
Environmental Factors Drive Chalcid Body Size Increases with Altitudinal Gradients for Two Hyper-Diverse Taxa

Abstract
:Simple Summary
Abstract
1. Introduction

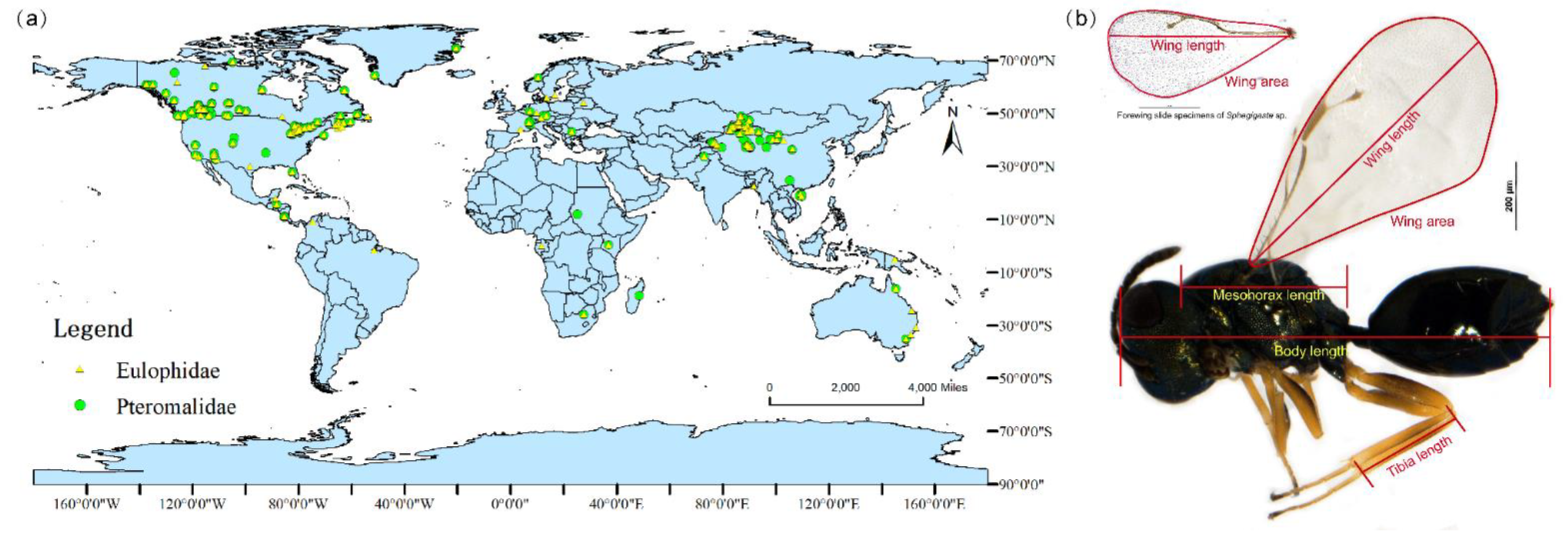{kind=link}
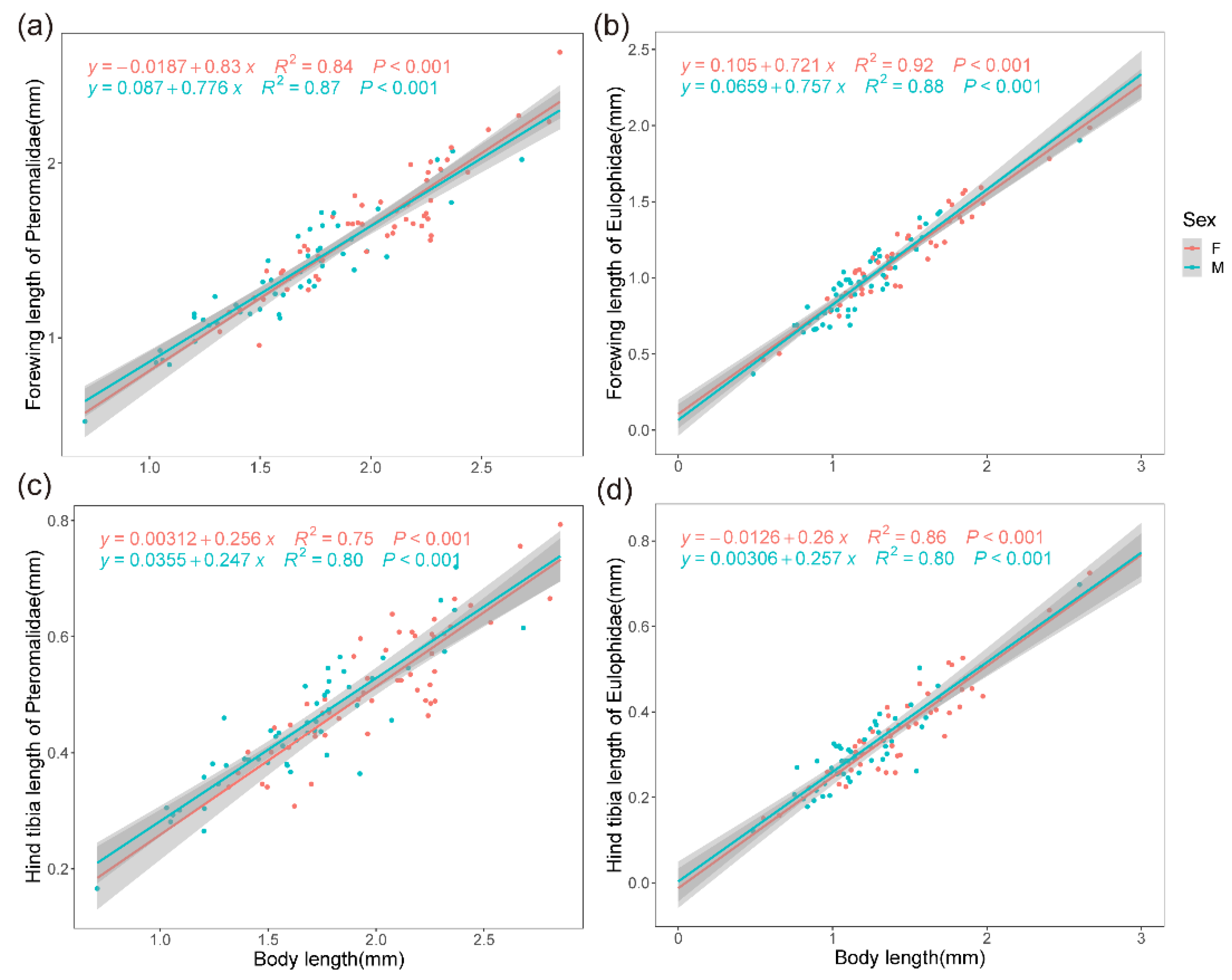{kind=link}
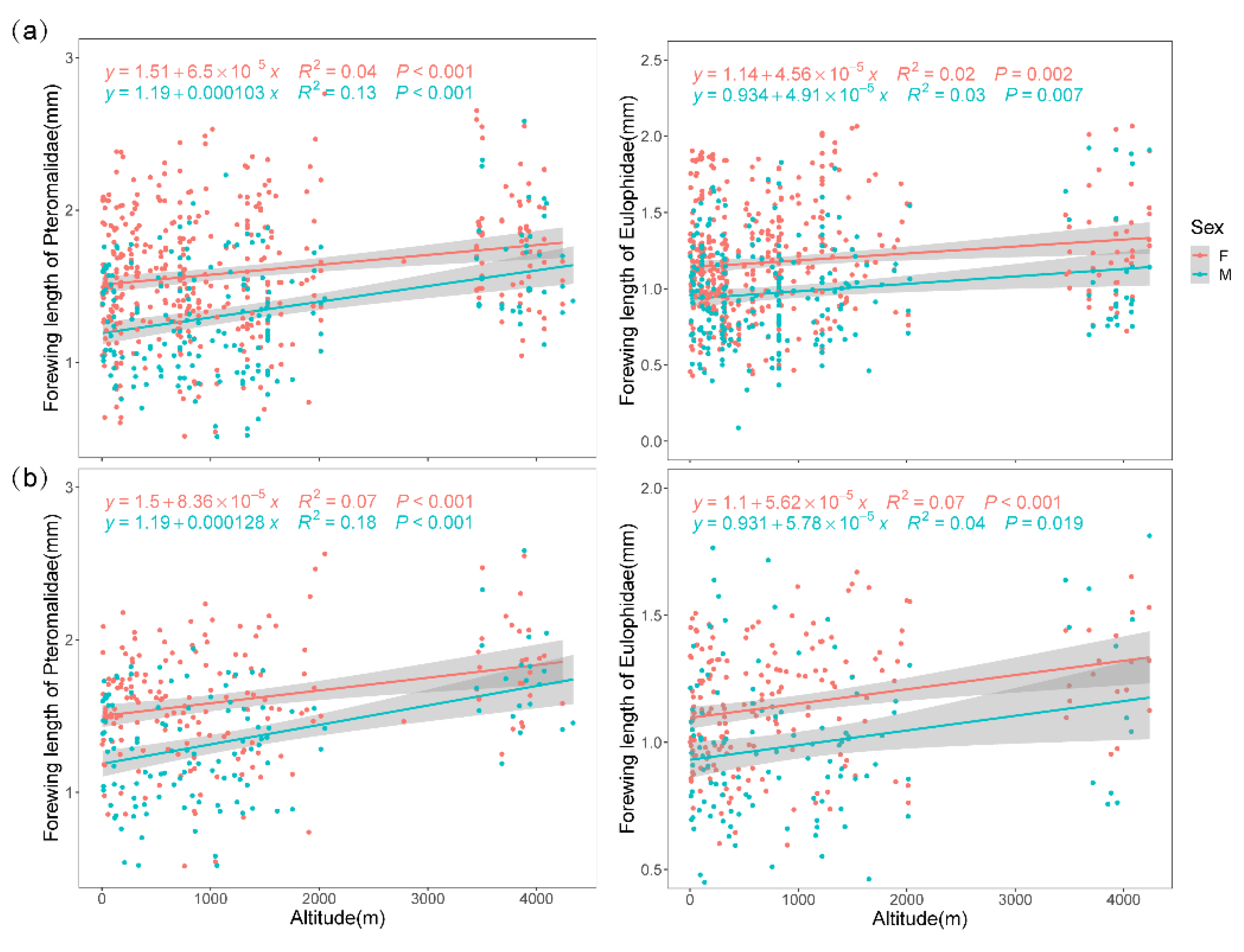{kind=link}
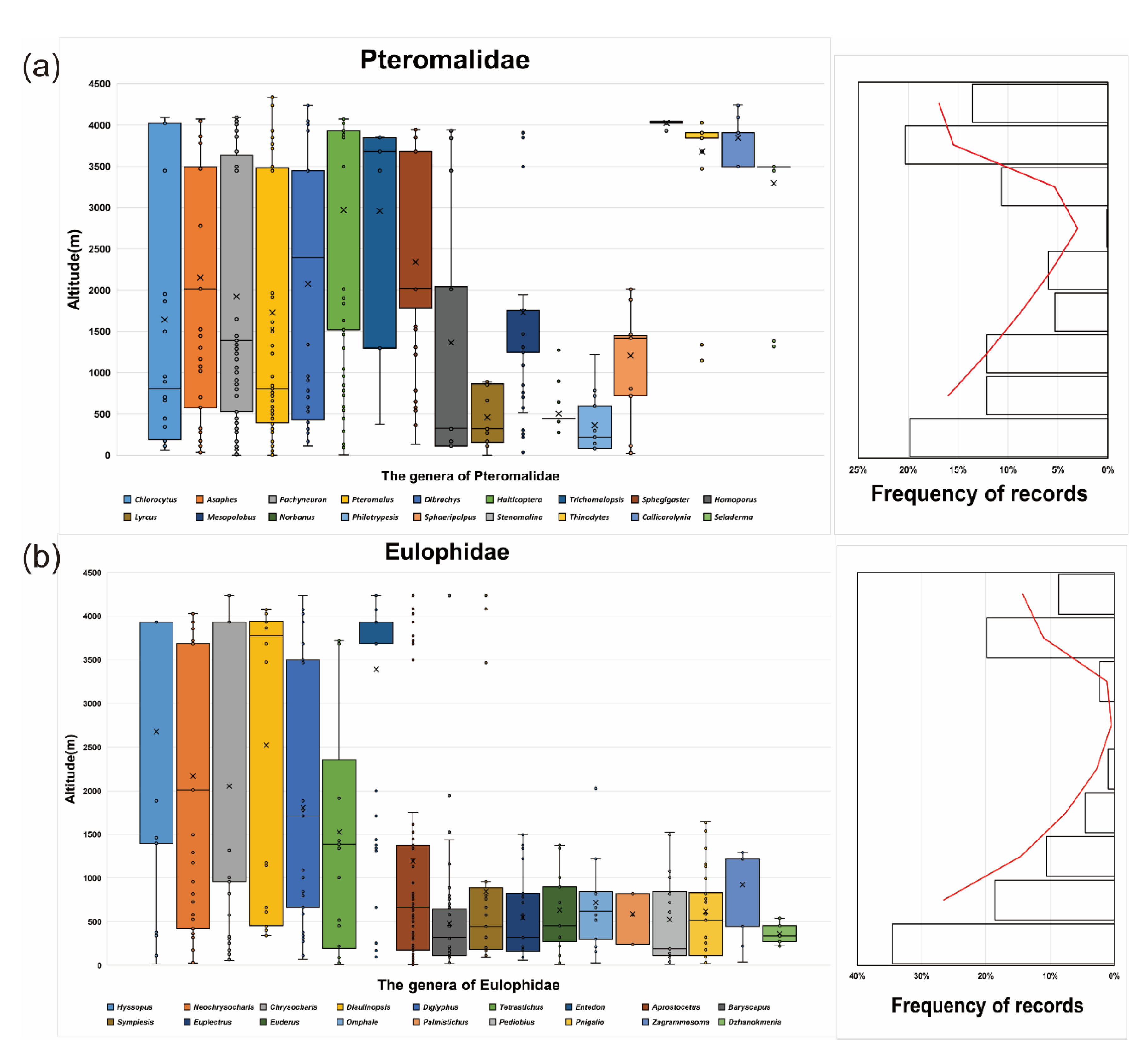{kind=link}
| Taxa | Genus or Species | Bergmann’s Rule | Data Sources | Factors | Measured Treat | Reference |
|---|---|---|---|---|---|---|
| Apidae | Bombus | CB | Specimens from museum | Latitude | Thorax width | [38] |
| Apidae | B. lucorum, B. magnus and B. cryptarum | B | Specimens collected from Great Britain | Latitude | Thorax width | [39] |
| Apidae | B. vancouverensis | B | Specimens collected from the United States | Latitude | Inter-tegular span (ITS), Forewing area Community-weighted mean of ITS, relative forewing length, relative hind tibia length, hair length at mesonotum, relative hair length at mesonotum | [40] |
| B. vosnesenskii | CB | Altitude | ||||
| Apoidea, Megachilidae, Andrenidae, Halictidae, Colletidae | Macrotera sp., Lasioglossum (Dialictus) sp., Lasioglossum (Lasioglossum) sp., Halictus sp. | B (only for Relative hair length) CB | Specimens collected from north Mexico | Altitude | [41] | |
| Apoidea | Andrena, Dasypoda, Halictus, Panurginus | B | Data from database | Latitude | ITS | [42] |
| Bombus, Colletes, Melitta | CB | |||||
| Apoidea | At family level | B, species larger than 27.81 mg (dry weight) | Specimens collected from Mediterranean; data from articles and museum collections | Temperature | ITS | [43] |
| CB, species less than 27.81 mg | ||||||
| Braconidae | At family level | CB | Literature recorded | Latitude | Body length | [44] |
| Ichneumonidae | ||||||
| Bethylidae | Pristocera rufa | CB | Literature recorded | Latitude | Body length | [20] |
| Braconinae | At family level | CB | ||||
| Encyrtidae | Comperiella bifasciata | CB | ||||
| Ichneumonidae | At family level | B | ||||
| Braconidae, Ichneumonidae | At family level | CB | Specimens collected from Canada and data from BOLD system | Latitude | Hind tibia lengths | [45] |
| Formicidae | Ant species from 427 genera | CB | Specimens collected from the United States | Altitude/latitude | Weber’s length | [46] |
| Formicidae | Leptothorax acervorum | B | Specimens collected from Alps | Altitude | Maximum cephalic width | [47] |
| Formicidae | At family level | CB | Specimens collected from Australia, South Africa | Altitude | Body mass, critical thermal maximum (CTmax) | [48] |
| Formicidae | Leptothorax acervorum | B | Specimens collected throughout Europe | Latitudes | Thorax length | [49] |
| Mymaridae | Anaphes cultripennis | B | Specimens collected from Norway | Altitude | Body length | [21] |
| Vespidae | Polistes bahamensis, P. bellicosus, P. comanchus, P. dorsalis, P. exclamans | B | Specimens from museum | Latitude | Body length | [50] |
| P. annularis, P. apachus, P. Carolina, P. flavus, P. fuscatus, P. metricus, P. dominula, P. aurifer, P. bahamensis | CB | |||||
| Vespidae | Agelaia pallipes | CB | Specimens collected from Santuario de Iguaque, Colombia | Altitude | Hind femur length, mesosoma height, head width/length | [51] |
| B | Forewing width/length |
2. Materials and Methods
2.1. Study Area and Specimen Measurement
2.2. Environmental Data
2.3. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ollason, J.G.R.H. Peters 1986. The Ecological Implications of Body Size. Cambridge University Press, Cambridge. 329 Pages. ISBN 0-521-2886-x. J. Trop. Ecol. 1987, 3, 286–287. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward A Metabolic Theory of Ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Chown, S.L.; Gaston, K.J. Body Size Variation in Insects: A Macroecological Perspective. Biol. Rev. 2010, 85, 139–169. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, C. Über die Verhältnisse der Wärmeökonomie der Thiere zu Ihrer Grösse; Vandenhoeck und Ruprecht: Göttingen, Germany, 1847; Volume 1. [Google Scholar]
- Ashton, K.G.; Tracy, M.C.; de Queiroz, A. Is Bergmann’s Rule Valid for Mammals? Am. Nat. 2000, 156, 390–415. [Google Scholar] [CrossRef]
- Nwaogu, C.J.; Tieleman, B.I.; Bitrus, K.; Cresswell, W. Temperature and Aridity Determine Body Size Conformity to Bergmann’s Rule Independent of Latitudinal Differences in a Tropical Environment. J. Ornithol. 2018, 159, 1053–1062. [Google Scholar] [CrossRef] [Green Version]
- Blanckenhorn, W.U. Bergmann and Converse Bergmann Latitudinal Clines in Arthropods: Two Ends of a Continuum? Integr. Comp. Biol. 2004, 44, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Salomão, R.P.; Arriaga-Jiménez, A.; Kohlmann, B. The Relationship between Altitudinal Gradients, Diversity, and Body Size in a Dung Beetle (Coleoptera: Scarabaeinae: Onthophagus) Model System. Can. J. Zool. 2021, 99, 33–43. [Google Scholar] [CrossRef]
- Bidau, C.J.; Martí, D.A. Clinal Variation of Body Size in Dichroplus Pratensis (Orthoptera: Acrididae): Inversion of Bergmann’s and Rensch’s Rules. Ann. Entomol. Soc. Am. 2007, 100, 850–860. [Google Scholar] [CrossRef]
- Schutze, M.K.; Clarke, A.R. Converse Bergmann Cline in a Eucalyptus Herbivore, Paropsis Atomaria Olivier (Coleoptera: Chrysomelidae): Phenotypic Plasticity or Local Adaptation? Glob. Ecol. Biogeogr. 2008, 17, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, D.R.; Patel, P.; Ghimire, K.C.; Bista, M.; Kumar, B. Size-Based Intraspecific Variations along Altitudinal Gradient and Follow of Bergmann’s Rule in Parthenium Beetle, Zygogramma Bicolorata Pallister. J. Asia-Pac. Entomol. 2019, 22, 1173–1179. [Google Scholar] [CrossRef]
- Chown, S.L.; Klok, C.J. Altitudinal Body Size Clines: Latitudinal Effects Associated with Changing Seasonality. Ecography 2003, 26, 445–455. [Google Scholar] [CrossRef]
- Brehm, G.; Zeuss, D.; Colwell, R.K. Moth Body Size Increases with Elevation along a Complete Tropical Elevational Gradient for Two Hyperdiverse Clades. Ecography 2019, 42, 632–642. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Cheng, W.; Nakamura, A.; Tang, C.C.; Huang, S.; Odell, E.; Goodale, E.; Goodale, U.M.; Bonebrake, T.C. Elevational Clines in Morphological Traits of Subtropical and Tropical Butterfly Assemblages. Biol. J. Linn. Soc. 2018, 123, 506–517. [Google Scholar] [CrossRef] [Green Version]
- McCulloch, G.A.; Waters, J.M. Does Wing Reduction Influence the Relationship between Altitude and Insect Body Size? A Case Study Using New Zealand’s Diverse Stonefly Fauna. Ecol. Evol. 2018, 8, 953–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Valencia, L.I.; Moya-Raygoza, G. Body Size Variation of Diaphorina citri (Hemiptera: Psyllidae) through an Elevation Gradient. Ann. Entomol. Soc. Am. 2015, 108, 800–806. [Google Scholar] [CrossRef]
- Beerli, N.; Bärtschi, F.; Ballesteros-Mejia, L.; Kitching, I.J.; Beck, J. How Has the Environment Shaped Geographical Patterns of Insect Body Sizes? A Test of Hypotheses Using Sphingid Moths. J. Biogeogr. 2019, 46, 1687–1698. [Google Scholar] [CrossRef]
- Horne, C.R.; Hirst, A.G.; Atkinson, D. Insect Temperature–Body Size Trends Common to Laboratory, Latitudinal and Seasonal Gradients Are Not Found across Altitudes. Funct. Ecol. 2018, 32, 948–957. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Arango, S.; Cárdenas Henao, H.; Montoya-Lerma, J. Divergence in Bergmann’s Clines: Elevational Variation and Heritability of Body Size in a Leaf-Cutting Ant. Insectes Soc. 2020, 67, 355–366. [Google Scholar] [CrossRef]
- Traynor, R.E.; Mayhew, P.J. A Comparative Study of Body Size and Clutch Size across the Parasitoid Hymenoptera. Oikos 2005, 109, 305–316. [Google Scholar] [CrossRef]
- Sveum, P.; Solem, J.O. Anaphes cultripennis Debauché, 1948 (Hymenoptera, Mymaridae) in Norway. Fauna Nor. 1980, 27, 17–18. [Google Scholar]
- Jervis, M.A.; Ferns, P.N.; Heimpel, G.E. Body Size and the Timing of Egg Production in Parasitoid Wasps: A Comparative Analysis. Funct. Ecol. 2003, 17, 375–383. [Google Scholar] [CrossRef]
- Sagarra, L.A.; Vincent, C.; Stewart, R.K. Body Size as an Indicator of Parasitoid Quality in Male and Female Anagyrus kamali (Hymenoptera: Encyrtidae). Bull. Entomol. Res. 2001, 91, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Hodkinson, I.D. Terrestrial Insects along Elevation Gradients: Species and Community Responses to Altitude. Biol. Rev. 2005, 80, 489. [Google Scholar] [CrossRef] [Green Version]
- García-Navas, V.; Noguerales, V.; Cordero, P.J.; Ortego, J. Ecological Drivers of Body Size Evolution and Sexual Size Dimorphism in Short-Horned Grasshoppers (Orthoptera: Acrididae). J. Evol. Biol. 2017, 30, 1592–1608. [Google Scholar] [CrossRef] [Green Version]
- Baranov, V.; Jourdan, J.; Hunter-Moffatt, B.; Noori, S.; Schölderle, S.; Haug, J.T. Global Size Pattern in a Group of Important Ecological Indicators (Diptera, Chironomidae) Is Driven by Latitudinal Temperature Gradients. Insects 2021, 13, 34. [Google Scholar] [CrossRef]
- Wonglersak, R.; Fenberg, P.B.; Langdon, P.G.; Brooks, S.J.; Price, B.W. Temperature-body Size Responses in Insects: A Case Study of British Odonata. Ecol. Entomol. 2020, 45, 795–805. [Google Scholar] [CrossRef]
- Acquah-Lamptey, D.; Brändle, M.; Brandl, R.; Pinkert, S. Temperature-driven Color Lightness and Body Size Variation Scale to Local Assemblages of European Odonata but Are Modified by Propensity for Dispersal. Ecol. Evol. 2020, 10, 8936–8948. [Google Scholar] [CrossRef]
- Baroni, D.; Masoero, G. Complex Influence of Climate on the Distribution and Body Size of an Alpine Species. Insect Conserv. Divers. 2018, 11, 435–448. [Google Scholar] [CrossRef]
- Colinet, H.; Boivin, G.; Hance, T. Manipulation of Parasitoid Size Using the Temperature-Size Rule: Fitness Consequences. Oecologia 2007, 152, 425–433. [Google Scholar] [CrossRef]
- Bernardo, U.; Pedata, P.A.; Viggiani, G. Phenotypic Plasticity of Pigmentation and Morphometric Traits in Pnigalio soemius (Hymenoptera: Eulophidae). Bull. Entomol. Res. 2007, 97, 101–109. [Google Scholar] [CrossRef]
- Wang, T.; Keller, M.A. Larger Is Better in the Parasitoid Eretmocerus Warrae (Hymenoptera: Aphelinidae). Insects 2020, 11, 39. [Google Scholar] [CrossRef]
- Montgomery, K. Variation in Temperature With Altitude and Latitude. J. Geogr. 2006, 105, 133–135. [Google Scholar] [CrossRef]
- Angilletta, M.J., Jr.; Dunham, A.E. The Temperature-Size Rule in Ectotherms: Simple Evolutionary Explanations May Not Be General. Am. Nat. 2003, 162, 332–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, D. Temperature and Organism Size—A Biological Law for Ectotherms? In Advances in Ecological Research; Elsevier: San Leandro, CA, USA, 1994; Volume 25, pp. 1–58. ISBN 978-0-12-013925-5. [Google Scholar]
- Zhang, D.-W.; Xiao, Z.-J.; Zeng, B.-P.; Li, K.; Tang, Y.-L. Insect Behavior and Physiological Adaptation Mechanisms Under Starvation Stress. Front. Physiol. 2019, 10, 163. [Google Scholar] [CrossRef]
- Gergs, A.; Jager, T. Body Size-Mediated Starvation Resistance in an Insect Predator. J. Anim. Ecol. 2014, 83, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Delgado, V.H.; Sanabria-Urbán, S.; Serrano-Meneses, M.A.; Cueva del Castillo, R. The Converse to Bergmann’s Rule in Bumblebees, a Phylogenetic Approach. Ecol. Evol. 2016, 6, 6160–6169. [Google Scholar] [CrossRef] [PubMed]
- Scriven, J.J.; Whitehorn, P.R.; Goulson, D.; Tinsley, M.C. Bergmann’s Body Size Rule Operates in Facultatively Endothermic Insects: Evidence from a Complex of Cryptic Bumblebee Species. PLoS ONE 2016, 11, e0163307. [Google Scholar] [CrossRef]
- Lozier, J.D.; Parsons, Z.M.; Rachoki, L.; Jackson, J.M.; Pimsler, M.L.; Oyen, K.J.; Strange, J.; Dillon, M.E. Divergence in Body Mass, Wing Loading, and Population Structure Reveals Species-Specific and Potentially Adaptive Trait Variation Across Elevations in Montane Bumble Bees. Insect Syst. Divers. 2021, 5, 3. [Google Scholar] [CrossRef]
- Osorio-Canadas, S.; Flores-Hernández, N.; Sánchez-Ortiz, T.; Valiente-Banuet, A. Changes in Bee Functional Traits at Community and Intraspecific Levels along an Elevational Gradient in a Mexical-Type Scrubland. Oecologia 2022, 200, 145–158. [Google Scholar] [CrossRef]
- Gérard, M.; Vanderplanck, M.; Franzen, M.; Kuhlmann, M.; Potts, S.G.; Rasmont, P.; Schweiger, O.; Michez, D. Patterns of Size Variation in Bees at a Continental Scale: Does Bergmann’s Rule Apply? Oikos 2018, 127, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Osorio-Canadas, S.; Arnan, X.; Rodrigo, A.; Torné-Noguera, A.; Molowny, R.; Bosch, J. Body Size Phenology in a Regional Bee Fauna: A Temporal Extension of Bergmann’s Rule. Ecol. Lett. 2016, 19, 1395–1402. [Google Scholar] [CrossRef]
- Santos, A.M.C.; Quicke, D.L.J. Large-Scale Diversity Patterns of Parasitoid Insects: Parasitoid Diversity Patterns. Entomol. Sci. 2011, 14, 371–382. [Google Scholar] [CrossRef]
- Eagalle, T.I. Latitudinal Effects on Diversity and Body Size: A Case Study with Parasitoid and Parasitic Wasps. Master’s Thesis, The University of Guelph, Guelph, ON, Canada, 2014. [Google Scholar]
- Geraghty, M.J.; Dunn, R.R.; Sanders, N.J. Body Size, Colony Size, and Range Size in Ants (Hymenoptera: Formicidae): Are Patterns along Elevational and Latitudinal Gradients Consistent with Bergmann’s Rule? Myrmecol. News 2007, 10, 51–58. [Google Scholar]
- Bernadou, A.; Römermann, C.; Gratiashvili, N.; Heinze, J. Body Size but Not Colony Size Increases with Altitude in the Holarctic Ant, Leptothorax acervorum: Bergmann’s Rule in a Holarctic Ant. Ecol. Entomol. 2016, 41, 733–736. [Google Scholar] [CrossRef]
- Nowrouzi, S.; Andersen, A.N.; Bishop, T.R.; Robson, S.K.A. Is Thermal Limitation the Primary Driver of Elevational Distributions? Not for Montane Rainforest Ants in the Australian Wet Tropics. Oecologia 2018, 188, 333–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heinze, J.; Foitzik, S.; Fischer, B.; Wanke, T.; Kipyatkov, V.E. The Significance of Latitudinal Variation in Body Size in a Holarctic Ant, Leptothorax acervorum. Ecography 2003, 26, 349–355. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.E.; Sheehan, M.J. Ecogeographical Patterns of Body Size Differ among North American Paper Wasp Species. Insectes Soc. 2021, 68, 109–122. [Google Scholar] [CrossRef]
- Rodríguez-Jimenez, A.; Sarmiento, C.E. Altitudinal Distribution and Body Resource Allocation in a High Mountain Social Wasp (Hymenoptera: Vespidae). Neotrop. Entomol. 2008, 37, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G. Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); NRC Research Press: Ottawa, ON, Canada, 1997; ISBN 0-660-16669-0. [Google Scholar]
- Bouček, Z. Australasian Chalcidoidea (Hymenoptera). A Biosystematic Revision of Genera of Fourteen Families, with a Reclassification of Species; The Cambrian News Ltd.: Aberystwyth, Wales; CAB International: Wallingford, UK, 1988. [Google Scholar]
- Huang, D.-W.; Xiao, H. Fauna Sinica, Insecta, Volume 42, Hymenoptera: Pteromalidae; Science Press: Beijing, China, 2005. [Google Scholar]
- Bouček, Z.; Rasplus, J.Y. Ilustrated Key to West-Palearctic Genera of Pteromalidae; Institut National de la Recherche Agronomique: Paris, France, 1991; ISBN 2-7380-0343-5. [Google Scholar]
- De Vere Graham, M.W.R. The Pteromalidae of North-Western Europe (Hymenoptera: Chalcidoidea). Bull. Br. Mus. (Nat. Hist.) Entomol. Suppl. 1969, 16, 1–908. [Google Scholar] [CrossRef]
- Montejo-Kovacevich, G.; Salazar, P.A.; Smith, S.H.; Gavilanes, K.; Bacquet, C.N.; Chan, Y.F.; Jiggins, C.D.; Meier, J.I.; Nadeau, N.J. Genomics of Altitude-associated Wing Shape in Two Tropical Butterflies. Mol. Ecol. 2021, 30, 6387–6402. [Google Scholar] [CrossRef]
- Starmer, W.T.; Wolf, L.L. Causes of Variation in Wing Loading among Drosophila Species. Biol. J. Linn. Soc. 1989, 37, 247–261. [Google Scholar] [CrossRef]
- Bhat, S.S.; Sheridan, J.; Hourigan, K.; Thompson, M.C. Aspect Ratio Studies on Insect Wings. Phys. Fluids 2019, 31, 121301. [Google Scholar] [CrossRef]
- Sarig, A.; Ribak, G. To What Extent Can the Tiny Parasitoid Wasps, Eretmocerus Mundus, Fly Upwind? J. Appl. Entomol. 2021, 145, 660–674. [Google Scholar] [CrossRef]
- Kühsel, S.; Brückner, A.; Schmelzle, S.; Heethoff, M.; Blüthgen, N. Surface Area-Volume Ratios in Insects: Surface Area Allometry and Water Loss. Insect Sci. 2017, 24, 829–841. [Google Scholar] [CrossRef] [PubMed]
- Remmert, H. Body Size of Terrestrial Arthropods and Biomass of Their Populations in Relation to the Abiotic Parameters of Their Milieu. Oecologia 1981, 50, 12–13. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km Spatial Resolution Climate Surfaces for Global Land Areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10.2.2; Environmental Systems Research Institute: Redlands, CA, USA, 2014. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2018. [Google Scholar]
- Rivero, A.; West, S.A. The Physiological Costs of Being Small in a Parasitic Wasp. Evol. Ecol. Res. 2002, 4, 407–420. [Google Scholar]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Revisiting Water Loss in Insects: A Large Scale View. J. Insect Physiol. 2001, 47, 1377–1388. [Google Scholar] [CrossRef]
- Dillon, M.E. Into Thin Air: Physiology and Evolution of Alpine Insects. Integr. Comp. Biol. 2006, 46, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Symonds, M.R.E.; Elgar, M.A. The Evolution of Body Size, Antennal Size and Host Use in Parasitoid Wasps (Hymenoptera: Chalcidoidea): A Phylogenetic Comparative Analysis. PLoS ONE 2013, 8, e78297. [Google Scholar] [CrossRef]
- Gilchrist, G.W. Plastic and Genetic Variation in Wing Loading as a Function of Temperature Within and Among Parallel Clines in Drosophila Subobscura. Integr. Comp. Biol. 2004, 44, 461–470. [Google Scholar] [CrossRef] [PubMed]




| Species-Based Method | Regression Coefficients | SE | 95% Confidence Interval | p-Value |
|---|---|---|---|---|
| Pteromalidae | ||||
| SexM | −0.1431 | 0.0334 | −0.208, 0.0778 | 2.83 × 10−5 *** |
| AMTEM | −0.0205 | 3.417 × 10−3 | −0.0291, −0.0158 | 3.59 × 10−8 *** |
| APRE | 1.449 × 10−4 | 6.262 × 10−5 | 1.253 × 10−4, 3.675 × 10−4 | 0.000116 *** |
| AMSR | 3.051 × 10−5 | 8.025 × 10−6 | −1.046 × 10−6, 3.003 × 10−5 | 0.0722 |
| Eulophidae | ||||
| SexM | −0.118 | 0.0321 | −0.018, −0.05 | 0.000262 ** |
| AMTEM | −0.01756 | 2.977 × 10−3 | 0.023, −0.0178 | 7.3 × 10−9 *** |
| APRE | 1.305 × 10−4 | 3.121 × 10−5 | 6.894 × 10−5, 1.921 × 10−4 | 3.5 × 10−5 *** |
| AMSR | 1.138 × 10−5 | 6.059 × 10−6 | −4.41 × 10−7, 2.32 × 10−5 | 0.061 |
| Assemblage-based method | ||||
| Pteromalidae | ||||
| SexM | −0.1886 | 0.0273 | −0.242, −0.135 | 2.84 × 10−11 *** |
| AMTEM | −0.0205 | 2.636 × 10−3 | −0.0256, −0.0154 | 1.33 × 10−13 *** |
| APRE | 1.499 × 10−4 | 2.773 × 10−5 | 8.945 × 10−5, 0.0002 | 3.27 × 10−7 *** |
| AMSR | 3.051 × 10−5 | 6.503 × 10−6 | 1.807 × 10−5, 4.298 × 10−5 | 4.13 × 10−6 *** |
| Eulophidae | ||||
| SexM | −0.1526 | 0.02768 | −0.2068, −0.0983 | 7.47 × 10−8 *** |
| AMTEM | −0.0149 | 2.595 × 10−3 | −0.0200, −9.792 × 10−3 | 7.36 × 10−6 *** |
| APRE | 1.267 × 10−4 | 2.777 × 10−5 | 7.079 × 10−5, 1.8309 × 10−4 | 2.38 × 10−8 *** |
| AMSR | 1.136 × 10−5 | 6.031 × 10−6 | −4.274 × 10−7, 2.317 × 10−5 | 0.0607 |
| CE | χ2 | p-Value | CE | χ2 | p-Value | ||
|---|---|---|---|---|---|---|---|
| Pteromalidae | Eulophidae | ||||||
| Halticoptera (n = 221) | Diglyphus (n = 98) | ||||||
| Altitude | 2.41 × 10−4 | 17.9192 | <0.0001 *** | Altitude | 8.5 × 10−5 | 20.6508 | <0.0001 *** |
| SexM | 0.01 | 1.5968 | 0.2077 | SexM | −0.026 | 1.1148 | 0.2937 |
| Altitude × Sex | −1.5 × 10−6 | 2.3774 | 0.1245 | Altitude × Sex | −1.27 × 10−5 | 0.1044 | 0.7474 |
| Pachyneuron (n = 162) | Diaulinopsis (n = 80) | ||||||
| Altitude | 4.28 × 10−5 | 19.1234 | <0.0001 *** | Altitude | 6.85 × 10−5 | 53.949 | <0.0001 *** |
| SexM | −3.96 | 16.3063 | 0.0001 *** | SexM | −0.01 | 30.326 | <0.0001 *** |
| Altitude × Sex | 1.08 × 10−3 | 2.0679 | 0.1524 | Altitude × Sex | −4 × 10−5 | 10.134 | 0.0021 ** |
| Sphegigaster (n = 71) | Hyssopus (n = 30) | ||||||
| Altitude | 1.81 × 10−5 | 16.2064 | 0.0001 *** | Altitude | 8.16 × 10−5 | 39.0975 | <0.0001 *** |
| SexM | −0.58 | 16.1733 | 0.0001 *** | SexM | 0.227 | 1.0853 | 0.3071 |
| Altitude × Sex | 1.45 × 10−4 | 3.7199 | 0.058 | Altitude × Sex | 5.43 × 10−5 | 1.5337 | 0.2266 |
| Asaphes (n = 64) | Chrysocharis (n = 107) | ||||||
| Altitude | 2.13 × 10−5 | 7 | 0.0104 * | Altitude | −1.7 × 10−6 | 1.354 | 0.2473 |
| SexM | −0.358 | 27.5398 | <0.0001 *** | SexM | −0.061 | 2.433 | 0.1219 |
| Altitude × Sex | 8.48 × 10−5 | 8.8015 | 0.0043 ** | Altitude × Sex | 1.6 × 10−6 | 0.002 | 0.9641 |
| Pteromalus (n = 145) | Entedon (n = 150) | ||||||
| Altitude | 4.84 × 10−5 | 0.7818 | 0.3777 | Altitude | 2.02 × 10−4 | 108.014 | <0.0001 *** |
| SexM | −3.885 | 36.9243 | <0.0001 *** | SexM | −0.015 | 2.682 | 0.1037 |
| Altitude × Sex | −4.9 × 10−5 | 1.4112 | 0.2364 | Altitude × Sex | −1.39 × 10−5 | 0.089 | 0.7656 |
| Homoporus (n = 30) | Neochrysocharis (n = 115) | ||||||
| Altitude | 7.69 × 10−5 | 3.623 | 0.0681 | Altitude | 2.7 × 10−6 | 1.0725 | 0.3026 |
| SexM | −0.6578 | 73.692 | <0.0001 *** | SexM | −1.62 | 7.7498 | 0.0063 ** |
| Altitude × Sex | −1.24 × 10−4 | 1.878 | 0.1823 | Altitude × Sex | 3.62 × 10−5 | 3.4063 | 0.0676 |
| Selderma (n = 33) | Pediobius (n = 30) | ||||||
| Altitude | 3.5 × 10−4 | 31.4179 | <0.0001 *** | Altitude | 1.18 × 10−4 | 0.2745 | 0.6047 |
| SexM | 0.1187 | 7.2794 | 0.0115 * | SexM | −0.397 | 7.1793 | 0.0126 * |
| Altitude × Sex | −1.15 × 10−4 | 1.497 | 0.231 | Altitude × Sex | 8.37 × 10−5 | 0.1210 | 0.7308 |
| Sphaeripalpus (n = 34) | Aprostocetus (n = 356) | ||||||
| Altitude | 3.5 × 10−4 | 12.5613 | 0.0012 ** | Altitude | 6.23 × 10−5 | 20.992 | <0.0001 *** |
| SexM | 0.0048 | 1.8255 | 0.1859 | SexM | −0.117 | 19.163 | <0.0001 *** |
| Altitude × Sex | −2.82 × 10−5 | 0.067 | 0.7973 | Altitude × Sex | −2.13 × 10−5 | 0.739 | 0.3904 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, N.; Hu, H.; Huang, Z.; Luo, S.; Guo, S. Environmental Factors Drive Chalcid Body Size Increases with Altitudinal Gradients for Two Hyper-Diverse Taxa. Insects 2023, 14, 67. https://doi.org/10.3390/insects14010067
Kang N, Hu H, Huang Z, Luo S, Guo S. Environmental Factors Drive Chalcid Body Size Increases with Altitudinal Gradients for Two Hyper-Diverse Taxa. Insects. 2023; 14(1):67. https://doi.org/10.3390/insects14010067
Chicago/Turabian StyleKang, Ning, Hongying Hu, Zengqian Huang, Shungang Luo, and Shuhan Guo. 2023. "Environmental Factors Drive Chalcid Body Size Increases with Altitudinal Gradients for Two Hyper-Diverse Taxa" Insects 14, no. 1: 67. https://doi.org/10.3390/insects14010067