
The Impact of Constrictotermes cyphergaster (Termitidae: Nasutitermitinae) Termites on Semiarid Ecosystems in Brazil: A Review of Current Research

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
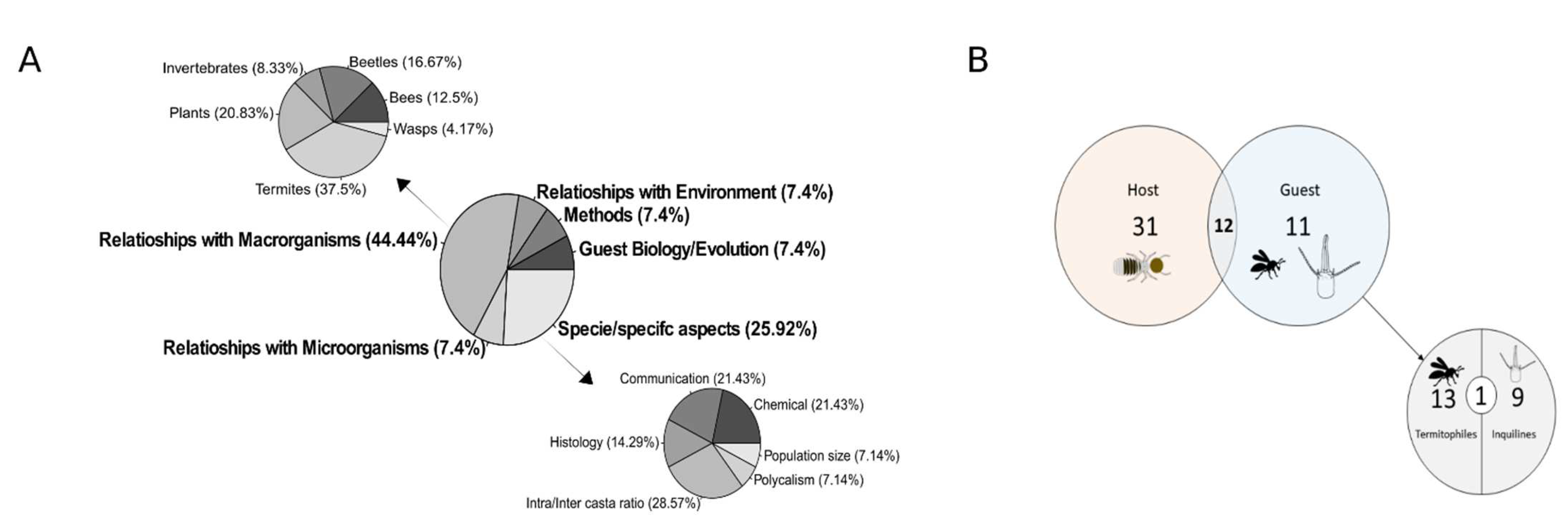
- Relationships with the environment: all papers directed towards the study of the modifications that termites made on the environment with their nests, and when the external environment influenced termites in their building location or behavior;
- Methods: studies aimed only at generating results for methods of maintenance of C. cyphergaster in bioassays or nest volume determination methods;
- Guest Biology/Evolution: Studies that focused on guest biology, or the evolution, phylogeny, or anatomy of guests of C. cyphergaster;
- Species/Specific aspects: Studies of specific characteristics of C. cyphergaster anatomy, morphology, the histology of individuals, or population size;
- Relationships with Microorganisms: Studies providing information on the relationships of C. cyphergaster with bacteria, lichens (mycobionts+ photobionts), and fungi;
- Relationships with Macroorganisms: works that focused on interactions with invertebrates or plants.
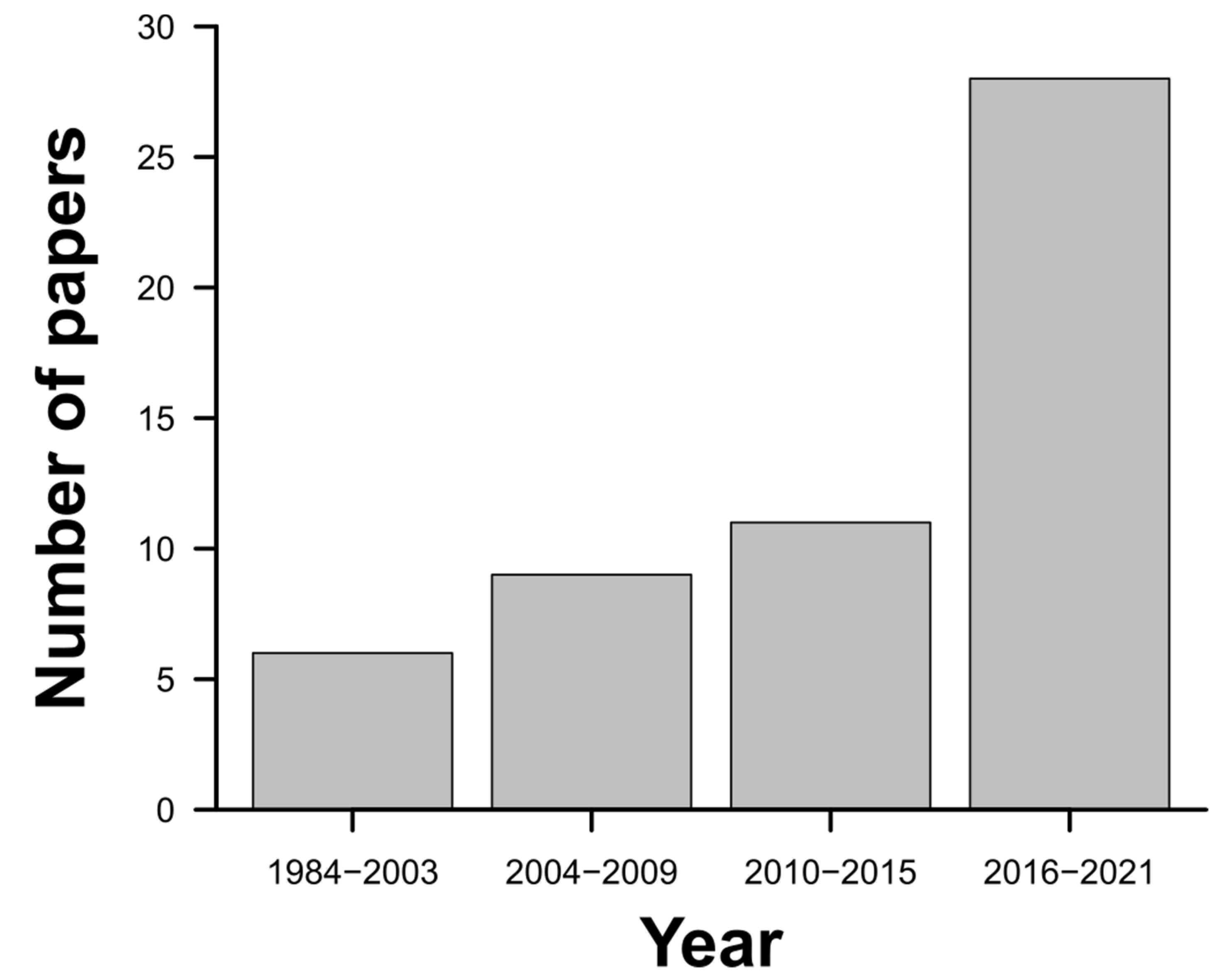
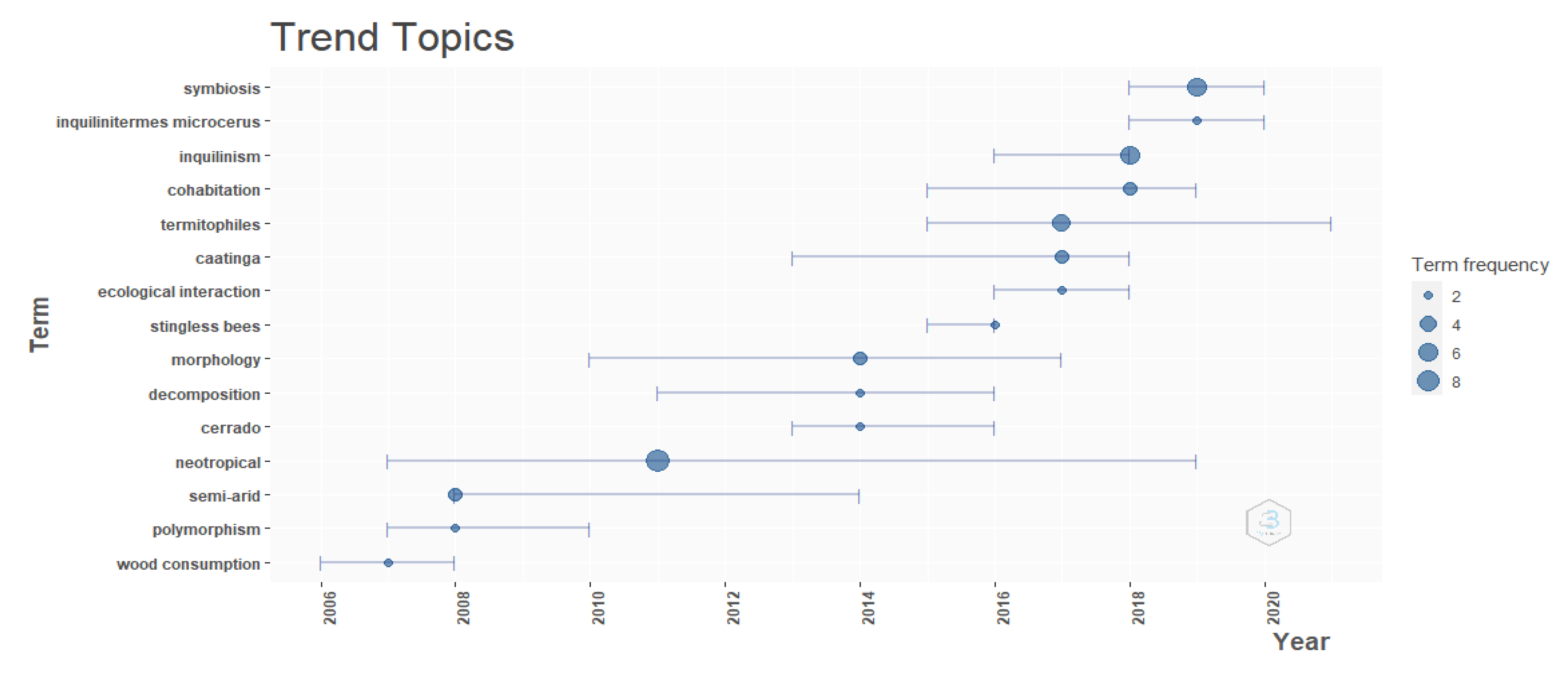
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clement, R.A.; Flores-Moreno, H.; Cernusak, L.A.; Cheesman, A.W.; Yatsko, A.R.; Allison, S.D.; Eggleton, P.; Zanne, A.E. Assessing the Australian Termite Diversity Anomaly: How Habitat and Rainfall Affect Termite Assemblages. Front. Ecol. Evol. 2021, 9, 1–13. [Google Scholar] [CrossRef]
- Abe, S.S.; Watanabe, Y.; Onishi, T.; Kotegawa, T.; Wakatsuki, T. Nutrient storage in termite (Macrotermes bellicosus) mounds and the implications for nutrient dynamics in a tropical savanna Ultisol. Soil Sci. Plant. Nutr. 2011, 57, 786–795. [Google Scholar] [CrossRef] [Green Version]
- Rückamp, D.; Martius, C.; Bragança, M.A.L.; Amelung, W. Lignin patterns in soil and termite nests of the Brazilian Cerrado. Appl. Soil Ecol. 2011, 48, 45–52. [Google Scholar] [CrossRef]
- Rückamp, D.; Amelung, W.; Theisz, N.; Bandeira, A.G.; Martius, C. Phosphorus forms in Brazilian termite nests and soils: Relevance of feeding guild and ecosystems. Geoderma 2010, 155, 269–279. [Google Scholar] [CrossRef]
- Rückamp, D.; Martius, C.; Bornemann, L.; Kurzatkowski, D. Geoderma Soil genesis and heterogeneity of phosphorus forms and carbon below mounds inhabited by primary and secondary termites. Geoderma 2012, 170, 239–250. [Google Scholar] [CrossRef]
- Alves, W.F.; Mota, A.S.; de Lima, R.A.A.; Bellezoni, R.; Vasconcellos, A. Termites as Bioindicators of Habitat Quality in the Caatinga, Brazil: Is There Agreement Between Structural Habitat Variables and the Sampled Assemblages? Neotrop. Entomol. 2011, 40, 39–46. [Google Scholar] [CrossRef] [Green Version]
- Ashton, L.A.; Griffiths, H.M.; Parr, C.L.; Evans, T.A.; Didham, R.K.; Hasan, F.; Teh, Y.A.; Tin, H.S.; Vairappan, C.S.; Eggleton, P. Termites mitigate the effects of drought in tropical rainforest. Science 2019, 363, 174–177. [Google Scholar] [CrossRef] [Green Version]
- Jouquet, P.; Traoré, S.; Choosai, C.; Hartmann, C.; Bignell, D. Influence of termites on ecosystem functioning. Ecosystem services provided by termites. Eur. J. Soil Biol. 2011, 47, 215–222. [Google Scholar] [CrossRef]
- Wood, T.G.; Sands, W.A. The role of termites in ecosystems. In Production Ecology of Ants and Termites; Brian, M.V., Ed.; Cambridge University Press: London, UK, 1978; pp. 245–292. [Google Scholar]
- de Bruyn, L.A.L.; Conacher, A.J. The role of termites and ants in soil modification: A review. Aust. J. Soil Res. 1990, 28, 55–93. [Google Scholar] [CrossRef]
- Whitford, W.G.; Ludwig, J.A.; Noble, J.C. The importance of subterranean termites in semi-arid ecosystems in south-eastern Australia. J. Arid Environ. 1992, 22, 87–91. [Google Scholar] [CrossRef]
- Banda, K.; Delgado-Salinas, A.; Dexter, K.G.; Linares-Palomino, R.; Oliveira-Filho, A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodríguez, G.M.; et al. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 2016, 353, 1383–1387. [Google Scholar]
- Werneck, F.; Costa, G.; Colli, G.; Prado, D.; Sites, J., Jr. Revisiting the historical distribution of Seasonally Dry Tropical Forests: New insights based on palaeodistribution modelling and palynological evidence. Glob. Ecol. Biogeogr. 2011, 20, 272–288. [Google Scholar] [CrossRef]
- Constantino, R. Termite Database 2020. Available online: http://termitologia.net (accessed on 11 June 2022).
- Mathews, A.G. Studies on Termites from Mato Grosso State, Brazil; Academia Brasileira de Ciencias: Rio de Janeiro, Brasil, 1977. [Google Scholar]
- Vasconcellos, A.; Araújo, V.F.P.; Moura, F.M.S.; Bandeira, A.G. Biomass and population structure of Constrictotermes cyphergaster (Silvestri) (Isoptera: Termitidae) in the dry forest of caatinga, northeastern Brazil. Neotrop. Entomol. 2007, 36, 693–698. [Google Scholar] [CrossRef] [Green Version]
- Constantino, R. Catalog of the living termites of the New World (Insecta: Isoptera). Arq. Zool. 1998, 35, 135–231. [Google Scholar] [CrossRef] [Green Version]
- Bezerra-Gusmão, M.A.; Barbosa, J.R.C.; Barbosa, M.R.D.V.; Bandeira, A.G.; Sampaio, E.V.S.B. Are nests of Constrictotermes cyphergaster (Isoptera, Termitidae) important in the C cycle in the driest area of semiarid caatinga in northeast Brazil? Appl. Soil Ecol. 2011, 47, 1–5. [Google Scholar] [CrossRef]
- da Cunha, H.F.; Brandão, D. Invertebrates associated with the neotropical termite Constrictotermes cyphergaster (Isoptera: Termitidae, Nasutitermitinae). Sociobiology 2001, 37, 593–599. [Google Scholar] [CrossRef] [Green Version]
- Barreto, L.S.; de Castro, M.S. Ecology of nesting of bees of the genus Partamona in caatinga, Milagres, Bahia. Biota Neotrop. 2007, 7, 1. [Google Scholar]
- Barbosa-Silva, A.M.; Silva, A.C.; Pereira, E.C.G.; Buril, M.L.L.; Silva, N.H.; Cáceres, M.E.S.; Aptroot, A.; Bezerra-Gusmão, M.A. Richness of Lichens Consumed by Constrictotermes cyphergaster in the Semi-arid Region of Brazil. Sociobiology 2019, 66, 154–160. [Google Scholar] [CrossRef] [Green Version]
- Barbosa-Silva, A.M.; Farias, M.A.A.; de Mello, A.P.; de Souza, A.E.F.; Garcia, H.H.M.; Bezerra-Gusmão, M.A. Lignocellulosic fungi in nests and food content of Constrictotermes cyphergaster and Inquilinitermes fur (Isoptera, Termitidae) from the semiarid region of Brazil. Fungal Ecol. 2016, 20, 75–78. [Google Scholar] [CrossRef]
- Oliveira, M.H.; Viana-Junior, A.B.; Nascimento, C.C.; Bezerra-Gusmão, M.A. Worker Dimorphism in Nasute Termites Reflects Different Tasks during Food Collection. J. Insect Behav. 2021, 34, 96–105. [Google Scholar] [CrossRef]
- Moura, F.M.S.; Vasconcellos, A.; Silva, N.B.; Bandeira, A.G. Caste development systems of the Neotropical termite Constrictotermes cyphergaster (Isoptera, Termitidae). Insectes Soc. 2011, 58, 169–175. [Google Scholar] [CrossRef]
- Leite, G.L.D.; Veloso, R.V.S.; Zanuncio, J.C.; Alves, S.M.; Amorim, C.A.D.; Souza, O.F.F. Factors Affecting Constrictotermes cyphergaster (Isoptera: Termitidae) Nesting on Caryocar brasiliense Trees in the Brazilian Savanna. Sociobiology 2011, 57, 165–180. [Google Scholar]
- Policarpo, I.S.; Vasconcellos, A.; Chaves, T.P.; Raimundo, J.P.; Medeiros, A.C.D.; Coutinho, H.D.M.; Alves, R.R.N. Transference of bioactive compounds from support plants to the termites Constrictotermes cyphergaster (Isoptera). Sci. Total Environ. 2018, 639, 921–928. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Aria, M.; Cuccurullo, C. bibliometrix: An R-tool for comprehensive science mapping analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Cristaldo, P.F.; Rosa, C.S.; Florencio, D.F.; Marins, A.; DeSouza, O. Termitarium volume as a determinant of invasion by obligatory termitophiles and inquilines in the nests of Constrictotermes cyphergaster (Termitidae, Nasutitermitinae). Insectes Soc. 2012, 59, 541–548. [Google Scholar] [CrossRef]
- Vargo, E.L.; Husseneder, C. Genetic Structure of Termites Colinies and Populations. In Biology of Termites: A Modern Synthesis; Bignell, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 321–347. [Google Scholar]
- Carrijo, T.F.; Pontes-Nogueira, M.; Santos, R.G.; Morales, A.C.; Cancello, E.M.; Scheffrahn, R.H. New World Heterotermes (Isoptera, Rhinotermitidae): Molecular phylogeny, biogeography and description of a new species. Syst. Entomol. 2020, 45, 527–539. [Google Scholar] [CrossRef]
- Scicchitano, V.; Dedeine, F.; Mantovani, B.; Luchetti, A. Molecular systematics, biogeography, and colony fusion in the European dry-wood termites Kalotermes spp. (Blattodea, Termitoidae, Kalotermitidae). Bull. Entomol. Res. 2018, 108, 523–531. [Google Scholar] [CrossRef]
- Hamilton, W.D. Altruism and Related Phenomena. Annu. Rev. Ecol. Syst. 1972, 3, 193–232. [Google Scholar] [CrossRef]
- Rosa, C.S.; Cristaldo, P.F.; Florencio, D.F.; Marins, A.; Lima, E.R.; DeSouza, O. On the Chemical Disguise of a Physogastric Termitophilous Rove Beetle. Sociobiology 2018, 65, 38–47. [Google Scholar] [CrossRef]
- Cristaldo, P.F.; DeSouza, O.; Krasulová, J.; Jirošová, A.; Kutalová, K.; Lima, E.R.; Šobotník, J.; Sillam-Dussès, D. Mutual use of trail-following chemical cues by a termite host and its inquiline. PLoS ONE 2014, 9, e85315. [Google Scholar] [CrossRef]
- Hugo, H.; Cristaldo, P.F.; DeSouza, O. Nonaggressive behavior: A strategy employed by an obligate nest invader to avoid conflict with its host species. Ecol. Evol. 2020, 10, 8741–8754. [Google Scholar] [CrossRef] [PubMed]
- Meer, R.; Breed, M.D.; Espelie, K.E.; Winston, M.L. Pheromone Communication in Social Insects, 1st ed.; Meer, R., Breed, M.D., Espelie, K.E., Winston, M.L., Eds.; CRC Press: New York, NY, USA, 1998; ISBN 0813389763. [Google Scholar]
- Moura, F.M.S.; Vasconcellos, A.; Araújo, V.F.P.; Bandeira, A.G. Feeding habit of Constrictotermes cyphergaster (Isoptera, Termitidae) in an area of caatinga, Northeast Brazil. Sociobiology 2006, 48, 1–6. [Google Scholar]
- Sun, P.; Li, G.; Jian, J.; Liu, L.; Chen, J.; Yu, S.; Xu, H.; Lei, C.; Zhou, X.; Huang, Q. Transcriptomic and Functional Analyses of Phenotypic Plasticity in a Higher Termite, Macrotermes barneyi Light. Front. Genet. 2019, 10, 964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, M.H.; Silva-Vieira, R.V.; Moreira, I.E.; Pires-Silva, C.M.; Lima, H.V.G.; Andrade, M.R.L.; Bezerra-Gusmão, M.A. “The road to reproduction”: Foraging trails of Constrictotermes cyphergaster (Termitidae: Nasutitermitinae) as maternities for Staphylinidae beetles. Sociobiology 2018, 65, 531–533. [Google Scholar] [CrossRef] [Green Version]
- Moreira, I.E.; Pires-Silva, C.M.; Ribeiro, K.G.; Zilberman, B.; Bezerra-Gusmão, M.A. Run to the nest: A parody on the Iron Maiden song by Corotoca spp.(Coleoptera, Staphylinidae). Pap. Avulsos Zool. 2019, 59, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Barbosa-Silva, A.M.; Vasconcellos, A. Consumption rate of lichens by Constrictotermes cyphergaster (Isoptera): Effects of C, N, and P contents and ratios. Insects 2019, 10, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa-Silva, A.M.; Santos, L.A.; Cáceres, M.E.S.; Vasconcellos, A. Constrictotermes cyphergaster (Blattaria, termitidae) termite nests as substrates for lichen fixation in the semiarid region of Northeastern Brazil. Braz. J. Biol. 2020, 80, 685–687. [Google Scholar] [CrossRef] [Green Version]
- Moura, F.M.S.; Vasconcellos, A.; Araújo, V.F.P.; Bandeira, A.G. Seasonality in foraging behaviour of Constrictotermes cyphergaster (Termitidae, Nasutitermitinae) in the Caatinga of northeastern Brazil. Insectes Soc. 2006, 53, 472–479. [Google Scholar] [CrossRef]
- Kistner, D.H. The social insects’ bestiary. In Social Insects; Hermann, H.R., Ed.; Academic Press: Cambridge, MA, USA, 1982; Volume 3, pp. 1–244. [Google Scholar]
- DeSouza, O.; Araújo, A.P.A.; Florencio, D.F.; Rosa, C.S.; Marins, A.; Costa, D.A.; Rodrigues, V.B.; Cristaldo, P.F. Allometric scaling of patrolling rate and nest volume in Constrictotermes cyphergaster termites: Hints on the settlement of inquilines. PLoS ONE 2016, 11, e0147594. [Google Scholar] [CrossRef]
- Da Cunha, H.F.; Lima, J.S.; De Souza, L.F.; Dos Santos, L.G.A.; Nabout, J.C. No Morphometric Distinction between the Host Constrictotermes cyphergaster (Silvestri) (Isoptera: Termitidae, Nasutitermitinae) and its Obligatory Termitophile Corotoca melantho Schiødte (Coleoptera: Staphylinidae). Sociobiology 2015, 62, 65–69. [Google Scholar] [CrossRef] [Green Version]
- da Cunha, H.F.; Costa, D.A.; do Espirito Santo Filho, K.; Silva, L.O.; Brandão, D. Relationship between Constrictotermes cyphergaster and inquiline termites in the Cerrado (Isoptera: Termitidae). Sociobiology 2003, 42, 761–770. [Google Scholar]
- Zilberman, B. Phylogenetic analysis of the genus Corotoca, with description of a new genus and species from Brazil (Coleoptera, Staphylinidae, Aleocharinae). Insect Syst. Evol. 2020, 1, 1–45. [Google Scholar] [CrossRef]
- Schiødte, J.M.C. On some Staphylinidae, found in the nests of termites. Proc. Zool. Soc. Lond. 1853, 21, 101–103. [Google Scholar]
- Silvestri, F. Contribuzione alla conoscenza dei Termitidi e Termitofili dell’ America Meridionale. Redia 1903, 1, 1–234. [Google Scholar]
- Pulleman, M.; Creamer, R.; Hamer, U.; Helder, J.; Pelosi, C.; Pérès, G.; Rutgers, M. Soil biodiversity, biological indicators and soil ecosystem services-an overview of European approaches. Curr. Opin. Environ. Sustain. 2012, 4, 529–538. [Google Scholar] [CrossRef]
- Zak, D.R.; Holmes, W.E.; White, D.C.; Peacock, A.D.; Tilman, D. Plant diversity, soil microbial communities, and ecosystem function: Are there any links? Ecology 2003, 84, 2042–2050. [Google Scholar] [CrossRef] [Green Version]
- Sutherland, W.J.; Freckleton, R.P.; Godfray, H.C.J.; Beissinger, S.R.; Benton, T.; Cameron, D.D.; Carmel, Y.; Coomes, D.A.; Coulson, T.; Emmerson, M.C.; et al. Identification of 100 fundamental ecological questions. J. Ecol. 2013, 101, 58–67. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conduction | No. | % | Description |
|---|---|---|---|
| Laboratory | 25 | 46.30 | Caatinga (7), Cerrado (10), undescribed (8) |
| Field | 21 | 38.89 | Caatinga (14), Cerrado (6), undescribed (1) |
| Both | 8 | 14.81 | Caatinga (4), Cerrado (4), undescribed (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliveira, M.H.d.; Viana-Junior, A.B.; Rolim, M.d.S.L.; Eloi, I.; Andrade, M.R.d.L.; Souza, J.J.L.L.d.; Bezerra-Gusmão, M.A. The Impact of Constrictotermes cyphergaster (Termitidae: Nasutitermitinae) Termites on Semiarid Ecosystems in Brazil: A Review of Current Research. Insects 2022, 13, 704. https://doi.org/10.3390/insects13080704
Oliveira MHd, Viana-Junior AB, Rolim MdSL, Eloi I, Andrade MRdL, Souza JJLLd, Bezerra-Gusmão MA. The Impact of Constrictotermes cyphergaster (Termitidae: Nasutitermitinae) Termites on Semiarid Ecosystems in Brazil: A Review of Current Research. Insects. 2022; 13(8):704. https://doi.org/10.3390/insects13080704
Chicago/Turabian StyleOliveira, Mário Herculano de, Arleu Barbosa Viana-Junior, Maria do Socorro Lacerda Rolim, Igor Eloi, Marllon Rinaldo de Lima Andrade, José João Lelis Leal de Souza, and Maria Avany Bezerra-Gusmão. 2022. "The Impact of Constrictotermes cyphergaster (Termitidae: Nasutitermitinae) Termites on Semiarid Ecosystems in Brazil: A Review of Current Research" Insects 13, no. 8: 704. https://doi.org/10.3390/insects13080704