
Interspecific Competitions between Frankliniella intonsa and Frankliniella occidentalis on Fresh Lentil Bean Pods and Pepper Plants
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Experimental Design
2.2.1. The Culture of Thrips under Sexual Reproductive Mode on Fresh Lentil Bean Pods
2.2.2. The Culture of Thrips under Parthenogenetic Reproductive Mode on Fresh Lentil Bean Pods
2.2.3. The Culture of Thrips under Sexual Reproductive Mode on Pepper Plants
2.2.4. The Culture of Thrips under Parthenogenetic Reproductive Mode on Pepper Plants
2.3. Data Analysis
3. Results
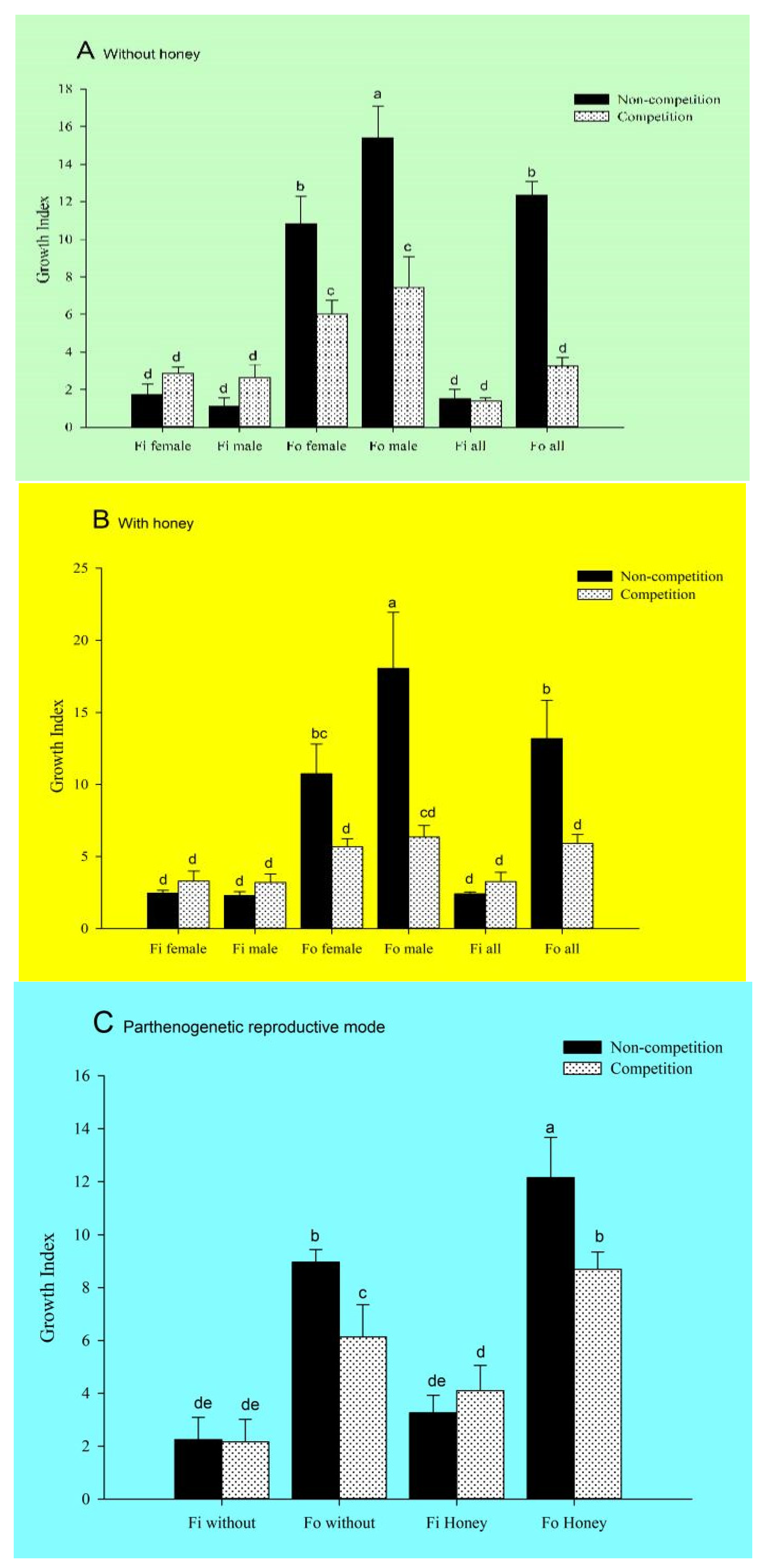
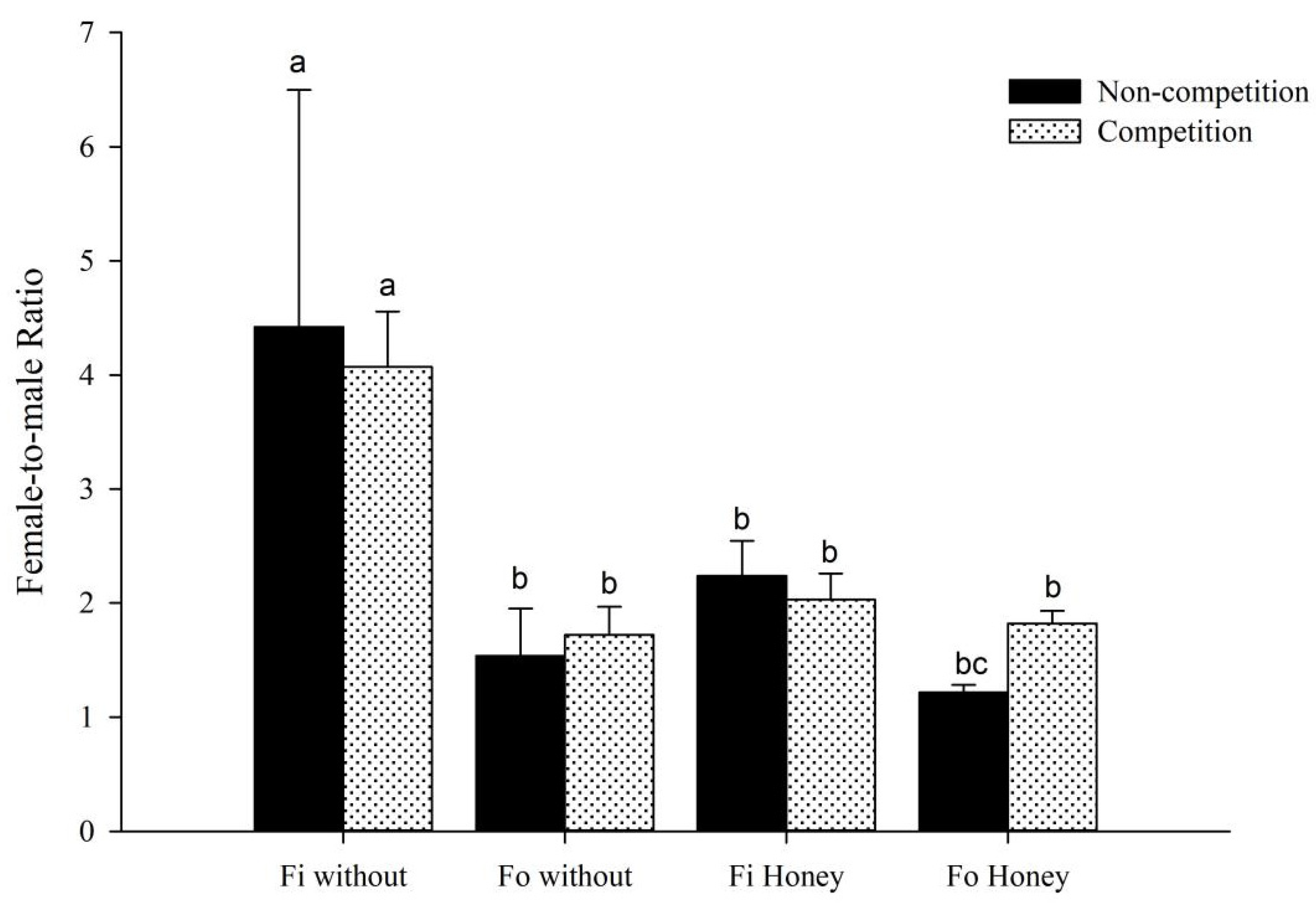
3.1. Effect of Interspecific Competitions on the Population Development of F. occidentalis and F. intonsa Cultured under Sexual Reproduction Mode on Fresh Lentil Bean Pods
3.2. Effect of Interspecific Competitions on the Population Development of F. occidentalis and F. intonsa on Fresh Lentil Bean Pods under the Parthenogenetic Reproductive Mode
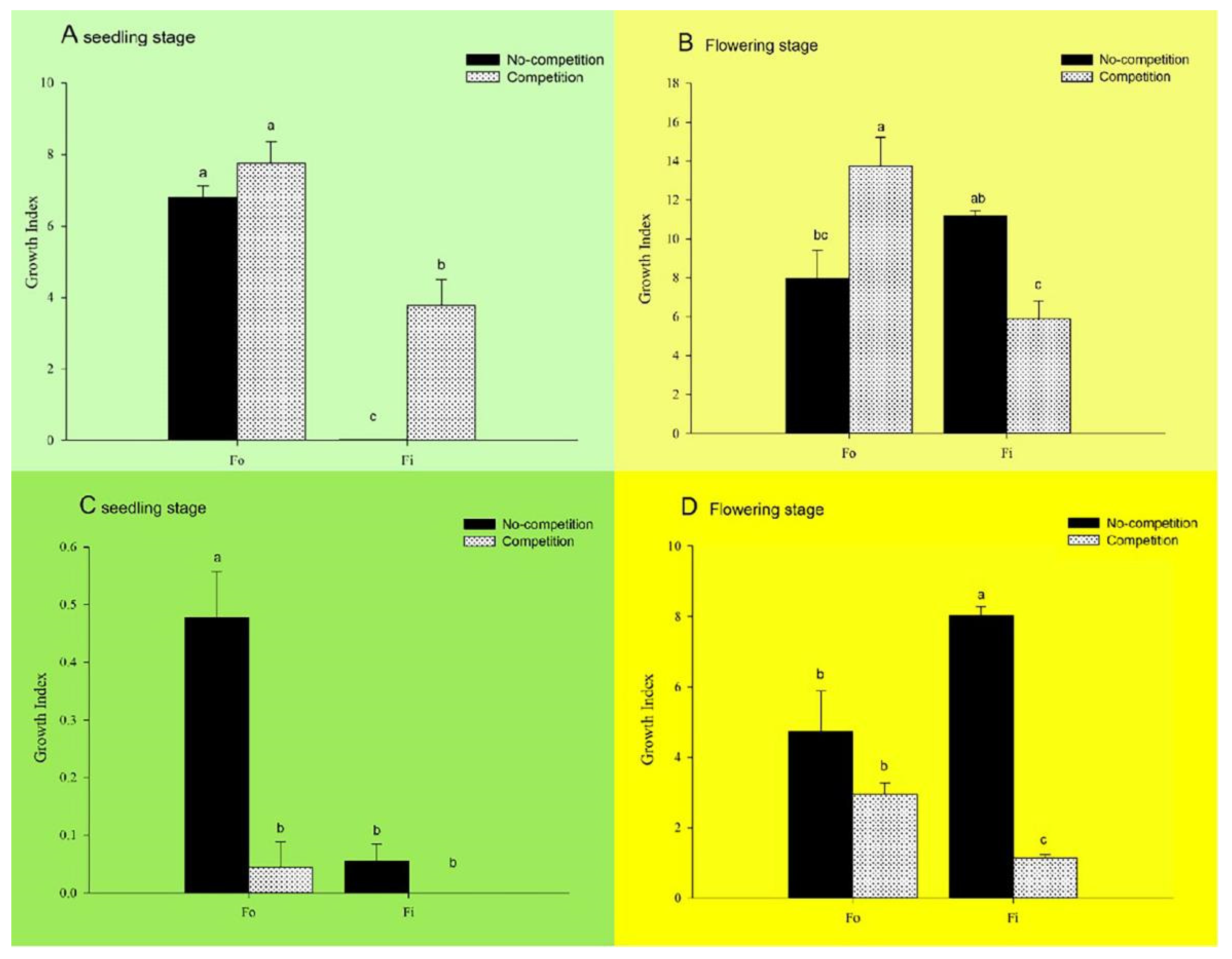
3.3. Effect of Interspecific Competitions on the Population Development of F. occidentalis and F. intonsa Cultured under Sexual Reproductive Mode on Pepper Seedlings
3.4. Effect of Interspecific Competitions on the Population Development of F. occidentalis and F. intonsa on Pepper Seedlings under Parthenogenetic Reproductive Mode
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, S.; Tang, L.; Zhang, X.; Xing, Z.; Lei, Z.; Gao, Y. A decade of a thrips invasion in China: Lessons learned. Ecotoxicology 2018, 27, 1032–1038. [Google Scholar] [CrossRef]
- Gerin, C.; Hance, T.; Impe, G.V. Demographical parameters of Frankliniella occidentalis (Pergande) (Thysanoptera, Thripidae). J. Appl. Eentomol. 1994, 118, 370–377. [Google Scholar] [CrossRef]
- Funderburk, J.; Stavisky, J.; Olson, S. Predation of Frankliniella occidentalis (Thysanoptera: Thripidae) in field peppers by Orius insidiosus (Hemiptera: Anthocoridae). Environ. Entomol. 2000, 29, 376–382. [Google Scholar] [CrossRef]
- Kirk, W.D.J.; Terry, L.I. The spread of the western flower thrips, Frankliniella occidentalis (Pergande). Agr. Forest. Entomol. 2003, 5, 301–310. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Wu, Q.J.; Xu, B.Y.; Zhu, G.R. The occurrence and damage of Frankliniella occidentalis (Thysanoptera: Thripidae): A dangerous alien invasive pest in Beijing. Plant. Protect. 2003, 29, 58–59. [Google Scholar]
- Zhang, B.; Qian, W.Q.; Qiao, X.; Xi, Y.; Wan, F.H. Invasion biology, ecology, and management of Frankliniella occidentalis in China. Arch. Insect Biochem. Physiol. 2019, 102, e21613. [Google Scholar] [CrossRef]
- Zhi, J.; Fitch, G.K.; Margolies, D.C.; Nechols, J.R. Apple pollen as a supplemental food for the western flower thrips, Frankliniella occidentalis: Response of individuals and populations. Entomol. Exp. Appl. 2005, 117, 185–192. [Google Scholar] [CrossRef]
- Frantz, G.; Mellinger, H.C. Shifts in western flower thrips, Frankliniella occidentalis (thysanoptera: Thripidae), population abundance and crop damage. Fla. Entomol. 2009, 92, 29–34. [Google Scholar] [CrossRef]
- Li, W.D.; Zhang, P.J.; Zhang, J.M.; Zhang, Z.J.; Huang, F.; Bei, Y.W.; Lin, W.C.; Lu, Y.B. An Evaluation of Frankliniella occidentalis (Thysanoptera: Thripidae) and Frankliniella intonsa (Thysanoptera: Thripidae) Performance on Different Plant Leaves Based on Life History Characteristics. J. Insect Sci. 2015, 15, 167. [Google Scholar] [CrossRef] [PubMed]
- Gai, H.T.; Zhi, J.R.; Yue, Z. Population dynamics of Frankliniella occidentalis and Frankliniella intonsa on pepper. Southwest Chin. J. Agr. Sci. 2012, 25, 337–339. [Google Scholar]
- Wu, Q.J.; Xu, B.Y.; Zhang, Z.J.; Zhang, Y.J.; Zhu, G.R. The investigation on populations and distribution of thrips in plant in Beijing, Zhejiang province and Yunnan province. Chin. Plant Protect. 2007, 27, 32–33. [Google Scholar]
- Li, J. Synthesis of A Male-produced Aggregation Pheromones in Frankliniella intonsa and Evaluation of Their Activities. Master’s Thesis, Nanjing Agricultural University, Nanjing, China, 2014. [Google Scholar]
- Moritz, G. Structure, Growth and Development: Trips as Crop Pests; CAB: New York, NY, USA, 1997; pp. 15–63. [Google Scholar]
- Ding, T.B.; Chi, H.; Gökçe, A.; Gao, Y.L.; Zhang, B. Demographic analysis of arrhenotokous parthenogenesis and bisexual reproduction of Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Sci. Rep. 2018, 8, 3346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tommasini, M.G.; Maini, S. Frankliniella occidentalis and other thrips harmful to vegetable and ornamental crops in Europe. Biol. Control. Thrips. Pests. 1995, 95, 1–42. [Google Scholar]
- Brunner, P.C.; Frey, J.E. Habitat-specific population structure in native western flower thrips Frankliniella occidentalis (insecta, thysanoptera). J. Evol. Biol. 2010, 23, 797–804. [Google Scholar] [CrossRef]
- Ullah, M.S.; Lim, U.T. Life History Characteristics of Frankliniella occidentalis and Frankliniella intonsa (Thysanoptera: Thripidae) in Constant and Fluctuating Temperatures. J. Econ. Entomol. 2015, 108, 1000–1009. [Google Scholar] [CrossRef]
- Todd, J.W.; Culbreath, A.K.; Chamberlin, J.R.; Beshear, R.J.; Mullinix, B.G. Colonization and population dynamics of thrips in peanuts in the southern United States. Thrips. Biol. Manag. 1995, 276, 453–460. [Google Scholar]
- Joost, H.; Riley, D.G. Sampling techniques for thrips (Thysanoptera: Thripidae) in pre-flowering tomato. J. Econ. Entomol. 2004, 97, 1450–1454. [Google Scholar] [CrossRef]
- Navas, S.; Funderburk, J.E.; Beshear, R.J.; Olson, S.M.; Mack, T.P. Seasonal patterns of Frankliniella (Thysanoptera:Thripidae) in tomato flowers. J. Econ. Entomol. 1991, 84, 1818–1822. [Google Scholar] [CrossRef]
- Riley, D.G.; Angelella, G.M.; Mcpherson, R.M. Pine pollen dehiscence relative to thrips population dynamics. Entomol. Exp. Appl. 2011, 138, 223–233. [Google Scholar] [CrossRef]
- Navas, V.; Funderburk, J.E.; Olson, S.M.; Beshear, R.J. Damage to tomato fruit by the western flower thrips (Thysanoptera:Thripidae). J. Entomol. Sci. 1991, 26, 436–442. [Google Scholar] [CrossRef]
- Shipp, J.L.; Wang, K.; Binns, M.R. Economic injury levels for western flower thrips (Thysanoptera: Thripidae) on greenhouse cucumber. J. Econ. Entomol. 2000, 93, 1732–1740. [Google Scholar] [CrossRef] [PubMed]
- Buxton, J.H.; Easterbrook, M.A. Thrips as a probable cause of severe fruit distortion in late-season strawberries. Plant. Pathol. 2010, 37, 278–280. [Google Scholar] [CrossRef]
- Wijkamp, J. Virus-Vector Relationships in the Transmission of Tospoviruses. Ph.D. Dissertation, University of Wageningen, Wageningen, The Netherlands, 1995. [Google Scholar]
- Sakurai, T.; Inoue, T.; Tsuda, S. Distinct efficiencies of Impatiens necrotic spot virus transmission by five thrips vector species (Thysanoptera: Thripidae) of tospoviruses in Japan. Appl. Entomol. Zool. 2004, 39, 71–78. [Google Scholar] [CrossRef] [Green Version]
- Nagata, T.; Almeida, A.C.L.; Resende, R.O.; DeAvila, A.C. The competence of four thrips species to transmit and replicate four tospoviruses. Plant Pathol. 2010, 53, 136–140. [Google Scholar] [CrossRef]
- Xie, Y.H.; Zhang, H.R.; Liu, J.; Li, Z.Y. Advances in research on vector thrips species (Thysanoptera, Thripidae). Chin. J. Appl. Entomol. 2013, 50, 1726–1736. [Google Scholar]
- Prins, M.M.; Goldbach, R. The emerging problem of tospovirus infection and nonconventional methods of control. Trends Microbiol. 1998, 6, 31–35. [Google Scholar] [CrossRef]
- Riley, D.G.; Joseph, S.V.; Srinivasan, R.; Diffie, S. Thrips vectors of tospoviruses. J. Integr. Pest Manag. 2011, 2, 11–110. [Google Scholar] [CrossRef]
- Reitz, S.R.; Gao, Y.; Kirk, W.; Hoddle, M.S.; Funderburk, J.E. Invasion biology, ecology, and management of western flower thrips. Annu. Rev. Entomol. 2020, 65, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Gai, H.T.; Zhi, J.R.; Li, Z.X.; Jiang, Y.J. Survival rates of Frankliniella occidentalis and Frankliniella intonsa after exposure to adverse temperature conditions. Chin. J. Ecol. 2010, 29, 1533–1577. [Google Scholar]
- Hu, C.X.; Li, Y.R.; Li, Z.Y.; Chen, G.H.; Zhang, H.R.; Gui, F.R.; Zhang, X.M. Effects of imidacloprid on interspecific competition between Frankliniella occidentalis and Frankliniella intonsa and their progeny development. Chin. J. Ecol. 2018, 37, 453–461. [Google Scholar]
- Wu, S.Y.; Xing, Z.L.; Ma, T.T.; Xu, D.W.; Gao, Y.L. Competitive interaction between Frankliniella occidentalis and locally present thrips species: A global review. J. Pest Sci. 2021, 94, 5–16. [Google Scholar] [CrossRef]
- Charlene, J.H.; Judith, H.M. Sex Ratio Patterns and Population Dynamics of Western Flower Thrips (Thysanoptera: Thripidae). Environ. Entomol. 1992, 21, 322–330. [Google Scholar]
- Hansen, E.A.; Funderburk, J.E.; Reitz, S.R.; Ramachandran, S.; Eger, J.E.; McAuslane, H. Within-plant distribution of Frankliniella species (Thysanoptera: Thripidae) and Orius insidiosus (Heteroptera: Anthocoridae) in field pepper. Environ. Entomol. 2003, 32, 1035–1044. [Google Scholar] [CrossRef] [Green Version]
- Gerin, C.; Hance, T.; Van-Impe, G. Impact of flowers on the demography of western flower thrips Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). J. Appl. Entomol. 1999, 123, 569–574. [Google Scholar] [CrossRef]
- Jiang, X.C.; Li, Z.H.; Cao, Z.Y.; He, S.Q.; Li, Z.Y.; Liu, J.Y.; Gui, F.R. Population dynamics and spatial distribution of thrips on vegetables flowers. Chin. J. Appl. Entomol. 2013, 50, 1628–1636. [Google Scholar]
- Paini, D.R.; Funderburk, J.E.; Reitz, S.R. Competitive exclusion of a worldwide invasive pest by a native. Quantifying competition between two phytophagous insects on two host plant species. J. Anim. Ecol. 2008, 77, 184–190. [Google Scholar] [CrossRef]
- Northfield, T.D.; Paini, D.R.; Reitz, S.R.; Funderburk, J.E. Within plant interspecific competition does not limit the highly invasive thrips, Frankliniella occidentalis in florida. Ecol. Entomol. 2011, 36, 181–187. [Google Scholar] [CrossRef]
- Sakai, A.K.; Allendorf, F.W.; Holt, J.S.; Lodge, D.M.; Molofsky, J.; With, K.A.; Baughman, S.; Cabin, R.J.; Cohen, J.E.; Ellstrand, N.C.; et al. The population biology of invasive species. Annu. Rev. Ecol. Syst. 2001, 32, 305–332. [Google Scholar] [CrossRef] [Green Version]
- Lewis, T. Pest thrips in perspective. In Thrips as Crop Pests; Lewis, T., Ed.; CAB International: New York, NY, USA, 1997; pp. 1–13. [Google Scholar]
- Gai, H.T.; Zhi, J.R.; Sun, M. Effects of temperature on the survival and fecundity of Frankliniella occidentalis and F. intonsa (Thysanoptera: Thripidae). Acta Phytophylacica Sin. 2011, 38, 521–526. [Google Scholar]
- Alim, M.A.; Song, J.; Seo, H.J.; Choi, J.J. Monitoring thrips species with yellow sticky traps in astringent persimmon orchards in Korea. Appl. Entomol. Zool. 2018, 53, 75–84. [Google Scholar] [CrossRef]
- Zhang, Z.K.; Wu, S.Y.; Lei, Z.R.; Kang, P.Z.; Du, Y.N.; Zhang, L.R. Population competition and occurrence trend of Frankliniella occidentalis and Frankliniella intonsa on Yinchuan greenhouse pepper. Plant Quar. 2019, 33, 13–17. [Google Scholar]
- Zhang, T.; Zhi, J.R.; Ye, M.; Zeng, G.; Ye, G.Q. Effect of pepper flowers on the population aboundance of Frankliniella occidentalis. China Plant Protect. 2018, 38, 35–38. [Google Scholar]
- Zhou, D.; Zhi, J.R.; Song, J.; Yue, W.B.; Zhang, T.; Zou, J. Effects of Different Foods on Nutrient Contents in Frankliniella Occidentalis. J. Mt. Agric. Biol. 2021, 40, 18–23. [Google Scholar]
- McNeill, S.; Southwood, T.R.E. Role of nitrogen in the development of insect plant relationships. In Biochemical Aspects of Plant and Animal Convolution; Harbourne, J.B., Ed.; Academic Press: New York, NY, USA, 1978; pp. 77–98. [Google Scholar]
- Mattson, W.J. Herbivory in relation to plant nitrogen content. Annu. Rev. Ecol. Syst. 1980, 102, 391–403. [Google Scholar] [CrossRef]
- Derridji, S.; Wu, B.R.; Stammitti, L.; Garrec, J.P.; Derrien, A. Chemi-calson the leaf surface: Information about the plant available to insects. Entomol. Exp. Appl. 1996, 80, 197–201. [Google Scholar] [CrossRef]
- Bhuyain, M.; Lim, U.T. Interference and Exploitation Competition between Frankliniella occidentalis and Frankliniella intonsa (Thysanoptera: Thripidae) in Laboratory Assays. Fla. Entomol. 2019, 102, 322–328. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. | Competition | Number of Fo Females | Number of Fo Males | Number of Fi Females | Number of Fi Males | Food | Sexual Reproduction | Number of Replications |
|---|---|---|---|---|---|---|---|---|
| 1 | No | 20 | 10 | 0 | 0 | lentil with honey | Yes | 4 |
| No | 20 | 10 | 0 | 0 | lentil without honey | Yes | 4 | |
| No | 0 | 0 | 20 | 10 | lentil with honey | Yes | 4 | |
| No | 0 | 0 | 20 | 10 | lentil without honey | Yes | 4 | |
| Yes | 10 | 5 | 10 | 5 | lentil with honey | Yes | 4 | |
| Yes | 10 | 5 | 10 | 5 | lentil without honey | Yes | 4 | |
| 2 | No | 20 | 0 | 0 | 0 | lentil with honey | No | 4 |
| No | 20 | 0 | 0 | 0 | lentil without honey | No | 3 | |
| No | 0 | 0 | 20 | 0 | lentil with honey | No | 3 | |
| No | 0 | 0 | 20 | 0 | lentil without honey | No | 3 | |
| Yes | 10 | 0 | 10 | 0 | lentil with honey | No | 3 | |
| Yes | 10 | 0 | 10 | 0 | lentil without honey | No | 3 | |
| 3 | No | 20 | 10 | 0 | 0 | pepper seedlings stages | Yes | 3 |
| No | 20 | 10 | 0 | 0 | pepper flowering stages | Yes | 3 | |
| No | 0 | 0 | 20 | 10 | pepper seedlings stages | Yes | 3 | |
| No | 0 | 0 | 20 | 10 | pepper flowering stages | Yes | 3 | |
| Yes | 10 | 5 | 10 | 5 | pepper seedlings stages | Yes | 3 | |
| Yes | 10 | 5 | 10 | 5 | pepper flowering stages | Yes | 3 | |
| 4 | No | 30 | 0 | 0 | 0 | pepper seedlings stages | No | 3 |
| No | 30 | 0 | 0 | 0 | pepper flowering stages | No | 3 | |
| No | 0 | 0 | 30 | 0 | pepper seedlings stages | No | 3 | |
| No | 0 | 0 | 30 | 0 | pepper flowering stages | No | 3 | |
| Yes | 15 | 0 | 15 | 0 | pepper seedlings stages | No | 3 | |
| Yes | 15 | 0 | 15 | 0 | pepper flowering stages | No | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, C.-H.; Qiao, F.-J.; Lu, Z.; Li, C.-Y.; Liu, T.-X.; Gao, Y.-L.; Zhang, B. Interspecific Competitions between Frankliniella intonsa and Frankliniella occidentalis on Fresh Lentil Bean Pods and Pepper Plants. Insects 2023, 14, 1. https://doi.org/10.3390/insects14010001
Yang C-H, Qiao F-J, Lu Z, Li C-Y, Liu T-X, Gao Y-L, Zhang B. Interspecific Competitions between Frankliniella intonsa and Frankliniella occidentalis on Fresh Lentil Bean Pods and Pepper Plants. Insects. 2023; 14(1):1. https://doi.org/10.3390/insects14010001
Chicago/Turabian StyleYang, Chun-Hong, Feng-Jiao Qiao, Zhaozhi Lu, Chang-You Li, Tong-Xian Liu, Yu-Lin Gao, and Bin Zhang. 2023. "Interspecific Competitions between Frankliniella intonsa and Frankliniella occidentalis on Fresh Lentil Bean Pods and Pepper Plants" Insects 14, no. 1: 1. https://doi.org/10.3390/insects14010001