.K.; Hunter-Fujita, F. Genotypic and phenotypic diversity of a baculovirus population within an individual insect host. J. Invertebr. Pathol. 2005, 89, 101–111. [Google Scholar] [CrossRef]
- Aguirre, E.; Beperet, I.; Williams, T.; Caballero, P. Genetic variability of Chrysodeixis includens nucleopolyhedrovirus (ChinNPV) and the insecticidal characteristics of selected genotypic variants. Viruses 2019, 11, 581. [Google Scholar] [CrossRef] [Green Version]
- Fuxa, J.R.; Geaghan, J.P. Multiple-regression analysis of factors affecting prevalence of nuclear polyhedrosis virus in Spodoptera frugiperda (Lepidoptera: Noctuidae) populations. Environ. Entomol. 1983, 12, 311–316. [Google Scholar] [CrossRef]
- Peng, F.; Fuxa, J.R.; Richter, A.R.; Johnson, S.J. Effect of heat-sensitive agents, soil type, moisture, and leaf surface on persistence of Anticarsia gemmatalis (Lepidoptera: Noctuidae) nucleopolyhedrovirus. Environ. Entomol. 1999, 28, 330–338. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Richter, A.R. Quantification of soil-to-plant transport of recombinant nucleopolyhedrovirus: Effects of soil type and moisture, air currents, and precipitation. Appl. Environ. Microbiol. 2001, 67, 5166–5170. [Google Scholar] [CrossRef] [PubMed]
- Fuxa, J.R.; Richter, A.R.; Milks, M.L. Threshold distances and depths of nucleopolyhedrovirus in soil for transport to cotton plants by wind and rain. J. Invertebr. Pathol. 2007, 95, 60–70. [Google Scholar] [CrossRef]
- Small, D.A.; Moore, N.F. Measurement of surface charge of baculovirus polyhedra. Appl. Environ. Microbiol. 1987, 53, 598–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Small, D.A.; Moore, N.F.; Entwistle, P.F. Hydrophobic interactions involved in attachment of a baculovirus to hydrophobic surfaces. Appl. Environ. Microbiol. 1986, 52, 220–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chesworth, W. Encyclopedia of Soil Science—Encyclopedia of Earth Sciences Series; Springer: Dordrecht, The Netherlands, 2008; p. 902. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; International Union of Soil Sciences (IUSS): Vienna, Austria, 2022; Available online: https://www3.ls.tum.de/fileadmin/w00bds/boku/downloads/wrb/WRB_fourth_edition_2022-09-19.pdf (accessed on 20 October 2022).
- ISRIC World Soil Information Hub. International Soil Reference and Information Centre. Available online: https://www.isric.org (accessed on 20 October 2022).
- FAO Soils Portal—Data Hub. Available online: https://www.fao.org/soils-portal/data-hub/en/ (accessed on 24 October 2022).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State/Country 1 | No. of Samples Tested | No. of Positive Samples (%) | No. of Larvae Tested Against OB-Positive Samples | No. of Virus Killed Larvae (%) |
|---|---|---|---|---|
| Chiapas | 120 | 14 (11.7%) | 765 | 22 (2.9%) |
| Tabasco | 1 | 0 (0%) | - | - |
| Campeche | 8 | 4 (50.0%) | 240 | 8 (3.3%) |
| Yucatán | 16 | 6 (37.5%) | 360 | 15 (4.2%) |
| Quintana Roo | 10 | 3 (30.0%) | 180 | 8 (4.4%) |
| Belize | 4 | 2 (50.0%) | 120 | 2 (1.7%) |
| Guatemala | 27 | 6 (22.2%) | 360 | 6 (1.7%) |
| Totals: | 186 | 35 (18.8%) | 2025 | 61 (3.0%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, T.; Melo-Molina, G.d.C.; Jiménez-Fernández, J.A.; Weissenberger, H.; Gómez-Díaz, J.S.; Navarro-de-la-Fuente, L.; Richards, A.R. Presence of Spodoptera frugiperda Multiple Nucleopolyhedrovirus (SfMNPV) Occlusion Bodies in Maize Field Soils of Mesoamerica. Insects 2023, 14, 80. https://doi.org/10.3390/insects14010080
Williams T, Melo-Molina GdC, Jiménez-Fernández JA, Weissenberger H, Gómez-Díaz JS, Navarro-de-la-Fuente L, Richards AR. Presence of Spodoptera frugiperda Multiple Nucleopolyhedrovirus (SfMNPV) Occlusion Bodies in Maize Field Soils of Mesoamerica. Insects. 2023; 14(1):80. https://doi.org/10.3390/insects14010080
Chicago/Turabian StyleWilliams, Trevor, Guadalupe del Carmen Melo-Molina, Jaime A. Jiménez-Fernández, Holger Weissenberger, Juan S. Gómez-Díaz, Laura Navarro-de-la-Fuente, and Andrew R. Richards. 2023. "Presence of Spodoptera frugiperda Multiple Nucleopolyhedrovirus (SfMNPV) Occlusion Bodies in Maize Field Soils of Mesoamerica" Insects 14, no. 1: 80. https://doi.org/10.3390/insects14010080