
The Role of Honey Bee Derived Aliphatic Esters in the Host-Finding Behavior of Varroa destructor

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Samples
2.2. Chemicals
2.3. Chemical Analysis
2.4. Electrotarsogram (ETG) Recordings
2.5. Y-Tube Olfactometer Bioassays
2.6. Statistical Analyses
3. Results
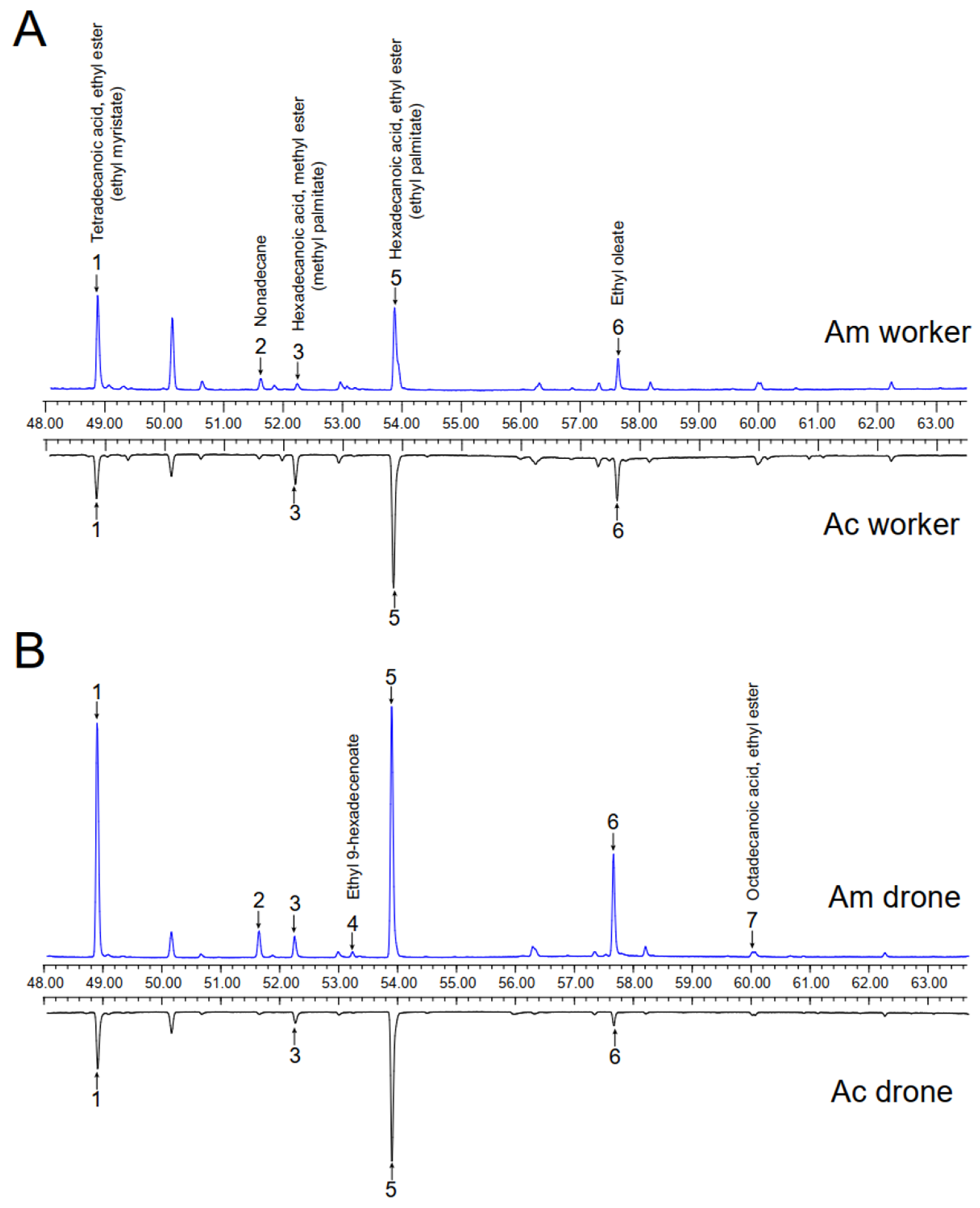
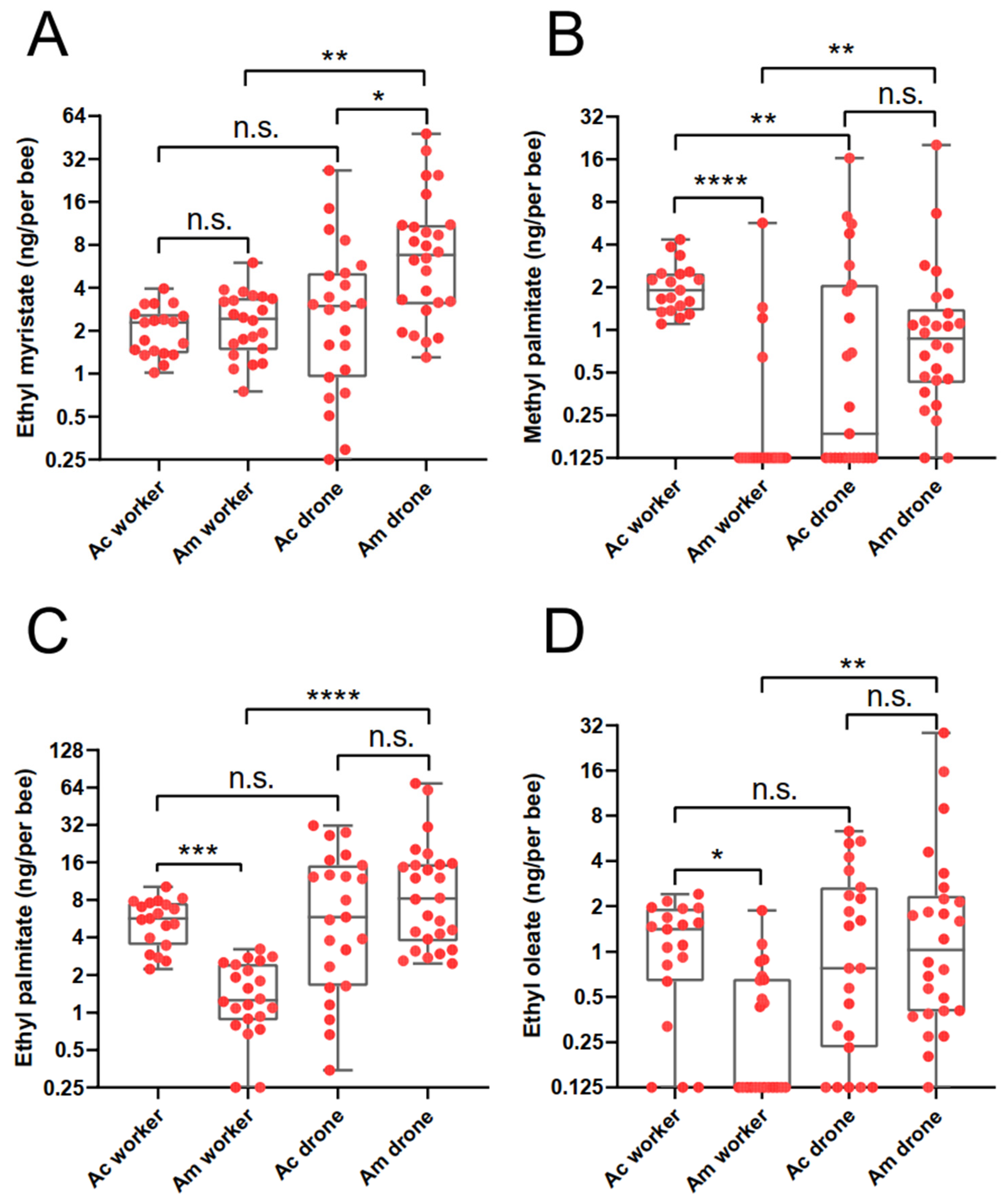
3.1. Identification and Comparison of Body Surface Volatiles of Four Kinds of Honey Bee Larvae
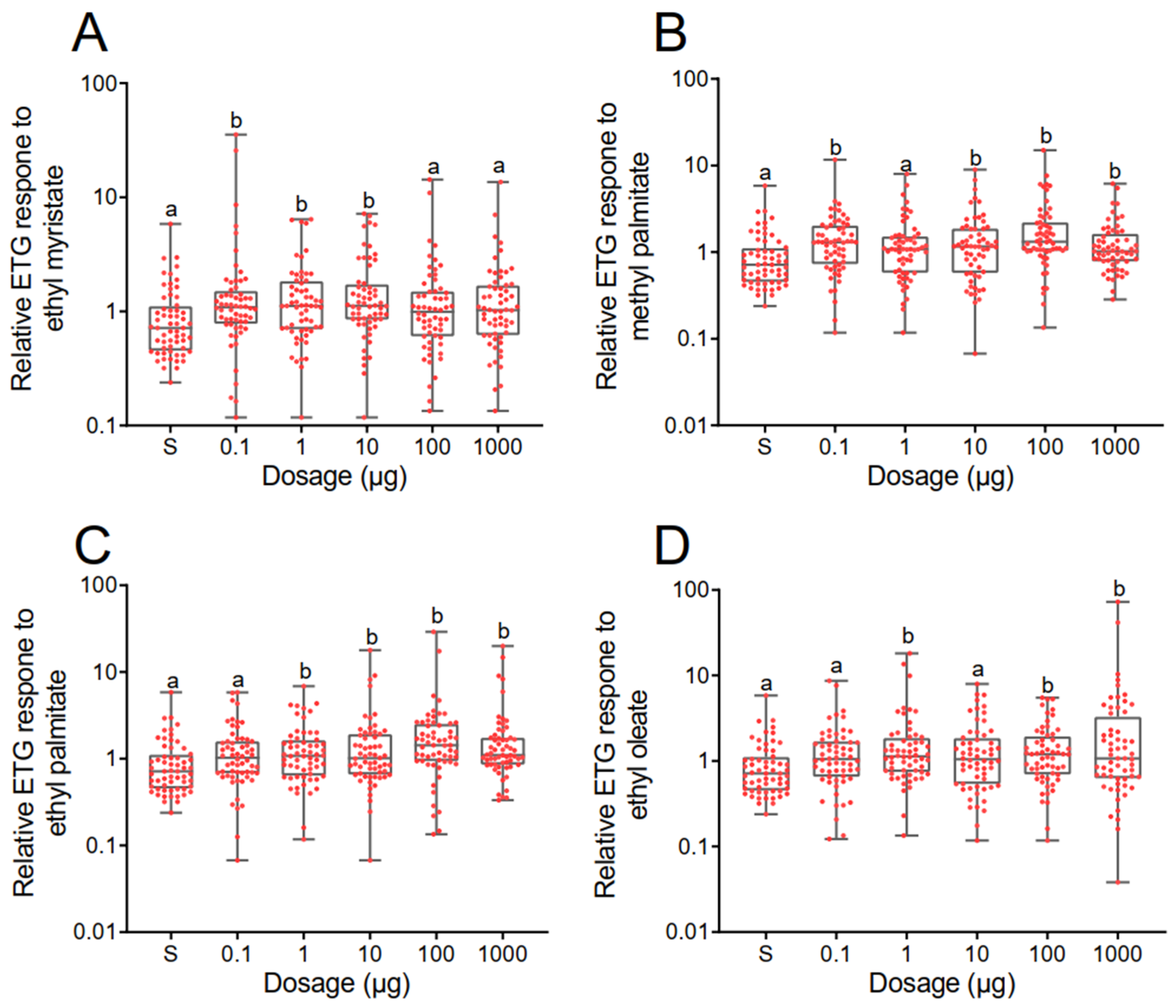
3.2. Electrophysiological Responses of Varroa destructor to the Identified Aliphatic Esters
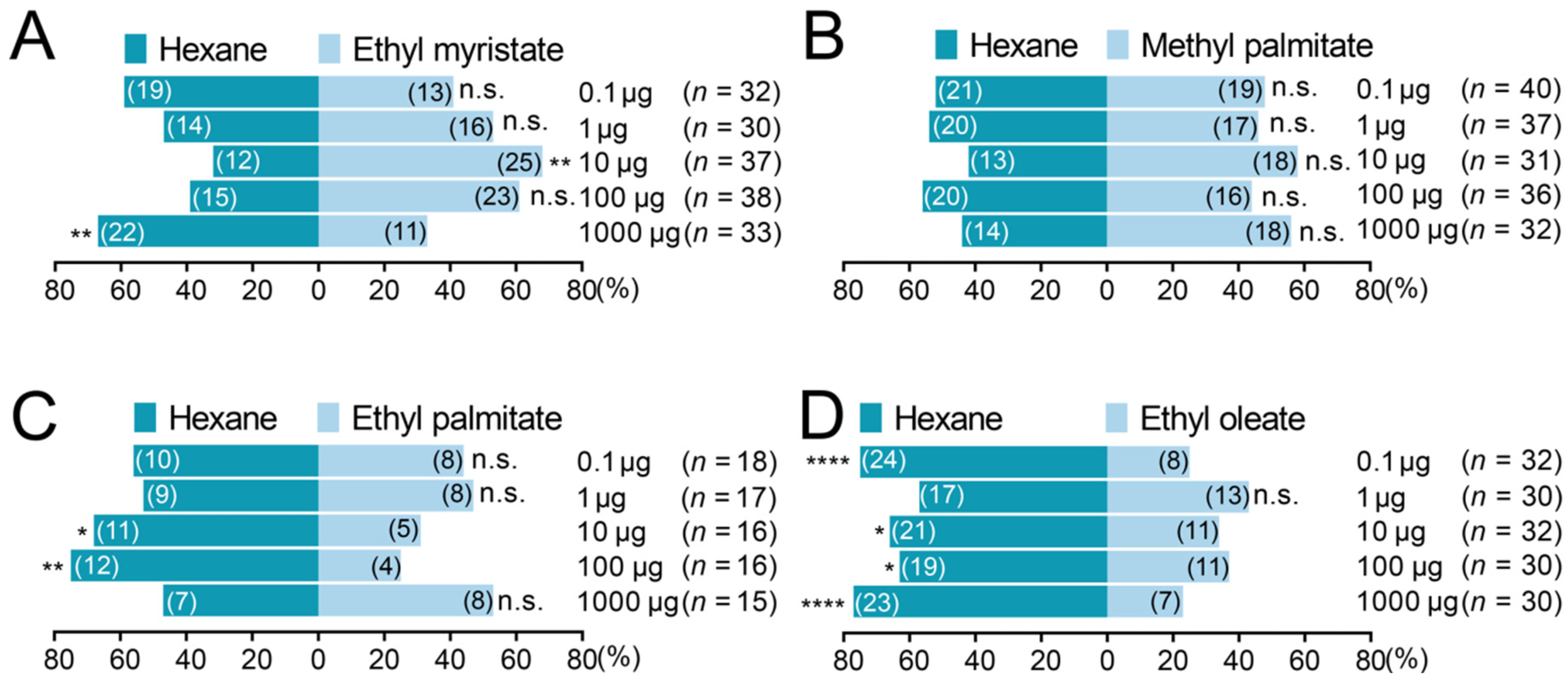
3.3. Olfactory Responses of Varroa destructor to the Identified Aliphatic Esters
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cox-Foster, D.L.; Conlan, S.; Holmes, E.C.; Palacios, G.; Evans, J.D.; Moran, N.A.; Quan, P.-L.; Briese, T.; Hornig, M.; Geiser, D.M.; et al. A metagenomic survey of microbes in honey bee colony collapse disorder. Science 2007, 318, 283–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, P.; Carreck, N.L. Honey bee colony losses. J. Apic. Res. 2010, 49, 1–6. [Google Scholar] [CrossRef] [Green Version]
- VanEngelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103, S80–S95. [Google Scholar] [CrossRef] [PubMed]
- VanEngelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony collapse disorder: A descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Ratnieks, F.L.W.; Carreck, N.L. Clarity on honey bee collapse? Science 2010, 327, 152–153. [Google Scholar] [CrossRef]
- Steinhauer, N.; Kulhanek, K.; Antúnez, K.; Human, H.; Chantawannakul, P.; Chauzat, M.-P.; vanEngelsdorp, D. Drivers of colony losses. Curr. Opin. Insect Sci. 2018, 26, 142–148. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Chen, Y. Nutrition, immunity and viral infections in honey bees. Curr. Opin. Insect Sci. 2015, 10, 170–176. [Google Scholar] [CrossRef] [Green Version]
- Traynor, K.S.; Mondet, F.; de Miranda, J.R.; Techer, M.; Kowallik, V.; Oddie, M.A.Y.; Chantawannakul, P.; McAfee, A. Varroa destructor: A complex parasite, crippling honey bees worldwide. Trends Parasitol. 2020, 36, 592–606. [Google Scholar] [CrossRef]
- Mondet, F.; Beaurepaire, A.; McAfee, A.; Locke, B.; Alaux, C.; Blanchard, S.; Danka, B.; Le Conte, Y. Honey bee survival mechanisms against the parasite Varroa destructor: A systematic review of phenotypic and genomic research efforts. Int. J. Parasitol. 2020, 50, 433–447. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef] [PubMed]
- Vilarem, C.; Piou, V.; Vogelweith, F.; Vétillard, A. Varroa destructor from the laboratory to the field: Control, biocontrol and IPM perspectives—A review. Insects 2021, 12, 800. [Google Scholar] [CrossRef] [PubMed]
- Benito-Murcia, M.; Bartolomé, C.; Maside, X.; Bernal, J.; Bernal, J.L.; del Nozal, M.J.; Meana, A.; Botías, C.; Martín-Hernández, R.; Higes, M. Residual tau-fluvalinate in honey bee colonies is coupled with evidence for selection for Varroa destructor resistance to pyrethroids. Insects 2021, 12, 731. [Google Scholar] [CrossRef]
- Hernández-Rodríguez, C.S.; Marín, Ó.; Calatayud, F.; Mahiques, M.J.; Mompó, A.; Segura, I.; Simó, E.; González-Cabrera, J. Large-scale monitoring of resistance to coumaphos, amitraz, and pyrethroids in Varroa destructor. Insects 2021, 12, 27. [Google Scholar] [CrossRef]
- Johnson, R.M.; Huang, Z.Y.; Berenbaum, M.R. Role of detoxification in Varroa destructor (Acari: Varroidae) tolerance of the miticide tau-fluvalinate. Int. J. Acarol. 2010, 36, 1–6. [Google Scholar] [CrossRef]
- Wang, R.; Liu, Z.; Dong, K.; Elzen, P.J.; Pettis, J.; Huang, Z. Association of novel mutations in a sodium channel gene with fluvalinate resistance in the mite, Varroa destructor. J. Apic. Res. 2002, 41, 17–25. [Google Scholar] [CrossRef]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the major ectoparasite of the western honey bee, Apis mellifera. Annu. Rev. Entomol. 2016, 61, 417–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor feeds primarily on honey bee fat body tissue and not hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef] [Green Version]
- Renou, M.; Anton, S. Insect olfactory communication in a complex and changing world. Curr. Opin. Insect Sci. 2020, 42, 1–7. [Google Scholar] [CrossRef]
- Ling, S.; Rizvi, S.A.H.; Xiong, T.; Liu, J.; Gu, Y.; Wang, S.; Zeng, X. Volatile signals from guava plants prime defense signaling and increase jasmonate-dependent herbivore resistance in neighboring citrus plants. Front. Plant Sci. 2022, 13, 833562. [Google Scholar] [CrossRef]
- Rizvi, S.A.H.; Ling, S.; Tian, F.; Liu, J.; Zeng, X. Interference mechanism of Sophora alopecuroides L. alkaloids extract on host finding and selection of the Asian citrus psyllid Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Environ. Sci. Pollut. Res. 2019, 26, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Giunti, G.; Palmeri, V.; Algeri, G.M.; Campolo, O. VOC emissions influence intra- and interspecific interactions among stored-product Coleoptera in paddy rice. Sci. Rep. 2018, 8, 2052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soroker, V.; Singh, N.K.; Eliash, N.; Plettner, E. Olfaction as a target for control of honeybee parasite mite Varroa destructor. In Olfactory Concepts of Insect Control—Alternative to Insecticides: Volume 1; Picimbon, J.-F., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 117–134. [Google Scholar] [CrossRef]
- Le Conte, Y.; Arnold, G.; Trouiller, J.; Masson, C.; Chappe, B.; Ourisson, G. Attraction of the parasitic mite Varroa to the drone larvae of honey bees by simple aliphatic esters. Science 1989, 245, 638–639. [Google Scholar] [CrossRef] [PubMed]
- Boot, W.J.; Schoenmaker, J.; Calis, J.N.M.; Beetsma, J. Invasion of Varroa jacobsoni into drone brood cells of the honey bee, Apis mellifera. Apidologie 1995, 26, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Calderone, N.W.; Kuenen, L.P.S. Effects of Western honey bee (Hymenoptera: Apidae) colony, cell type, and larval sex on host acquisition by female Varroa destructor (Acari: Varroidae). J. Econ. Entomol. 2001, 94, 1022–1030. [Google Scholar] [CrossRef]
- Fuchs, S. Preference for drone brood cells by Varroa jacobsoni Oud in colonies of Apis mellifera carnica. Apidologie 1990, 21, 193–199. [Google Scholar] [CrossRef]
- Le Conte, Y.; Arnold, G.; Trouiller, J.; Masson, C.; Chappe, B. Identification of a brood pheromone in honeybees. Naturwissenschaften 1990, 77, 334–336. [Google Scholar] [CrossRef]
- Zetlmeisl, K.; Rosenkranz, P. Varroa females in a bioassay: Host recognition of honey-bee larvae and adult bees. Apidologie 1994, 25, 507–508. [Google Scholar]
- Rickli, M.; Guerin, P.M.; Diehl, P.A. Palmitic acid released from honeybee worker larvae attracts the parasitic mite Varroa jacobsoni on a servosphere. Naturwissenschaften 1992, 79, 320–322. [Google Scholar] [CrossRef] [Green Version]
- Blomquist, G.J.; Bagnères, A.-G. (Eds.) Insect Hydrocarbons: Biology, Biochemistry, and Chemical Ecology; Cambridge University Press: New York, NY, USA, 2010. [Google Scholar]
- Rickli, M.; Diehl, P.A.; Guerin, P.M. Cuticle alkanes of honeybee larvae mediate arrestment of bee parasite Varroa jacobsoni. J. Chem. Ecol. 1994, 20, 2437–2453. [Google Scholar] [CrossRef]
- Aumeier, P.; Rosenkranz, P.; Francke, W. Cuticular volatiles, attractivity of worker larvae and invasion of brood cells by Varroa mites. A comparison of Africanized and European honey bees. Chemoecology 2002, 12, 65–75. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Y.; Peng, H.; Zhang, R.; Wang, Z.; Huang, Z.Y.; Chen, Y.P.; Han, R. The cell invasion preference of Varroa destructor between the original and new honey bee hosts. Int. J. Parasitol. 2022, 52, 125–134. [Google Scholar] [CrossRef]
- Lin, Z.; Qin, Y.; Page, P.; Wang, S.; Li, L.; Wen, Z.; Hu, F.; Neumann, P.; Zheng, H.; Dietemann, V. Reproduction of parasitic mites Varroa destructor in original and new honeybee hosts. Ecol. Evol. 2018, 8, 2135–2145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliash, N.; Singh, N.K.; Kamer, Y.; Pinnelli, G.R.; Plettner, E.; Soroker, V. Can we disrupt the sensing of honey bees by the bee parasite Varroa destructor? PLoS ONE 2014, 9, e106889. [Google Scholar] [CrossRef]
- Singh, N.K.; Eliash, N.; Stein, I.; Kamer, Y.; Ilia, Z.; Rafaeli, A.; Soroker, V. Identification and gene-silencing of a putative odorant receptor transcription factor in Varroa destructor: Possible role in olfaction. Insect Mol. Biol. 2016, 25, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Torto, B.; Carroll, M.; Duehl, A.; Fombong, A.; Gozansky, T.; Nazzi, F.; Soroker, V.; Teal, P. Standard methods for chemical ecology research in Apis mellifera. J. Apic. Res. 2013, 52, 1–34. [Google Scholar] [CrossRef]
- Singh, N.K.; Eliash, N.; Kamer, Y.; Zaidman, I.; Plettner, E.; Soroker, V. The effect of DEET on chemosensing of the honey bee and its parasite Varroa destructor. Apidologie 2014, 46, 380–391. [Google Scholar] [CrossRef]
- Pernal, S.F.; Baird, D.S.; Birmingham, A.L.; Higo, H.A.; Slessor, K.N.; Winston, M.L. Semiochemicals influencing the host-finding behaviour of Varroa destructor. Exp. Appl. Acarol. 2005, 37, 1–26. [Google Scholar] [CrossRef]
- Crane, E. The Varroa mite. Bee World 1978, 59, 164–167. [Google Scholar] [CrossRef]
- Xie, X.; Huang, Z.Y.; Zeng, Z. Why do Varroa mites prefer nurse bees? Sci. Rep. 2016, 6, 28228. [Google Scholar] [CrossRef]
- Salvy, M.; Martin, C.; Bagnères, A.G.; Provost, É.; Roux, M.; Le Conte, Y.; Clément, J.L. Modifications of the cuticular hydrocarbon profile of Apis mellifera worker bees in the presence of the ectoparasitic mite Varroa jacobsoni in brood cells. Parasitology 2001, 122, 145–159. [Google Scholar] [CrossRef] [PubMed]
- Rath, W. Co-adaptation of Apis cerana Fabr. and Varroa jacobsoni Oud. Apidologie 1999, 30, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Martin, C.; Salvy, M.; Provost, E.; Bagnères, A.-G.; Roux, M.; Crauser, D.; Clement, J.-L.; Le Conte, Y. Variations in chemical mimicry by the ectoparasitic mite Varroa jacobsoni according to the developmental stage of the host honey-bee Apis mellifera. Insect Biochem. Mol. Biol. 2001, 31, 365–379. [Google Scholar] [CrossRef]
- McAfee, A.; Chapman, A.; Iovinella, I.; Gallagher-Kurtzke, Y.; Collins, T.F.; Higo, H.; Madilao, L.L.; Pelosi, P.; Foster, L.J. A death pheromone, oleic acid, triggers hygienic behavior in honey bees (Apis mellifera L.). Sci. Rep. 2018, 8, 5719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maisonnasse, A.; Lenoir, J.-C.; Beslay, D.; Crauser, D.; Le Conte, Y. E-β-Ocimene, a volatile brood pheromone involved in social regulation in the honey bee colony (Apis mellifera). PLoS ONE 2010, 5, e13531. [Google Scholar] [CrossRef] [Green Version]
- Maisonnasse, A.; Lenoir, J.-C.; Costagliola, G.; Beslay, D.; Choteau, F.; Crauser, D.; Becard, J.-M.; Plettner, E.; Le Conte, Y. A scientific note on E-beta-ocimene, a new volatile primer pheromone that inhibits worker ovary development in honey bees. Apidologie 2009, 40, 562–564. [Google Scholar] [CrossRef]
- Reyes-Garcés, N.; Gionfriddo, E.; Gómez-Ríos, G.A.; Alam, M.N.; Boyacı, E.; Bojko, B.; Singh, V.; Grandy, J.; Pawliszyn, J. Advances in solid phase microextraction and perspective on future directions. Anal. Chem. 2018, 90, 302–360. [Google Scholar] [CrossRef]
- Le Conte, Y.; Sreng, L.; Poitout, S.H. Brood pheromone can modulate the feeding behavior of Apis mellifera workers (Hytnenoptera: Apidae). J. Econ. Entomol. 1995, 88, 798–804. [Google Scholar] [CrossRef]
- Le Conte, Y.; Mohammedi, A.; Robinson, G.E. Primer effects of a brood pheromone on honeybee behavioural development. Proc. R. Soc. B 2001, 268, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Mohammedi, A.; Crauser, D.; Paris, A.; Le Conte, Y. Effect of a brood pheromone on honeybee hypopharyngeal glands. Comptes Rendus De L’academie Des Sci. Ser. III Sci. De La Vie 1996, 319, 769–772. [Google Scholar]
- Tarpy, D.R.; Talley, E.; Metz, B.N. Influence of brood pheromone on honey bee colony establishment and queen replacement. J. Apic. Res. 2021, 60, 220–228. [Google Scholar] [CrossRef]
- Fuchs, S. Choice in Varroa jacobsoni Oud. between honey bee drone or workerbrood cells for reproduction. Behav. Ecol. Sociobiol. 1992, 31, 429–435. [Google Scholar] [CrossRef]
- Olsson, S.B.; Hansson, B.S. Electroantennogram and single sensillum recording in insect antennae. In Pheromone Signaling: Methods and Protocols; Touhara, K., Ed.; Humana Press: Totowa, NJ, USA, 2013; pp. 157–177. [Google Scholar]
- Koschier, E.H.; De Kogel, W.J.; Visser, J.H. Assessing the attractiveness of volatile plant compounds to western flower thrips Frankliniella occidentalis. J. Chem. Ecol. 2000, 26, 2643–2655. [Google Scholar] [CrossRef]
- Takabayashi, J.; Dicke, M. Response of predatory mites with different rearing histories to volatiles of uninfested plants. Entomol. Exp. Appl. 1992, 64, 187–193. [Google Scholar] [CrossRef]
- Boot, W.J. Methyl palmitate does not elicit invasion of honeybee brood cells by Varroa mites. Exp. Appl. Acarol. 1994, 18, 587–592. [Google Scholar] [CrossRef] [Green Version]
- Calderone, N.W.; Lin, S. Behavioural responses of Varroa destructor (Acari: Varroidae) to extracts of larvae, cocoons and brood food of worker and drone honey bees, Apis mellifera (Hymenoptera: Apidae). Physiol. Entomol. 2001, 26, 341–350. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zhang, R.; Tang, R.; Zhang, Y.; Guo, R.; Xu, G.; Chen, D.; Huang, Z.Y.; Chen, Y.; Han, R.; et al. The Role of Honey Bee Derived Aliphatic Esters in the Host-Finding Behavior of Varroa destructor. Insects 2023, 14, 24. https://doi.org/10.3390/insects14010024
Liu J, Zhang R, Tang R, Zhang Y, Guo R, Xu G, Chen D, Huang ZY, Chen Y, Han R, et al. The Role of Honey Bee Derived Aliphatic Esters in the Host-Finding Behavior of Varroa destructor. Insects. 2023; 14(1):24. https://doi.org/10.3390/insects14010024
Chicago/Turabian StyleLiu, Jiamei, Ruonan Zhang, Rui Tang, Yi Zhang, Rui Guo, Guojun Xu, Dafu Chen, Zachary Y. Huang, Yanping Chen, Richou Han, and et al. 2023. "The Role of Honey Bee Derived Aliphatic Esters in the Host-Finding Behavior of Varroa destructor" Insects 14, no. 1: 24. https://doi.org/10.3390/insects14010024