
Functional Role of AsAP in the Reproduction of Adelphocoris suturalis (Hemiptera: Miridae)

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. cDNA Cloning and Sequence Analysis
2.3. Reverse Transcription–Quantitative Polymerase Chain Reaction (RT-qPCR)
2.4. RNA Interference (RNAi) in A. suturalis
2.5. Data Analysis
3. Results
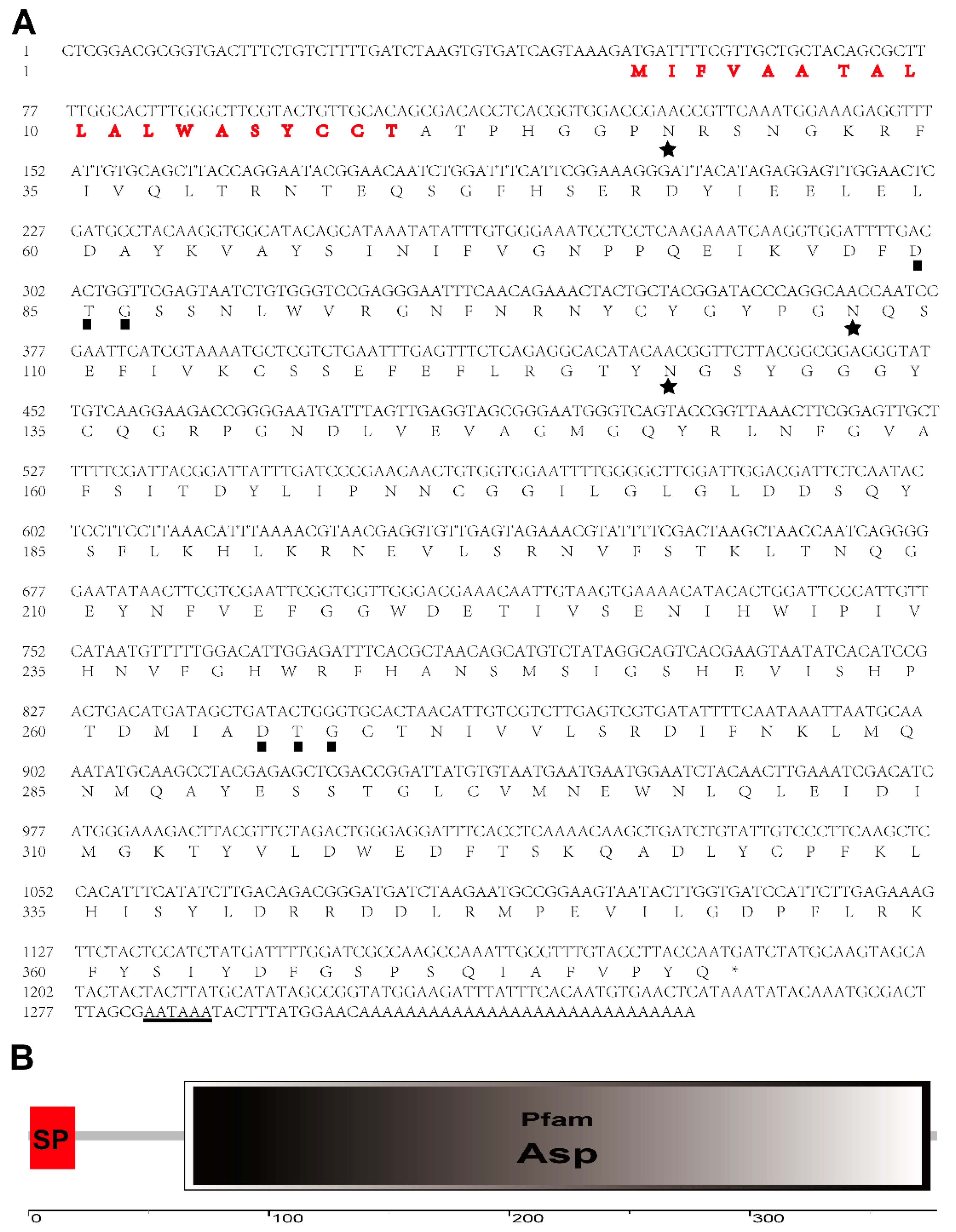
3.1. Cloning and Identification of A. suturalis Aspartic Protease (AsAP)
3.2. AsAP Is Highly Expressed in the Fat Body and Ovaries of Sexually Mature Females
3.3. AsAP Is Required for Female Fertility in A. suturalis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huis, A.V. The Global Impact of Insects; Wageningen UR: Wageningen, The Netherlands, 2014. [Google Scholar]
- Boldbaatar, D.; Sikalizyo Sikasunge, C.; Battsetseg, B.; Xuan, X.; Fujisaki, K. Molecular cloning and functional characterization of an aspartic protease from the hard tick Haemaphysalis longicornis. Insect Biochem. Mol. Biol. 2006, 36, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, N.D.; Barrett, A.J.; Finn, R. Twenty years of the MEROPS database of proteolytic enzymes, their substrates and inhibitors. Nucleic Acids Res. 2016, 44, D343–D350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Otín, C.; Bond, J.S. Proteases: Multifunctional enzymes in life and disease. J. Biol. Chem. 2008, 283, 30433–30437. [Google Scholar] [CrossRef] [Green Version]
- Sojka, D.; Franta, Z.; Horn, M.; Caffrey, C.R.; Mareš, M.; Kopáček, P. New insights into the machinery of blood digestion by ticks. Trends Parasitol. 2013, 29, 276–285. [Google Scholar] [CrossRef] [PubMed]
- Lehane, M.J. The Biology of Blood-Sucking in Insects, 3rd ed.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2005. [Google Scholar]
- Balczun, C.; Siemanowski, J.; Pausch, J.K.; Helling, S.; Marcus, K.; Stephan, C.; Meyer, H.E.; Schneider, T.; Cizmowski, C.; Oldenburg, M.; et al. Intestinal aspartate proteases TiCatD and TiCatD2 of the haematophagous bug Triatoma infestans (Reduviidae): Sequence characterisation, expression pattern and characterisation of proteolytic activity. Insect Biochem. Mol. Biol. 2012, 42, 240–250. [Google Scholar] [CrossRef]
- Sojka, D.; Hartmann, D.; Bartošová-Sojková, P.; Dvořák, J. Parasite Cathepsin D-Like Peptidases and Their Relevance as Therapeutic Targets. Trends Parasitol. 2016, 32, 708–723. [Google Scholar] [CrossRef]
- Cruz, C.E.; Fogaça, A.C.; Nakayasu, E.S.; Angeli, C.B.; Belmonte, R.; Almeida, I.C.; Miranda, A.; Miranda, M.T.M.; Tanaka, A.S.; Braz, G.R.; et al. Characterization of proteinases from the midgut of Rhipicephalus (Boophilus) microplus involved in the generation of antimicrobial peptides. Parasites Vectors 2010, 3, 63. [Google Scholar] [CrossRef] [Green Version]
- Logullo, C.; Da Silva Vaz, I.; Sorgine, M.H.F.; Paiva-Silva, G.O.; Faria, F.S.; Zingali, R.B.; De Lima, M.F.R.; Abreu, L.; Oliveira, E.F.; Alves, E.W.; et al. Isolation of an aspartic proteinase precursor from the egg of a hard tick, Boophilus microplus. Parasitology 1998, 116, 525–532. [Google Scholar] [CrossRef]
- Fire, A.; Xu, S.; Montgomery, M.K.; Kostas, S.A.; Driver, S.E.; Mello, C.C. Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature 1998, 391, 806–811. [Google Scholar] [CrossRef]
- Nandety, R.S.; Kuo, Y.W.; Nouri, S.; Falk, B.W. Emerging strategies for RNA interference (RNAi) applications in insects. Bioengineered 2015, 6, 8–19. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Nunes, M.A.; España, M.U.; Namin, H.H.; Jin, P.; Bensoussan, N.; Zhurov, V.; Rahman, T.; De Clercq, R.; Hilson, P.; et al. RNAi-based reverse genetics in the chelicerate model Tetranychus urticae: A comparative analysis of five methods for gene silencing. PLoS ONE 2017, 12, e0180654. [Google Scholar] [CrossRef] [PubMed]
- Ben-Amar, A.; Daldoul, S.; Reustle, G.M.; Krczal, G.; Mliki, A. Reverse genetics and high throughput sequencing methodologies for plant functional genomics. Curr. Genom. 2016, 17, 460–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San Miguel, K.; Scott, J.G. The next generation of insecticides: dsRNA is stable as a foliar-applied insecticide. Pest Manag. Sci. 2016, 72, 801–809. [Google Scholar] [CrossRef]
- Zhang, J.; Khan, S.A.; Heckel, D.G.; Bock, R. Next-Generation Insect-Resistant Plants: RNAi-Mediated Crop Protection. Trends Biotechnol. 2017, 35, 871–882. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Lu, Y.H.; Zeng, J. Forecast and Management of Mirid Bugs in Multiplie Agroecosystems of China; China Agriculture Press: Beijing, China, 2015. [Google Scholar]
- Zhen, C.; Miao, L.; Liang, P.; Gao, X. Survey of organophosphate resistance and an Ala216Ser substitution of acetylcholinesterase-1 gene associated with chlorpyrifos resistance in Apolygus lucorum (Meyer-Dur) collected from the transgenic Bt cotton fields in China. Pestic. Biochem. Physiol. 2016, 132, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Santiago, P.B.; de Araújo, C.N.; Motta, F.N.; Praça, Y.R.; Charneau, S.; Bastos, I.M.D.; Santana, J.M. Proteases of haematophagous arthropod vectors are involved in blood-feeding, yolk formation and immunity—A review. Parasites Vectors 2017, 10, 79. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Cheng, Y.; Guo, L.; Wang, A.; Lu, M.; Xu, L. Variation of gut microbiota caused by an imbalance diet is detrimental to bugs’ survival. Sci. Total Environ. 2021, 771, 144880. [Google Scholar] [CrossRef]
- Luo, J.; Liu, X.Y.; Liu, L.; Zhang, P.Y.; Chen, L.J.; Gao, Q.; Ma, W.H.; Chen, L.H.; Lei, C.L. De novo analysis of the Adelphocoris suturalis Jakovlev metathoracic scent glands transcriptome and expression patterns of pheromone biosynthesis-related genes. Gene 2014, 551, 271–278. [Google Scholar] [CrossRef]
- Liu, W.; Li, Y.; Zhu, L.; Zhu, F.; Lei, C.L.; Wang, X.P. Juvenile hormone facilitates the antagonism between adult reproduction and diapause through the methoprene-tolerant gene in the female Colaphellus bowringi. Insect Biochem. Mol. Biol. 2016, 74, 50–60. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Lindskog, I.; Gasteiger, E.; Bairoch, A.; Sanchez, J.C.; Hochstrasser, D.F.; Appel, R.D. Detailed peptide characterization using PEPTIDEMASS–a World-Wide-Web-accessible tool. Electrophoresis 1997, 18, 403–408. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Ma, C.; Li, Z.; Zhu, B.Q.; Zhang, J.; Lei, C.L.; Jin, S.X.; Hull, J.J.; Chen, L.Z. Assessment of suitable reference genes for qRT-PCR analysis in Adelphocoris suturalis. J. Integr. Agric. 2018, 17, 2745–2757. [Google Scholar] [CrossRef]
- Ma, M.; Tu, C.; Luo, J.; Lu, M.; Zhang, S.; Xu, L. Metabolic and immunological effects of gut microbiota in leaf beetles at the local and systemic levels. Integr. Zool. 2021, 16, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Luo, J.; Liang, S.; Li, J.; Xu, Z.; Li, L.; Zhu, B.; Li, Z.; Lei, C.; Lindsey, K.; Chen, L.; et al. A transgenic strategy for controlling plant bugs (Adelphocoris suturalis) through expression of double-stranded RNA homologous to fatty acyl-coenzyme A reductase in cotton. New Phytol. 2017, 215, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Raikhel, A.S.; Dhadialla, T.S. Accumulation of Yolk Proteins in Insect Oocytes. Annu. Rev. Entomol. 1992, 37, 217–251. [Google Scholar] [CrossRef]
- Cho, W.L.; Raikhel, A.S. Cloning of cDNA for mosquito lysosomal aspartic protease. Sequence analysis of an insect lysosomal enzyme similar to cathepsins D and E. J. Biol. Chem. 1992, 267, 21823–21829. [Google Scholar] [CrossRef]
- Swevers, L.; Raikhel, A.S.; Sappington, T.W.; Shirk, P.; Iatrou, K. Vitellogenesis and Post-Vitellogenic Maturation of the Insect Ovarian Follicle. In Comprehensive Molecular Insect Science; Gilbert, L.I., Iatrou, K., Gill, S.S., Eds.; Elsevier: Pergamon, Turkey, 2005; Volume 1, pp. 87–155. [Google Scholar]
- Cho, W.L.; Tsao, S.M.; Hays, A.R.; Walter, R.; Chen, J.S.; Snigirevskaya, E.S.; Raikhel, A.S. Mosquito Cathepsin B-like Protease Involved in Embryonic Degradation of Vitellin Is Produced as a Latent Extraovarian Precursor. J. Biol. Chem. 1999, 274, 13311–13321. [Google Scholar] [CrossRef] [Green Version]
- Tufail, M.; Takeda, M. Molecular characteristics of insect vitellogenins. J. Insect Physiol. 2008, 54, 1447–1458. [Google Scholar] [CrossRef]
- Hays, A.R.; Raikhel, A.S. A novel protein produced by the vitellogenic fat body and accumulated in mosquito oocytes. Rouxs. Arch. Dev. Biol. 1990, 199, 114–121. [Google Scholar] [CrossRef]
- Cho, W.L.; Deitsch, K.W.; Raikhel, A.S. An extraovarian protein accumulated in mosquito oocytes is a carboxypeptidase activated in embryos. Proc. Natl. Acad. Sci. USA 1991, 88, 10821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamahama, Y.; Yamamoto, Y.; Watabe, S.; Takahashi, S.Y. Regulation of yolk protein degradation during insect embryogenesis. In Progress in Vitellogenesis, Reproductive Biology of Invertebrates; Raikhel, A.S., Ed.; Science Publishers: Valencia, CA, USA, 2005; Volume 12, Part B, pp. 315–354. [Google Scholar]
- Nussenzveig, R.H.; Oliveira, P.L.; Masuda, H. Identification of yolk platelet-associated hydrolases in the oocytes of Rhodnius prolixus. Arch. Insect Biochem. 1992, 21, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Pohl, P.C.; Sorgine, M.H.F.; Leal, A.T.; Logullo, C.; Oliveira, P.L.; da Silva Vaz, I.; Masuda, A. An extraovarian aspartic protease accumulated in tick oocytes with vitellin-degradation activity. Comp. Biochem. Physiol. 2008, 151, 392–399. [Google Scholar] [CrossRef]
- Abreu, L.A.; Valle, D.; Manso, P.P.A.; Façanha, A.R.; Pelajo-Machado, M.; Masuda, H.; Masuda, A.; Vaz, I., Jr.; Lenzi, H.; Oliveira, P.L. Proteolytic activity of Boophilus microplus Yolk pro-Cathepsin D (BYC) is coincident with cortical acidification during embryogenesis. Insect Biochem. Mol. Biol. 2004, 34, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Leyria, J.; Fruttero, L.L.; Nazar, M.; Canavoso, L.E. The Role of DmCatD, a Cathepsin D-Like Peptidase, and Acid Phosphatase in the Process of Follicular Atresia in Dipetalogaster maxima (Hemiptera: Reduviidae), a Vector of Chagas’ Disease. PLoS ONE 2015, 10, e0130144. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F.; Nordin, J.H. Biochemical and ultrastructural aspects of vitellin utilization during embryogenesis. In Progress in Vitellogenesis, Reproductive Biology of Invertebrates; Part B; Raikhel, A.S., Ed.; Science Publishers: Valencia, CA, USA, 2005; Volume 12, pp. 355–392. [Google Scholar]
- Yamamoto, Y.; Takahashi, S.Y. Cysteine proteinase from Bombyx eggs: Role in programmed degradation of yolk proteins during embryogenesis. Comp. Biochem. Physiol. 1993, 106, 35–45. [Google Scholar] [CrossRef]
- Fagotto, F. Regulation of yolk degradation, or how to make sleepy lysosomes. J. Cell Sci. 1995, 108, 3645–3647. [Google Scholar] [CrossRef]
- Bell, W.J.; Bohm, M.K. Oosorption In Insects. Biol. Rev. 1975, 50, 373–396. [Google Scholar] [CrossRef]
- Moore, P.J.; Attisano, A. Oosorption in response to poor food: Complexity in the trade-off between reproduction and survival. Ecol. Evol. 2011, 1, 37–45. [Google Scholar] [CrossRef]
- Kotaki, T. Oosorption in the stink bug, Plautia crossota stali: Induction and vitellogenin dynamics. J. Insect Physiol. 2003, 49, 105–113. [Google Scholar] [CrossRef]
- Uchida, K.; Ohmori, D.; Ueno, T.; Nishizuka, M.; Eshita, Y.; Fukunaga, A.; Kominami, E. Preoviposition Activation of Cathepsin-like Proteinases in Degenerating Ovarian Follicles of the Mosquito Culex pipiens pallens. Dev. Biol. 2001, 237, 68–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medeiros, M.N.; Ramos, I.B.; Oliveira, D.M.P.; da Silva, R.C.B.; Gomes, F.M.; Medeiros, L.N.; Kurtenbach, E.; Chiarini, L.B.; Masuda, H.; de Souza, W.; et al. Microscopic and molecular characterization of ovarian follicle atresia in Rhodnius prolixus Stahl under immune challenge. J. Insect Physiol. 2011, 57, 945–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, A.N.; Boocock, M.R. Ovarian development in mosquitoes: Stages of growth and arrest, and follicular resorption. Physiol. Entomol. 1984, 9, 1–8. [Google Scholar] [CrossRef]
- Isoe, J.; Rascón, A.A., Jr.; Kunz, S.; Miesfeld, R.L. Molecular genetic analysis of midgut serine proteases in Aedes aegypti mosquitoes. Insect Biochem. Mol. Biol. 2009, 39, 903–912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro, J.M.C.; Genta, F.A.; Sorgine, M.H.F.; Logullo, R.; Mesquita, R.D.; Paiva-Silva, G.O.; Majerowicz, D.; Medeiros, M.; Koerich, L.; Terra, W.R.; et al. An Insight into the Transcriptome of the Digestive Tract of the Bloodsucking Bug, Rhodnius prolixus. PLoS Negl. Trop. Dis. 2014, 8, e2594. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Xu, X.; Gao, Y.; Yang, Q.; Zhu, Y.; Wang, J.; Wan, F.; Zhou, H. Levels of Salivary Enzymes of Apolygus lucorum (Hemiptera: Miridae), From 1st Instar Nymph to Adult, and Their Potential Relation to Bug Feeding. PLoS ONE 2016, 11, e0168848. [Google Scholar] [CrossRef]
- Lehane, M.J.; Billingsley, P. Biology of the Insect Midgut; Springer Science & Business: Berlin, Germany, 2012. [Google Scholar]
- Agustí, N.; Cohen, A.C. Lygus hesperus and L. lineolaris (Hemiptera: Miridae), Phytophages, Zoophages, or Omnivores: Evidence of Feeding Adaptations Suggested by the Salivary and Midgut Digestive Enzymes. J. Entomol. Sci. 2000, 35, 176–186. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.B.; Cai, W.J.; Wang, J.W.; Hong, G.J.; Tao, X.Y.; Wang, L.J.; Huang, Y.P.; Chen, X.Y. Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol. Nat. Biotechnol. 2007, 25, 1307–1313. [Google Scholar] [CrossRef]
- Kumar, P.; Pandit, S.S.; Steppuhn, A.; Baldwin, I.T. Natural history-driven, plant-mediated RNAi-based study reveals CYP6B46’s role in a nicotine-mediated antipredator herbivore defense. Proc. Natl. Acad. Sci. USA 2014, 111, 1245–1252. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.D.; Liu, Z.C.; Huang, S.L.; Chen, Z.Q.; Sun, Y.W.; Duan, P.F.; Ma, Y.Z.; Xia, L.Q. RNAi-mediated plant protection against aphids. Pest Manag. Sci. 2016, 72, 1090–1098. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Khan, S.A.; Hasse, C.; Ruf, S.; Heckel, D.G.; Bock, R. Full crop protection from an insect pest by expression of long double-stranded RNAs in plastids. Science 2015, 347, 991–994. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′–3′) |
|---|---|
| 5′ RACE | |
| B517-1 (GSP1) | tttcttgaggaggatt |
| B517-2 (GSP2) | atgctgtatgccaccttg |
| B517-3 (GSP3) | gttccaactcctctatgtaa |
| 3′ RACE | |
| C374-1 (Outer) | cacatttcatatcttgacagacggga |
| C374-2 (Inner) | tgaaatcgacatcatgggaaagact |
| ORF Cloning | |
| AsAP-F | tgatctaagtgtgatcagtaaagatgattt |
| AsAP-R | gttccataaagtatttaattcgctaaagtcg |
| RT-qPCR | |
| AsAP-F | acccaggcaaccaatccgaa |
| AsAP-R | actgacccattcccgctacc |
| EF1γ-F | ttggcccttgctgcagaacc |
| EF1γ-R | tctccgagccagatggagtagtt |
| dsRNA Synthesis | |
| dsAsAP-F | gcgtaatacgactcactataggcccgaacaactgtggtggaa |
| dsAsAP-R | gcgtaatacgactcactataggttggcttggcgatccaaaatc |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, S.; Zhu, B.; Huang, X.; Hull, J.J.; Chen, L.; Luo, J. Functional Role of AsAP in the Reproduction of Adelphocoris suturalis (Hemiptera: Miridae). Insects 2022, 13, 755. https://doi.org/10.3390/insects13080755
Qin S, Zhu B, Huang X, Hull JJ, Chen L, Luo J. Functional Role of AsAP in the Reproduction of Adelphocoris suturalis (Hemiptera: Miridae). Insects. 2022; 13(8):755. https://doi.org/10.3390/insects13080755
Chicago/Turabian StyleQin, Shidong, Bangqin Zhu, Xingxing Huang, J. Joe Hull, Lizhen Chen, and Jing Luo. 2022. "Functional Role of AsAP in the Reproduction of Adelphocoris suturalis (Hemiptera: Miridae)" Insects 13, no. 8: 755. https://doi.org/10.3390/insects13080755