
Spatial Distribution Pattern and Sampling Plans for Two Sympatric Tomicus Species Infesting Pinus yunnanensis during the Shoot-Feeding Phase

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Field Sampling
2.3. Statistical Analysis
2.4. Fixed-Precision Sequential Sampling Plan
2.5. Model Validation
3. Results
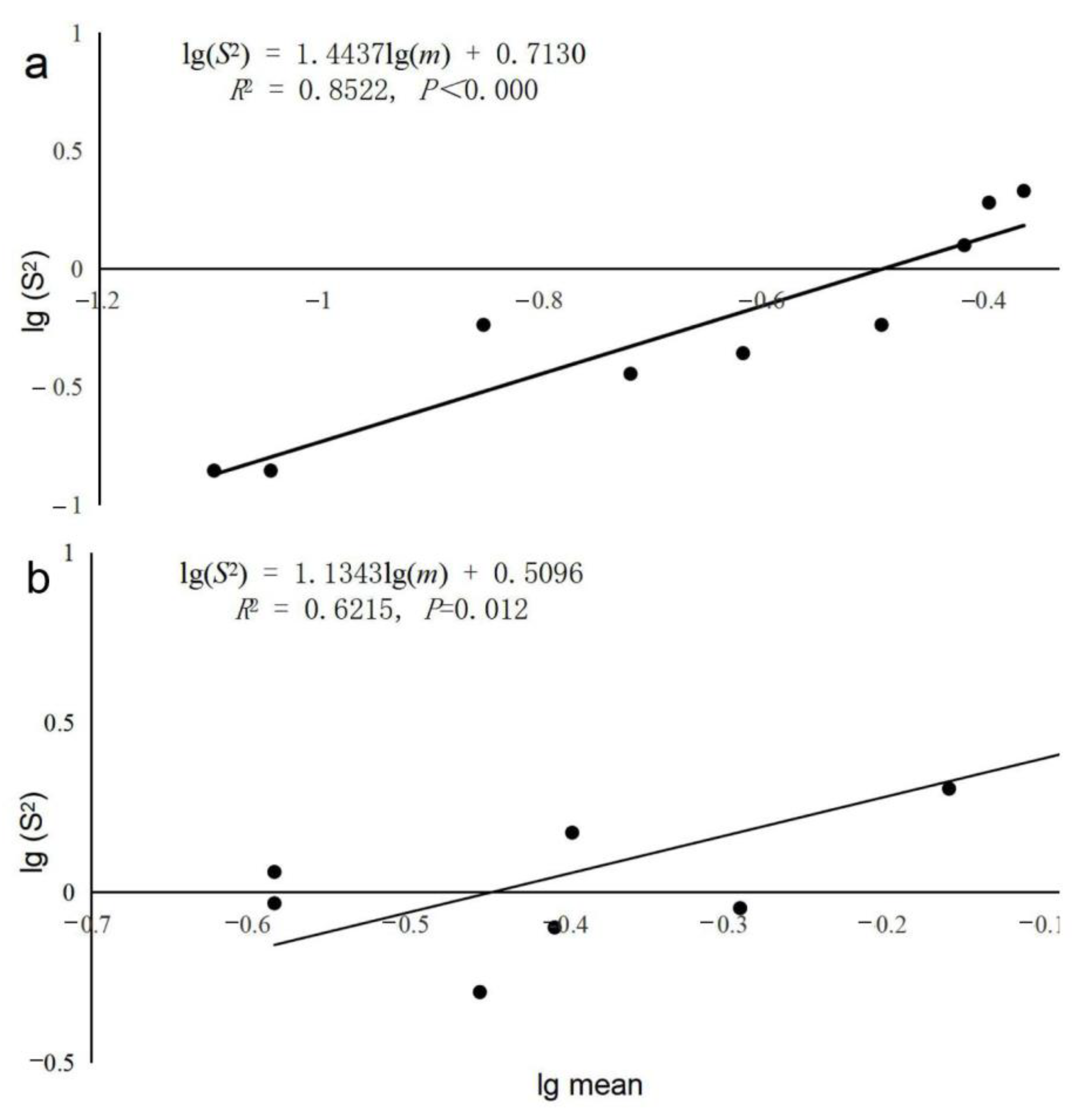
3.1. Spatial Distribution of Two Sympatric Tomicus spp.
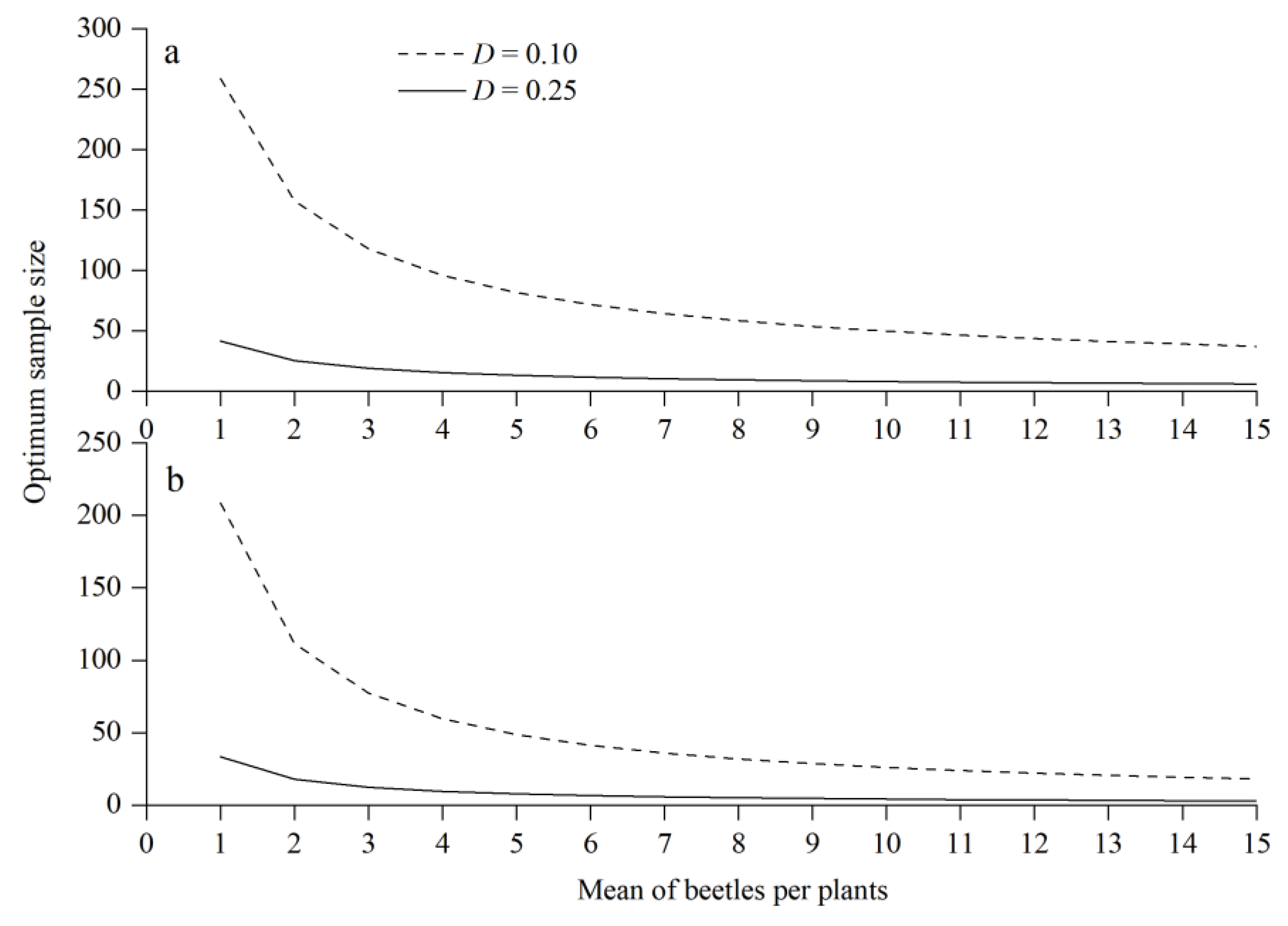
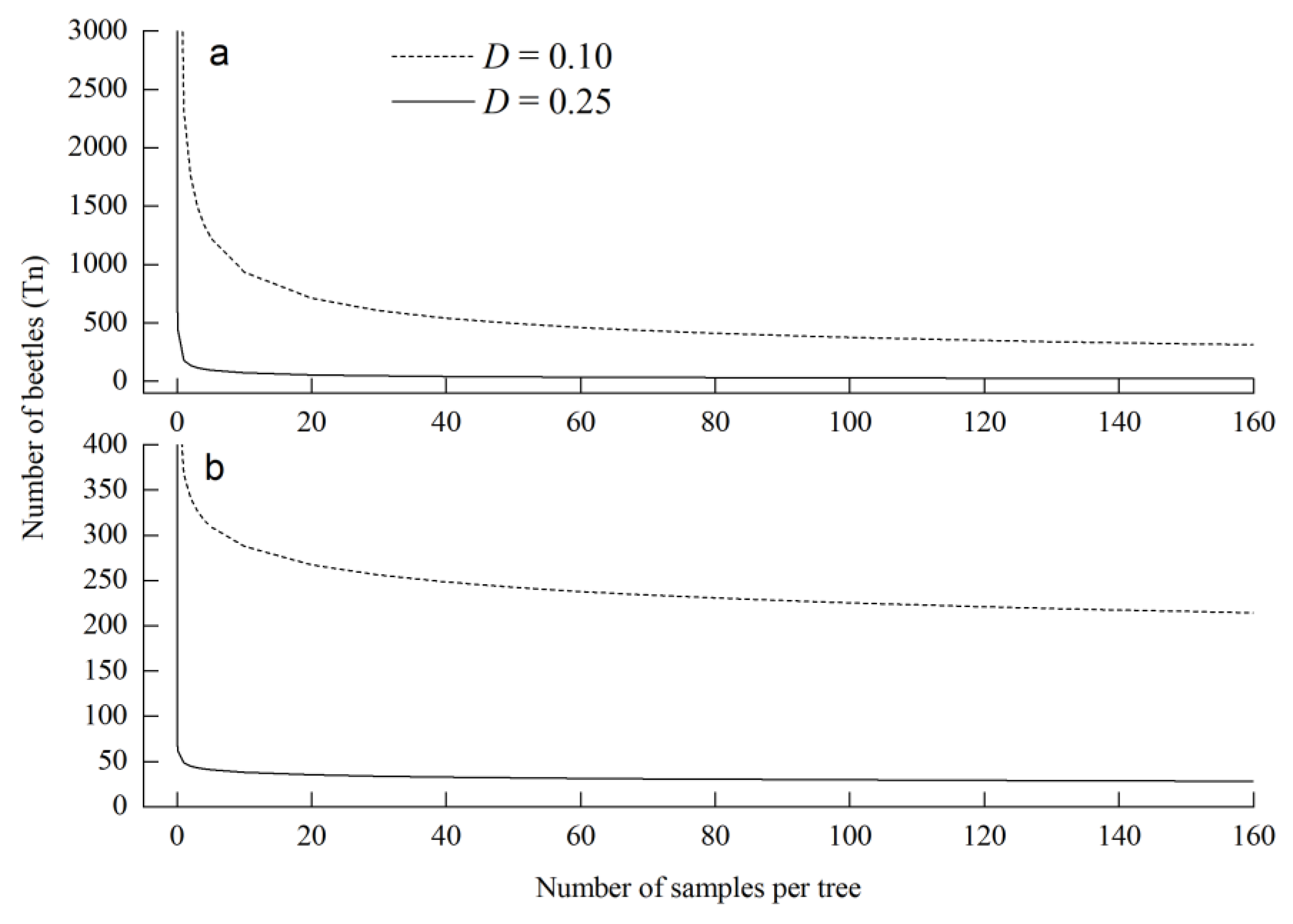
3.2. Fixed-Precision Sequential Sampling
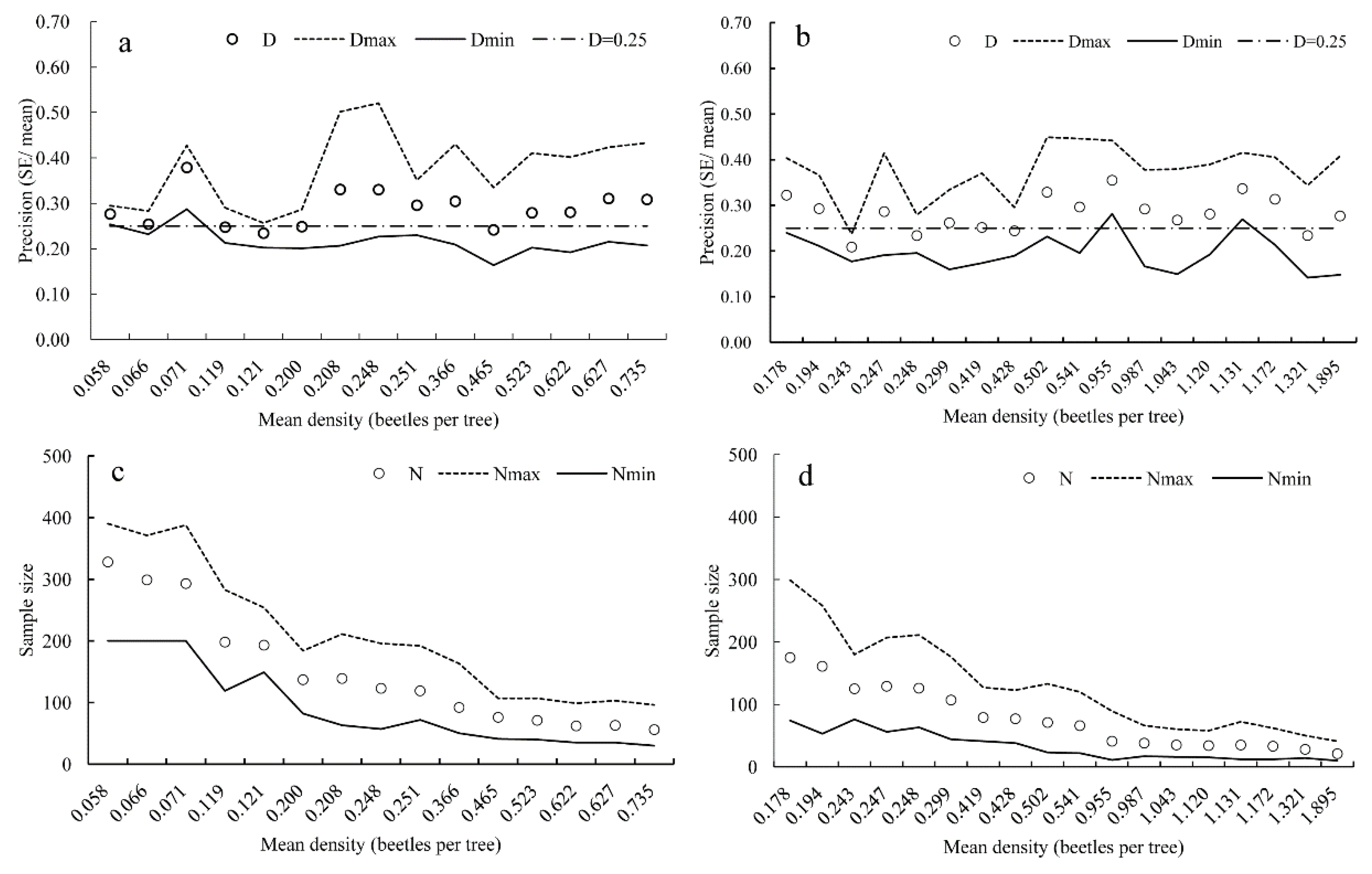
3.3. Validation of Developed Sampling Plan
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirkendall, L.R.; Faccoli, M.; Ye, H. Description of the Yunnan shoot borer, Tomicus yunnanensis Kirkendall & Faccoli sp. n. (Curculionidae, Scolytinae), an unusually aggressive pine shoot beetle from southern China, with a key to the species of Tomicus. Zootaxa 2008, 1819, 25–39. [Google Scholar]
- Li, X.; Zhang, Z.; Wang, H.; Wu, W.; Cao, P.; Zhang, P. Tomicus armandii Li & Zhang (Curculionidae, Scolytinae), a new pine shoot borer from China. Zootaxa 2010, 2572, 57–64. [Google Scholar]
- Lieutier, F.; Långström, B.; Faccoli, M. The Genus Tomicus. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2015; pp. 371–426. [Google Scholar]
- Långström, B.; Hellqvist, C. Spatial distribution of crown damage and growth losses caused by recurrent attacks of pine shoot beetles in pine stands surrounding a pulpmill in southern Sweden. J. Appl. Entomol. 1990, 110, 261–269. [Google Scholar] [CrossRef]
- Långström, B.; Hellqvist, C. Shoot damage and growth losses following three years of Tomicus attack in Scots pine stand close to a timber storage site. Silva Fenn. 1991, 25, 133–145. [Google Scholar] [CrossRef]
- Czokajlo, D.; Wink, R.A.; Warren, J.C.; Teale, S.A. Growth reduction of Scots pine, Pinus sylvestris, caused by the larger pine shoot beetle, Tomicus piniperda (Coleoptera, Scolytidae), in New York state. Can. J. For. Res. 1997, 27, 1394–1397. [Google Scholar] [CrossRef]
- Borkowski, A. Threats to pine stands by the pine shoot beetles Tomicus piniperda (L.) and T. minor (Hart.) around a sawmill in southern Poland. J. Appl. Entomol. 2001, 125, 489–492. [Google Scholar] [CrossRef]
- Borkowski, A. A spatial distribution of losses in growth of trees caused by feeding of pine shoot beetles Tomicus piniperda and T. minor (Col., Scolytidae) in Scots pine stands growing within the range of influence of a timber yard in southern Poland. J. For. Sci. 2006, 52, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Wood, S.L.; Bright, D.E. A catalogue of Scolytidae and Platypodidae (Coleoptera), part 2: Taxonomic index. Great Basin Nat. Mem. 1992, 13, 1–1553. [Google Scholar]
- Duan, Y.; Kerdelhué, C.; Ye, H.; Lieutier, F. Genetic study of the forest pest Tomicus piniperda (Col., Scolytinae) in Yunnan province (China) compared to Europe: New insights for the systematics and evolution of the genus Tomicus. Heredity 2004, 93, 416. [Google Scholar] [CrossRef] [Green Version]
- Ye, H.; Ding, X.S. Impacts of Tomicus minor on distribution and reproduction of Tomicus piniperda (Col., Scolytidae) on the trunk of the living Pinus yunnanensis trees. J. Appl. Entomol. 1999, 123, 329–333. [Google Scholar]
- Långström, B.; Lisha, L.; Hongpin, L.; Peng, C.; Haoran, L.; Hellqvist, C.; Lieutier, F. Shoot feeding ecology of Tomicus piniperda and T. minor (Col., Scolytidae) in southern China. J. Appl. Entomol. 2002, 126, 333–342. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Z.; Ye, H.; Wang, H.B.; Clarke, S.R.; Jun, L. Response of Tomicus yunnanensis (Coleoptera: Scolytinae) to infested and uninfested Pinus yunnanensis bolts. J. Econ. Èntomol. 2010, 103, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zhao, T.; Ye, H. The shoot-feeding ecology of three Tomicus species in Yunnan Province, southwestern China. J. Insect Sci. 2014, 14, 37. [Google Scholar] [CrossRef] [Green Version]
- Ye, H. Studies on the biology of Tomicus piniperda (L.) (Co., Scolytidae) in the shoot-feeding period (in Chinese). Acta Entomol. Sin. 1996, 39, 58–62. [Google Scholar]
- Ye, H. Mass attack by Tomicus piniperda L. (Col., Scolytidae) on Pinus yunnanensis tree in the Kunming region, southwestern China. In Proceedings: Integrating Cultural Tactics into the Management of Bark Beetle and Reforestation Pests; Gregoire, J.C., Liebhold, A.M., Day, K.R., Salom, S.M., Eds.; Forest Service General Technical Report, Vallombrosa, Italy, NE-236; U.S. Department of Agriculture: Washington, DC, USA, 1997; pp. 225–227. [Google Scholar]
- Wu, C.X.; Zang, L.P.; Zhang, S.F.; Kong, X.B.; Liu, F.; Zhang, Z.; Li, Y.; Xu, F.L.; Huang, G.Y. Spatial distribution patterns of three sympatric Tomicus species initially infecting Pinus yunnanensis trunks. Acta Ecol. Sin. 2020, 40, 3646–3655. [Google Scholar]
- Wu, C.X.; Liu, F.; Kong, X.B.; Zhang, S.F.; Zhang, Z. Spatiotemporal niche of competition and coexistence of three Tomicus spp. Infesting Pinus yunnanensis during the transferring stage from shoots to trunk. Sci. Silvae Sin. 2020, 56, 90–99. [Google Scholar]
- Milinski, M.; Parker, G.A. Competition for Resources. Behavioural Ecology, Evolutionary Approach, 3rd ed.; Krebs, J.R., Davies, N.B., Eds.; Blackwell Scientific Publications: London, UK, 1991; pp. 137–168. [Google Scholar]
- Southwood, T.R.E. Ecological methods with particular reference to the study of insect populations. Ecology 1979, 60, 1290–1291. [Google Scholar]
- Namvar, P.; Safaralizadeh, M.H.; Baniameri, V.; Pourmiza, A.A.; Karimzadeh Isfahani, J. Spatial distribution and fixed-precision sequential sampling of Liriomyza sativae Blanchard (Diptera: Agromyzidae) on cucumber greenhouse. Mid. East J Sci. Res. 2011, 10, 157–163. [Google Scholar]
- Southwood, T.R.E.; Henderson, P.A. Ecological Methods, 4th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2016; ISBN 9781118895252. [Google Scholar]
- Binns, M.R. Sequential sampling for classifying pest status. In Handbook of Sampling Methods for Arthropods in Agriculture; Pedigo, L.P., Buntin, G.D., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 137–174. [Google Scholar]
- Kogan, M.; Herzog, D.C. Sampling Methods in Soybean Entomology; Springer: New York, NY, USA, 1980; pp. 30–60. [Google Scholar]
- Panthi, B.R.; Renkema, J.M.; Lahiri, S.; Liburd, O.E. Spatio-temporal distribution and fixed-precision sampling plan of Scirtothrips dorsalis (Thysanoptera: Thripidae) in Florida blueberry. Insects 2021, 12, 256. [Google Scholar] [CrossRef]
- Kafeshani, F.A.; Rajabpour, A.; Aghajanzadeh, S.; Gholamian, E.; Farkhari, M. Spatial distribution and sampling plans with fixed level of precision for Citrus aphids (Hom., Aphididae) on two orange species. J. Econ. Entomol. 2018, 111, 931–941. [Google Scholar]
- Sileshi, G.J.; Baumgaertner, J.; Sithanantham, S.; Ogol, C.K.P.O. Spatial distribution and sampling plans for Mesoplatys ochroptera (Coleoptera: Chrysomelidae) on sesbania. J. Econ. Entomol. 2002, 95, 499–506. [Google Scholar] [CrossRef]
- Taylor, L.R. Aggregation, variance and the mean. Nature 1961, 189, 732–735. [Google Scholar] [CrossRef]
- Taylor, L.R. Assessing and interpreting the spatial distribution of insect populations. Annu. Rev. Entomol. 1984, 29, 321–357. [Google Scholar] [CrossRef]
- Hutchison, W.D. Sequential sampling to determine population density. In Handbook of Sampling Methods for Arthropods in Agriculture; Pedigo, L.P., Buntin, G.D., Eds.; CRC Press: Boca Raton, FL, USA, 1994; pp. 207–244. [Google Scholar]
- Southwood, T.R.E. Ecological Methods, with Particular Reference to the Study of Insect Population; Chapman and Hall: London, UK, 1995. [Google Scholar]
- Green, R.H. On fixed precision sequential sampling. Popul. Ecol. 1970, 12, 249–251. [Google Scholar] [CrossRef]
- Naranjo, S.E.; Hutchison, W.D. Validation of arthropod sampling plans using a resampling approach: Software and analysis. Am. Entomol. 1997, 43, 48–57. [Google Scholar] [CrossRef]
- Ye, H.; Li, L.S. The distribution of Tomicus piniperda (L.) population in the crown of Yunnan pine during the shoot feeding period (in Chinese). Acta Entomol. Sin. 1994, 37, 311–316. [Google Scholar]
- Chen, P.; Lu, J.; Haack, R.A.; Ye, H. Attack pattern and reproductive ecology of Tomicus brevipilosus (Coleoptera: Curculionidae) on Pinus yunnanensis in Southwestern China. J. Insect Sci. 2015, 15, 43. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.X.; Liu, F.; Zhang, S.F.; Kong, X.B.; Zhang, Z. Semiochemical regulation of the intraspecific and interspecific behavior of Tomicus yunnanensis and Tomicus minor during the shoot-feeding phase. J. Chem. Ecol. 2019, 45, 227–240. [Google Scholar] [CrossRef]
- Taylor, L.R.; Wolwod, I.P.; Perry, J.N. The density-dependence of spatial behavior and the rarity of randomness. J. Anim. Ecol. 1978, 47, 383–406. [Google Scholar] [CrossRef]
- Ye, H.; Lieutier, F. Shoot aggregation by Tomicus piniperda L (col: Scolytidae) in Yunnan, southwestern China. Ann. For. Sci. 1997, 54, 635–641. [Google Scholar]
- Lieutier, F.; Ye, H.; Yart, A. Shoot damage by Tomicus sp. (Coleoptera: Scolytidae) and effect on Pinus yunnanensis resistance to subsequent reproductive attacks in the stem. Agric. For. Entomol. 2003, 5, 227–233. [Google Scholar] [CrossRef]
- Lanne, B.S.; Schlyter, F.; Byers, J.A.; Lofqvist, J.; Leufven, A.; Bergstrom, G.; van der Pers, J.N.; Unelius, R.; Baeckstrom, P.; Norin, T. Differences in attraction to semiochemicals present in sympatric pine shoot beetles, Tomicus minor and T. piniperda. J. Chem. Ecol. 1987, 13, 1045–1067. [Google Scholar] [CrossRef] [PubMed]
- Romón, P.; Aparicio, D.; Palacios, F.; Iturrondobeitia, J.C.; Hance, T.; Goldarazena, A. Seasonal terpene variation in needles of Pinus radiata (Pinales: Pinaceae) trees attacked by Tomicus piniperda (Coleoptera: Scolytinae) and the effect of limonene on beetle aggregation. J. Insect Sci. 2017, 17, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Wu, C.; Zhang, S.; Kong, X.; Zhang, Z.; Wang, P. Initial location preference together with aggregation pheromones regulate the attack pattern of Tomicus brevipilosus (Coleoptera: Curculionidae) on Pinus kesiya. Forests 2019, 10, 156. [Google Scholar] [CrossRef] [Green Version]
- Faccoli, M.; Anfora, G.; Tasin, M. Responses of the Mediterranean pine shoot beetle Tomicus destruens (Wollaston) to pine shoot and bark volatiles. J. Chem. Ecol. 2008, 34, 1162–1169. [Google Scholar] [CrossRef]
- Shahbi, M.; Rajabpour, A. A fixed-precision sequential sampling plan for the potato tuberworm moth, Phthorimaea operculella Zeller (Lepidoptera: Gelechidae), on potato cultivars. Neotrop. Entomol. 2017, 46, 388–395. [Google Scholar] [CrossRef]
- Borden, J.H. Semiochemical-mediated aggregation and dispersion in the Coleoptera. In Insect Communication; Lewis, T., Ed.; Academic Press: London, UK, 1984; pp. 123–149. [Google Scholar]
- Wood, D.L.; Akers, R.P.; Owen, D.R.; Parmeter, J.R., Jr. The behaviour of bark beetles colonizing ponderosa pine. In Insects and the Plant Surface; Juniper, B., Southwood, R., Eds.; Edward Arnold: London, UK, 1986; pp. 91–103. [Google Scholar]
- Wang, J.; Zhang, Z.; Kong, X.; Wang, H.; Zhang, S. Intraspecific and interspecific attraction of three Tomicus beetle species during the shoot-feeding phase. Bull. Entomol. Res. 2015, 105, 225–233. [Google Scholar] [CrossRef]
- Raffa, K.F.; Berryman, A.A. The role of host plant resistance in the colonization behaviour and ecology of bark beetles (Coleoptera: Scolytidae). Ecol. Monogr. 1983, 53, 27–49. [Google Scholar] [CrossRef] [Green Version]
- Kapatos, E.T.; Stratopoulou, E.T.; Sahinoglu, A.; Tsitsipis, J.A.; Lycouresis, D.P. Development of an optimum sampling plan for population of Aphis gossypii (Hom., Aphididae) on cotton in Greece. J. Appl. Entomol. 1996, 120, 245–248. [Google Scholar] [CrossRef]
- Borkowski, A. Spatial distribution of fallen shoots of Scots pine pruned by pine shoot beetles (Tomicus spp.), and evaluation of methods of shoot collection in central Poland. J. Forest Res. 2007, 12, 358–364. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Data Sets | EXP | Yr | Num. | Tomicus minor | Tomicus yunnanensis | ||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Variance S2 | Mean | SE | Variance S2 | ||||
| 1 | A | 2016 | 100 | 0.38 | 0.11 | 1.26 | 0.51 | 0.09 | 0.90 |
| 2 | B | 2016 | 100 | 0.08 | 0.04 | 0.14 | 0.26 | 0.11 | 1.15 |
| 3 | C | 2016 | 100 | 0.40 | 0.14 | 1.91 | 0.26 | 0.10 | 0.93 |
| 4 | A | 2017 | 100 | 0.14 | 0.08 | 0.58 | 0.39 | 0.09 | 0.79 |
| 5 | B | 2017 | 100 | 0.09 | 0.04 | 0.14 | 0.40 | 0.12 | 1.50 |
| 6 | C | 2017 | 100 | 0.19 | 0.06 | 0.36 | 0.69 | 0.14 | 2.02 |
| 7 | A | 2018 | 100 | 0.32 | 0.08 | 0.58 | 0.86 | 0.19 | 3.46 |
| 8 | B | 2018 | 100 | 0.24 | 0.07 | 0.44 | 0.35 | 0.07 | 0.51 |
| 9 | C | 2018 | 100 | 0.43 | 0.15 | 2.14 | 0.85 | 0.19 | 3.54 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, C.; Chen, S.; Yang, M.; Zhang, Z. Spatial Distribution Pattern and Sampling Plans for Two Sympatric Tomicus Species Infesting Pinus yunnanensis during the Shoot-Feeding Phase. Insects 2023, 14, 60. https://doi.org/10.3390/insects14010060
Wu C, Chen S, Yang M, Zhang Z. Spatial Distribution Pattern and Sampling Plans for Two Sympatric Tomicus Species Infesting Pinus yunnanensis during the Shoot-Feeding Phase. Insects. 2023; 14(1):60. https://doi.org/10.3390/insects14010060
Chicago/Turabian StyleWu, Chengxu, Siyu Chen, Maofa Yang, and Zhen Zhang. 2023. "Spatial Distribution Pattern and Sampling Plans for Two Sympatric Tomicus Species Infesting Pinus yunnanensis during the Shoot-Feeding Phase" Insects 14, no. 1: 60. https://doi.org/10.3390/insects14010060