
Invasive Aedes japonicus Mosquitoes Dominate the Aedes Fauna Collected with Gravid Traps in Wooster, Northeastern Ohio, USA

Abstract
:Simple Summary
Abstract
1. Background
2. Material and Methods
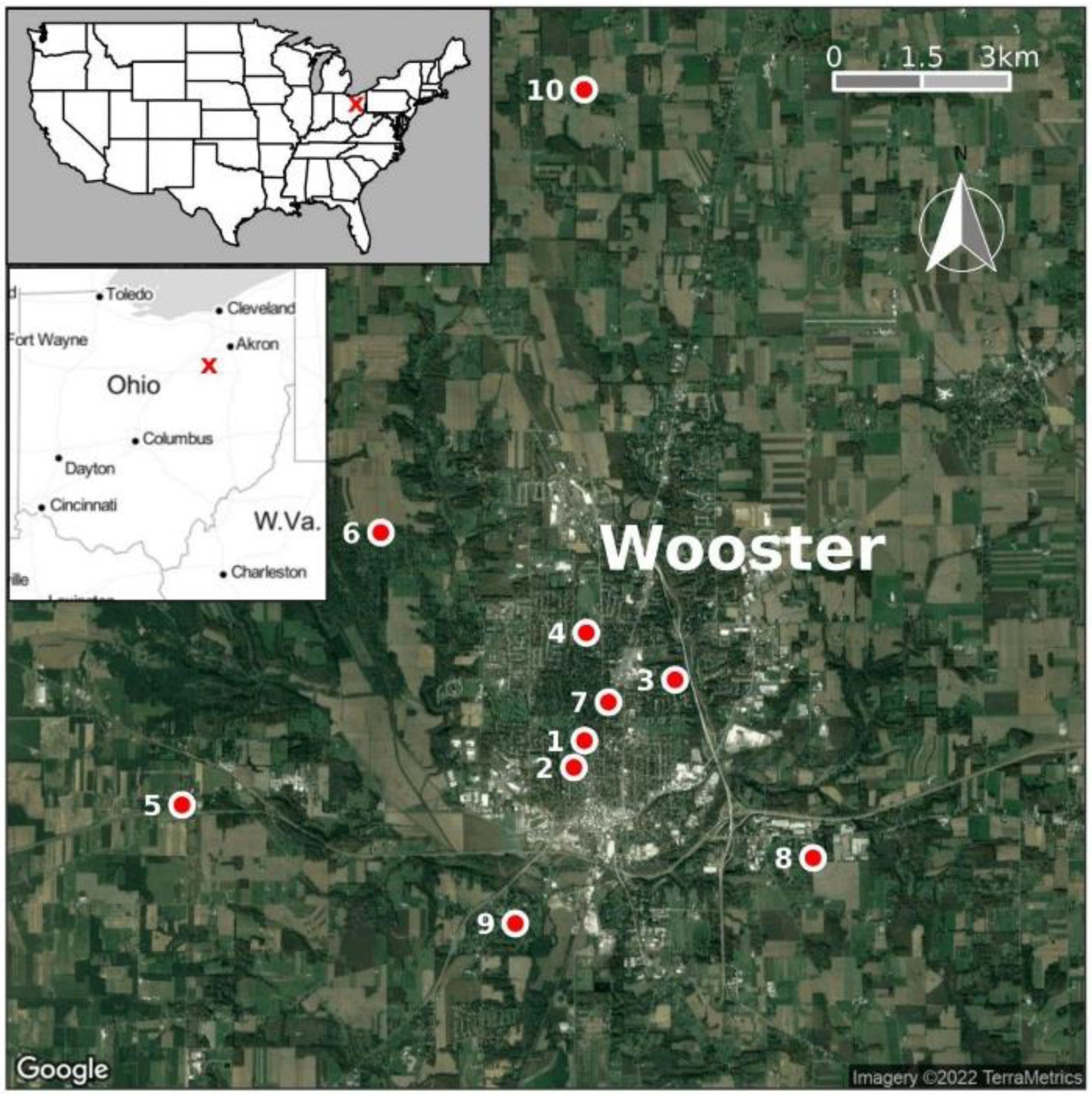
2.1. Study Site and Mosquito Collection
2.2. Mosquito Identification
2.3. DNA Isolation
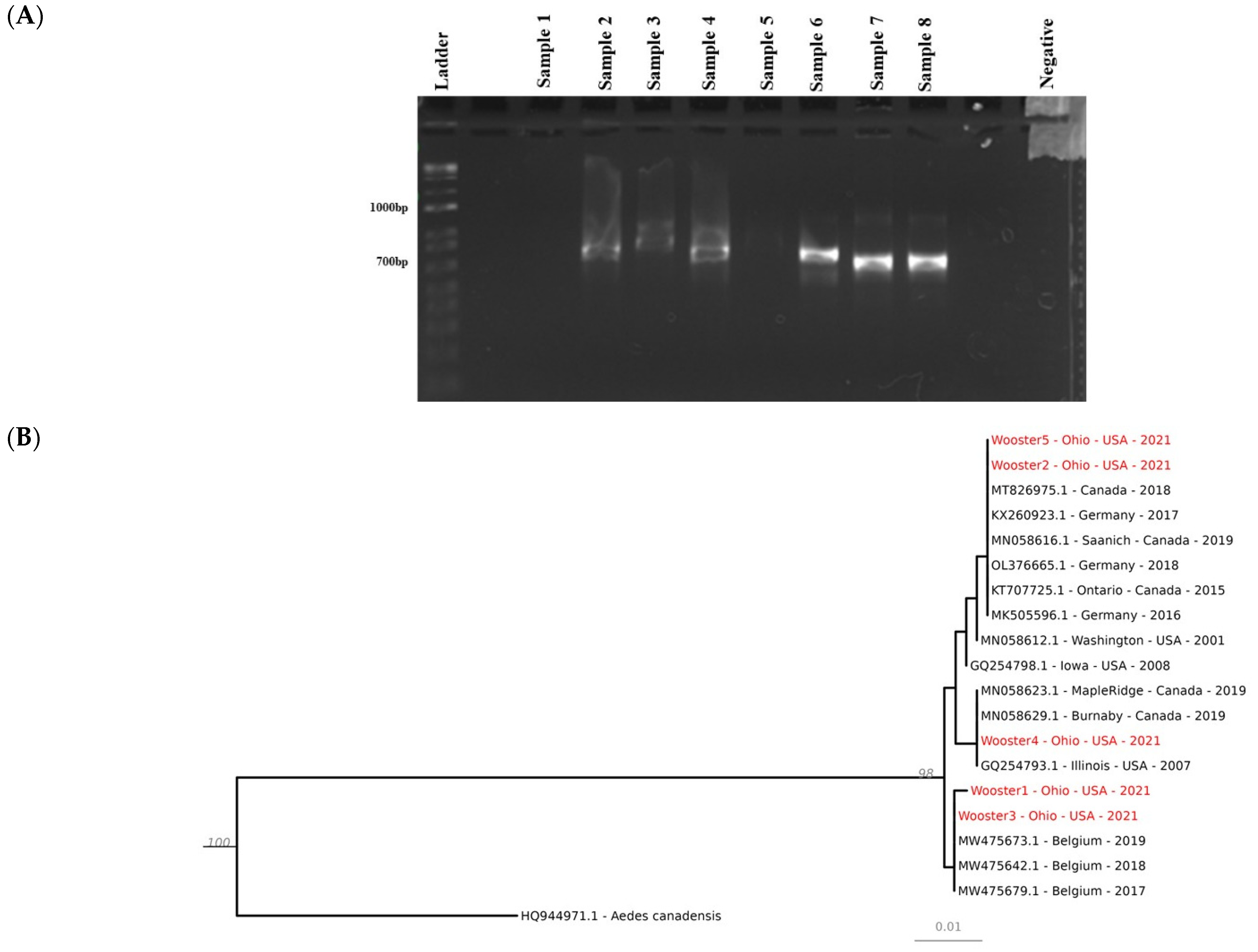
2.4. PCR, Electrophoresis, and Sanger Sequencing
2.5. Phylogenetic Analyses
3. Results
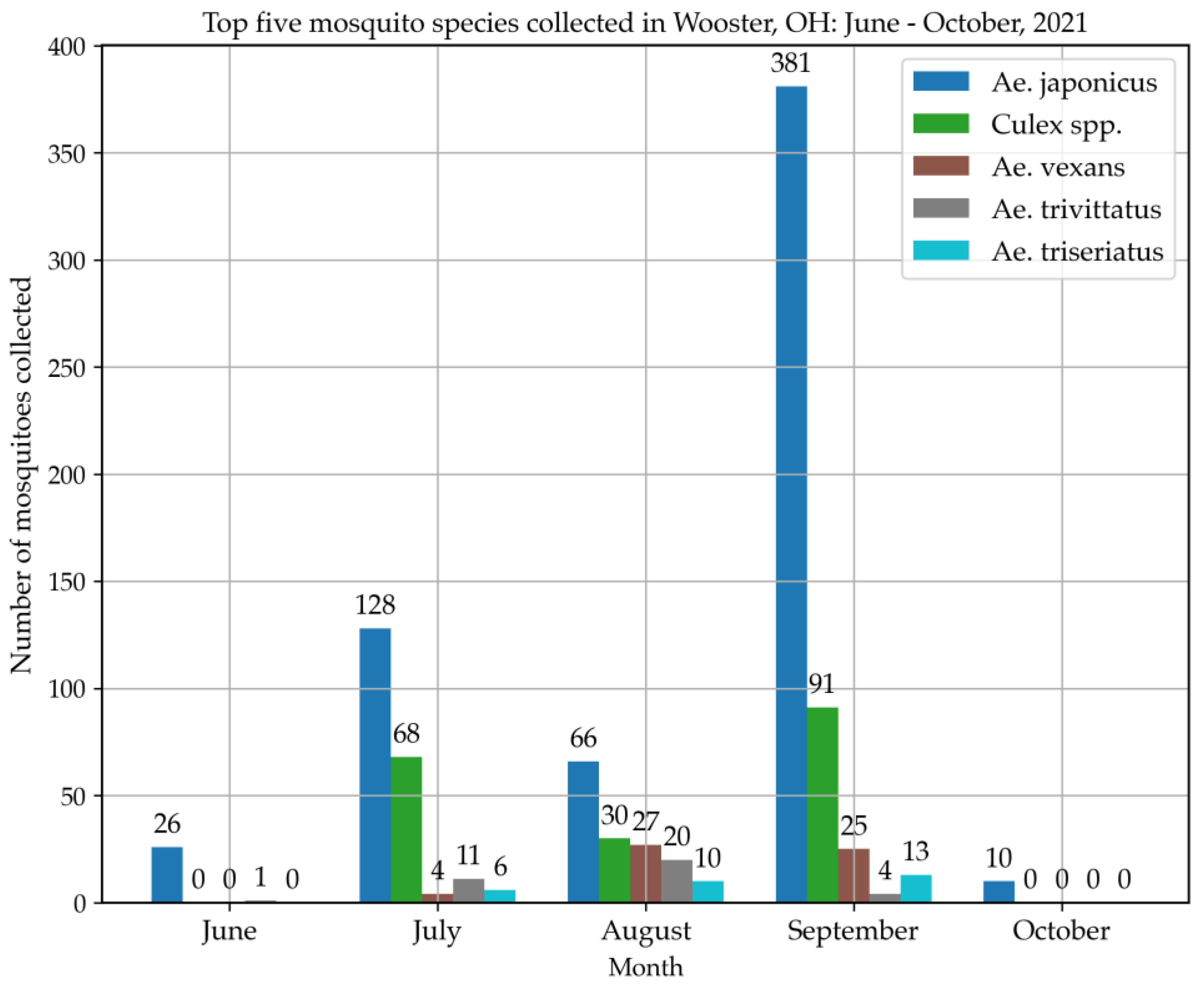
Mosquito Collections and Ae. japonicus in Wooster
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BLAST | Basic local alignment search tool |
| CDC | Centers for Disease Control |
| COI | Cytochrome oxidase subunit 1 |
| LACV | La Crosse virus |
| WNV | West Nile virus (WNV) |
References
- Montarsi, F.; Martini, S.; Michelutti, A.; Da Rold, G.; Mazzucato, M.; Qualizza, D.; Di Gennaro, D.; Di Fant, M.; Pont, M.D.; Palei, M.; et al. The invasive mosquito Aedes japonicus japonicus is spreading in northeastern Italy. Parasites Vectors 2019, 12, 120. [Google Scholar] [CrossRef] [PubMed]
- Horváth, C.; Cazan, C.D.; Mihalca, A.D. Emergence of the invasive Asian bush mosquito, Aedes (Finlaya) japonicus japonicus, in an urban area, Romania. Parasites Vectors 2021, 14, 192. [Google Scholar] [CrossRef] [PubMed]
- Andreadis, T.G.; Anderson, J.F.; Munstermann, L.E.; Wolfe, R.J.; Florin, D.A. Discovery, Distribution, and Abundance of the Newly Introduced Mosquito Ochlerotatus japonicus (Diptera: Culicidae) in Connecticut, USA. J. Med. Entomol. 2001, 38, 774–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cameron, E.C.; Wilkerson, R.C.; Mogi, M.; Miyagi, I.; Toma, T.; Kim, H.-C.; Fonseca, D.M. Molecular Phylogenetics of Aedes japonicus, a Disease Vector That Recently Invaded Western Europe, North America, and the Hawaiian Islands. J. Med. Entomol. 2010, 47, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, E.A.H.; Hutchinson, M.L.; Price, K.J.; Marini, A.; Shepard, J.J.; Molaei, G. Spatiotemporal distribution, abundance, and host interactions of two invasive vectors of arboviruses, Aedes albopictus and Aedes japonicus, in Pennsylvania, USA. Parasites Vectors 2022, 15, 36. [Google Scholar] [CrossRef] [PubMed]
- Morris, J.A.; Lampman, R.L.; Ballmes, G.; Funes, J.; Halvorsen, J.; Novak, R.J. First record of Aedes japonicus japonicus in Illinois: Defining its spatial distribution and associated mosquito species. J. Am. Mosq. Control. Assoc. 2007, 23, 243–251. [Google Scholar] [CrossRef] [PubMed]
- Kampen, H.; Werner, D. Out of the bush: The Asian bush mosquito Aedes japonicus japonicus (Theobald, 1901) (Diptera, Culicidae) becomes invasive. Parasites Vectors 2014, 7, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alto, B.W. Interspecific Larval Competition Between Invasive Aedes japonicus and Native Aedes triseriatus (Diptera: Culicidae) and Adult Longevity. J. Med. Entomol. 2011, 48, 232–242. [Google Scholar] [CrossRef]
- Kaufman, M.G.; Stanuszek, W.W.; Brouhard, E.A.; Knepper, R.G.; Walker, E.D. Establishment of Aedes japonicus japonicus and its colonization of container habitats in Michigan. J. Med. Entomol. 2012, 49, 1307–1317. [Google Scholar] [CrossRef] [Green Version]
- Sáringer-Kenyeres, M.; Bauer, N.; Kenyeres, Z. Active dispersion, habitat requirements and human biting behaviour of the invasive mosquito Aedes japonicus japonicus (Theobald, 1901) in Hungary. Parasitol. Res. 2020, 119, 403–410. [Google Scholar] [CrossRef]
- Jansen, S.; Heitmann, A.; Lühken, R.; Jöst, H.; Helms, M.; Vapalahti, O.; Schmidt-Chanasit, J.; Tannich, E. Experimental transmission of Zika virus by Aedes japonicus japonicus from southwestern Germany. Emerg. Microbes Infect. 2018, 7, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molaei, G.; Farajollahi, A.; Scott, J.J.; Gaugler, R.; Andreadis, T.G. Human Bloodfeeding by the Recently Introduced Mosquito, Aedes japonicus japonicus, and Public Health Implications. J. Am. Mosq. Control Assoc. 2009, 25, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Silaghi, C.; Beck, R.; Capelli, G.; Montarsi, F.; Mathis, A. Development of Dirofilaria immitis and Dirofilaria repens in Aedes japonicus and Aedes geniculatus. Parasites Vectors 2017, 10, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeCarlo, C.H.; Campbell, S.R.; Bigler, L.L.; Mohammed, H.O. Aedes japonicus and West Nile Virus in New York. J. Am. Mosq. Control Assoc. 2020, 36, 261–263. [Google Scholar] [CrossRef]
- Westby, K.M.; Fritzen, C.; Paulsen, D.; Poindexter, S.; Moncayo, A.C. La Crosse Encephalitis Virus Infection in Field-Collected Aedes albopictus, Aedes japonicus, and Aedes triseriatus in Tennessee. J. Am. Mosq. Control Assoc. 2015, 31, 233–241. [Google Scholar] [CrossRef]
- Martinet, J.-P.; Ferté, H.; Failloux, A.-B.; Schaffner, F.; Depaquit, J. Mosquitoes of North-Western Europe as Potential Vectors of Arboviruses: A Review. Viruses 2019, 11, 1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glavinic, U.; Varga, J.; Paslaru, A.I.; Hauri, J.; Torgerson, P.; Schaffner, F.; Veronesi, E. Assessing the role of two populations of Aedes japonicus japonicus for Zika virus transmission under a constant and a fluctuating temperature regime. Parasites Vectors 2020, 13, 479. [Google Scholar] [CrossRef]
- Kahle, D.; Wickham, H. ggmap: Spatial Visualization with ggplot2. R J. 2013, 5, 144. [Google Scholar] [CrossRef] [Green Version]
- Harrison, B.A.; Byrd, B.D.; Sither, C.B.; Whitt, P.B. The Mosquitoes of the Mid-Atlantic Region: An Identification Guide; Western Carolina University: Cullowhee, NC, USA, 2016. [Google Scholar]
- WRBU. Indentification Keys. 2021. Available online: https://www.wrbu.si.edu/index.php/vectorspecies/keys (accessed on 1 June 2021).
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T.-Y. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Escher, R.L. Sex ratios of mosquitoes from long-term censuses of Florida tree holes. J. Am. Mosq. Control. Assoc. 2008, 24, 11–15. [Google Scholar] [CrossRef] [Green Version]
- Sriwichai, P.; Karl, S.; Samung, Y.; Sumruayphol, S.; Kiattibutr, K.; Payakkapol, A.; Mueller, I.; Yan, G.; Cui, L.; Sattabongkot, J. Evaluation of CDC light traps for mosquito surveillance in a malaria endemic area on the Thai-Myanmar border. Parasites Vectors 2015, 8, 636. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, J.; Li, T.; Liu, Q.; Gong, Z.; Hou, J. Effect of different carbon dioxide (CO2) flows on trapping Aedes albopictus with BG traps in the field in Zhejiang Province, China. PLoS ONE 2020, 15, e0243061. [Google Scholar] [CrossRef]
- Harwood, J.F.; Arimoto, H.; Nunn, P.J.; Richardson, A.G.; Obenauer, P.J. Assessing Carbon Dioxide and Synthetic Lure-Baited Traps for Dengue and Chikungunya Vector Surveillance (3). J. Am. Mosq. Control Assoc. 2015, 31, 242–247. [Google Scholar] [CrossRef]
- Wilke, A.B.B.; Carvajal, A.; Medina, J.; Anderson, M.; Nieves, V.J.; Ramirez, M.; Vasquez, C.; Petrie, W.; Cardenas, G.; Beier, J.C. Assessment of the effectiveness of BG-Sentinel traps baited with CO2 and BG-Lure for the surveillance of vector mosquitoes in Miami-Dade County, Florida. PLoS ONE 2019, 14, e0212688. [Google Scholar] [CrossRef]
- Bradt, D.; Coburn, L.; Bradley, K.K.; Noden, B.H. First Record of Aedes japonicus japonicus in Oklahoma, 2017. J. Am. Mosq. Control Assoc. 2018, 34, 38–41. [Google Scholar] [CrossRef] [Green Version]
- Rowe, R.D.; Odoi, A.; Paulsen, D.; Moncayo, A.C.; Fryxell, R.T.T. Spatial-temporal clusters of host-seeking Aedes albopictus, Aedes japonicus, and Aedes triseriatus collections in a La Crosse virus endemic county (Knox County, Tennessee, USA). PLoS ONE 2020, 15, e0237322. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number (%) of Females | Number (%) of Males |
|---|---|---|
| Ae. japonicus | 590 (66.44) | 21 (26.25) |
| Ae. albopictus | 0 (0) | 1 (1.25) |
| Ae. cinereus | 5 (0.56) | 0 (0) |
| Ae. sticticus | 2 (0.23) | 0 (0) |
| Ae. triseriatus | 29 (3.27) | 1 (1.25) |
| Ae. trivittatus | 34 (3.83) | 1 (1.25) |
| Ae. vexans | 52 (5.86) | 8 (10) |
| Total Aedes | 712 (80.19) | 32 (40) |
| Culex spp. | 149 (16.78) | 40 (50) |
| An. barberi | 0 (0) | 1 (1.25) |
| An. perplexens | 1 (0.11) | 0 (0) |
| An. punctipennis | 10 (1.13) | 3 (3.75) |
| An. Quadrimaculatus sensu lato | 13 (1.46) | 3 (3.75) |
| Total Anopheles spp. | 24 (2.70) | 7 (8.75) |
| Coquillettidia perturbans | 1 (0.11) | 0 (0) |
| Orthopodomyia signifera | 1 (0.11) | 0 (0) |
| Uranotaenia sapphirina | 1 (0.11) | 1 (1.25) |
| Total number | 888 (100) | 80 (100) |
| Species | Gravid Trap | CDC Light | BG-Sentinel |
|---|---|---|---|
| Ae. japonicus | 566 | 45 | 0 |
| Ae. albopictus | 1 | 0 | 0 |
| Ae. cinereus | 5 | 0 | 0 |
| Ae. sticticus | 2 | 0 | 0 |
| Ae. triseriatus | 30 | 0 | 0 |
| Ae. trivittatus | 33 | 2 | 0 |
| Ae. vexans | 31 | 29 | 0 |
| Culex spp. | 133 | 55 | 1 |
| An. barberi | 1 | 0 | 0 |
| An. perplexens | 1 | 0 | 0 |
| An. punctipennis | 10 | 3 | 0 |
| An. quadrimaculatus | 12 | 3 | 1 |
| Coquillettidia perturbans | 1 | 0 | 0 |
| Orthopodomyia signifera | 1 | 0 | |
| Uranotaenia sapphirina | 2 | 0 | 0 |
| Total | 829 | 137 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nanfack-Minkeu, F.; Delong, A.; Luri, M.; Poelstra, J.W. Invasive Aedes japonicus Mosquitoes Dominate the Aedes Fauna Collected with Gravid Traps in Wooster, Northeastern Ohio, USA. Insects 2023, 14, 56. https://doi.org/10.3390/insects14010056
Nanfack-Minkeu F, Delong A, Luri M, Poelstra JW. Invasive Aedes japonicus Mosquitoes Dominate the Aedes Fauna Collected with Gravid Traps in Wooster, Northeastern Ohio, USA. Insects. 2023; 14(1):56. https://doi.org/10.3390/insects14010056
Chicago/Turabian StyleNanfack-Minkeu, Ferdinand, Alexander Delong, Moses Luri, and Jelmer W. Poelstra. 2023. "Invasive Aedes japonicus Mosquitoes Dominate the Aedes Fauna Collected with Gravid Traps in Wooster, Northeastern Ohio, USA" Insects 14, no. 1: 56. https://doi.org/10.3390/insects14010056