
Live Drosophila melanogaster Larvae Deter Oviposition by Drosophila suzukii

 , , , and
, , , and 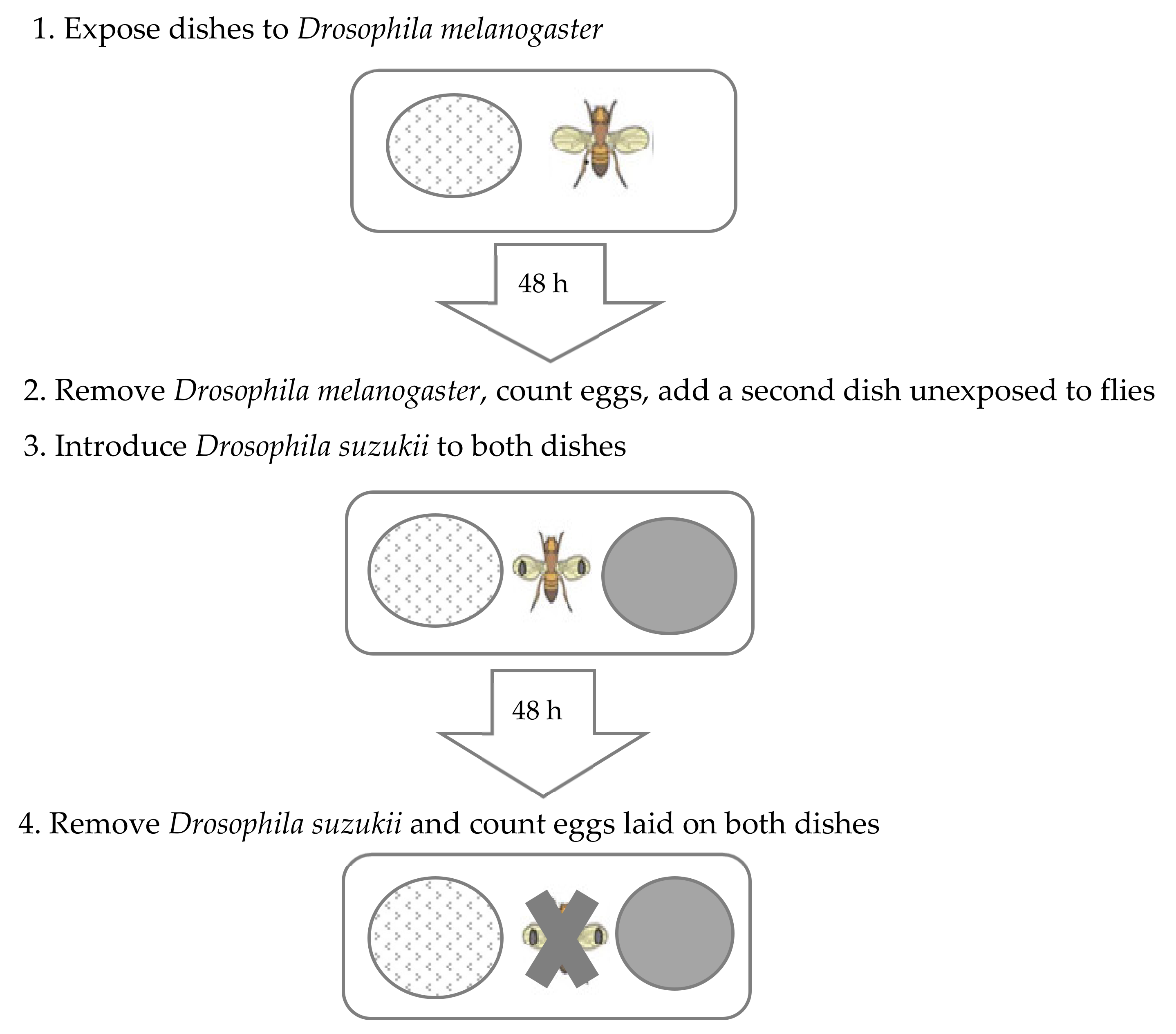{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila Cultures
2.2. General Procedure: Oviposition Choice Test
2.3. Experiments to Determine Life Stage of Drosophila melanogaster Responsible for Reduction in Oviposition by D. suzukii
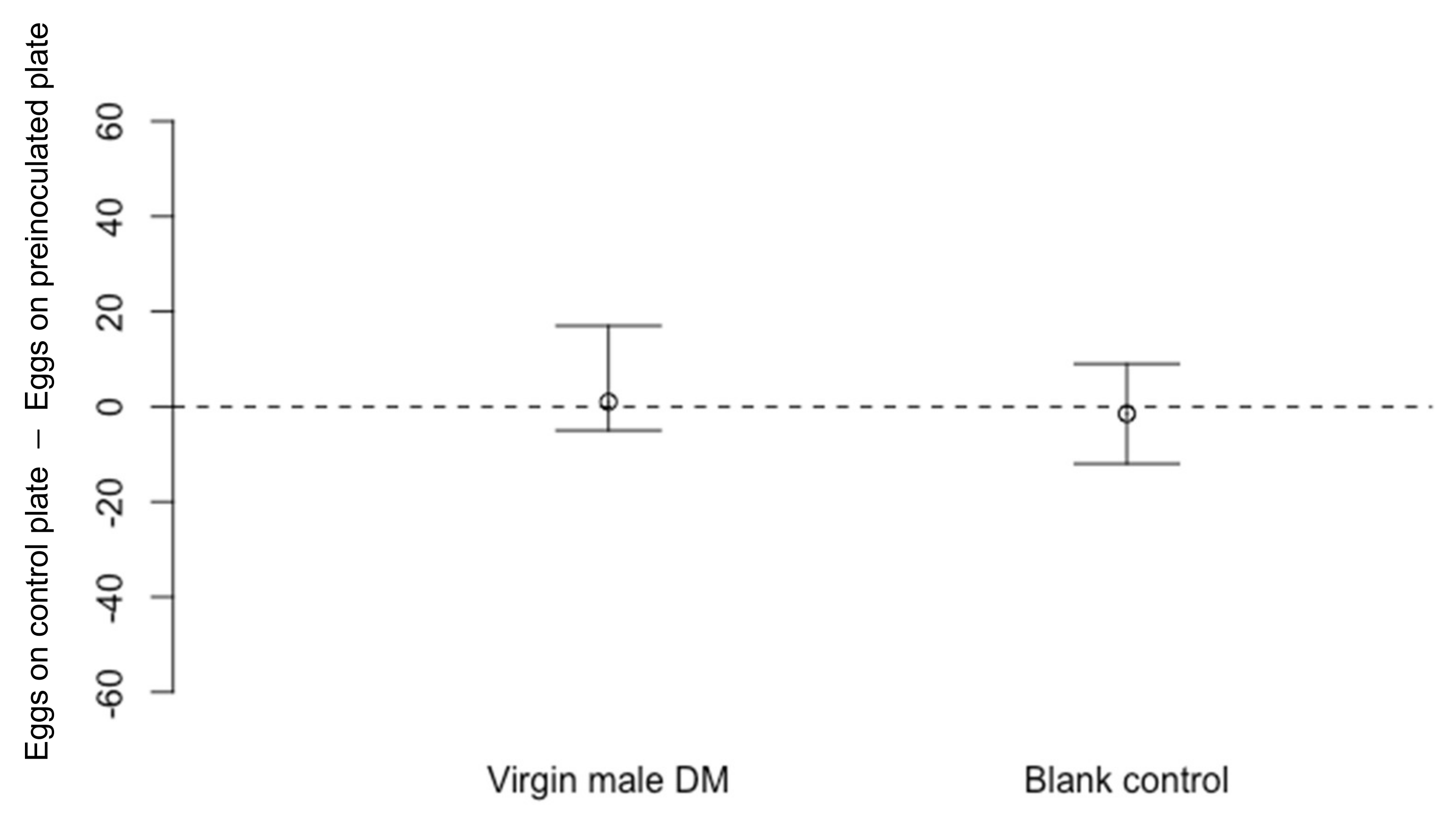
2.3.1. Effect of Adult Drosophila melanogaster on Oviposition by D. suzukii
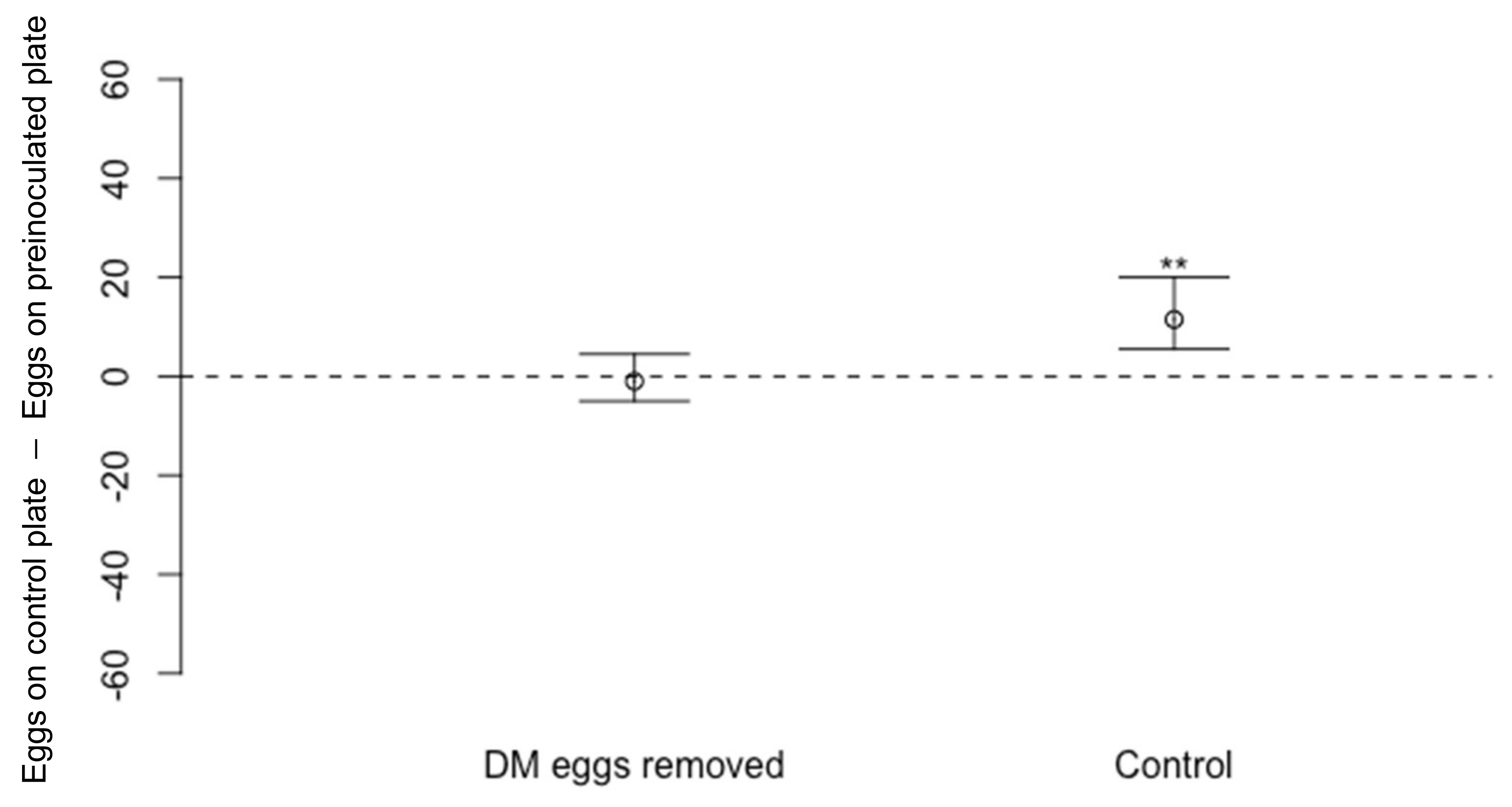
2.3.2. Effect of Drosophila melanogaster Eggs on Oviposition by D. suzukii
2.3.3. Effect of Drosophila melanogaster Larvae on Oviposition by D. suzukii
2.3.4. Effect of Drosophila melanogaster Larval Feeding Activity on Oviposition by D. suzukii
2.4. Extraction and Analyses of Cuticular Hydrocarbons (CHCs) from D. melanogaster and D. suzukii
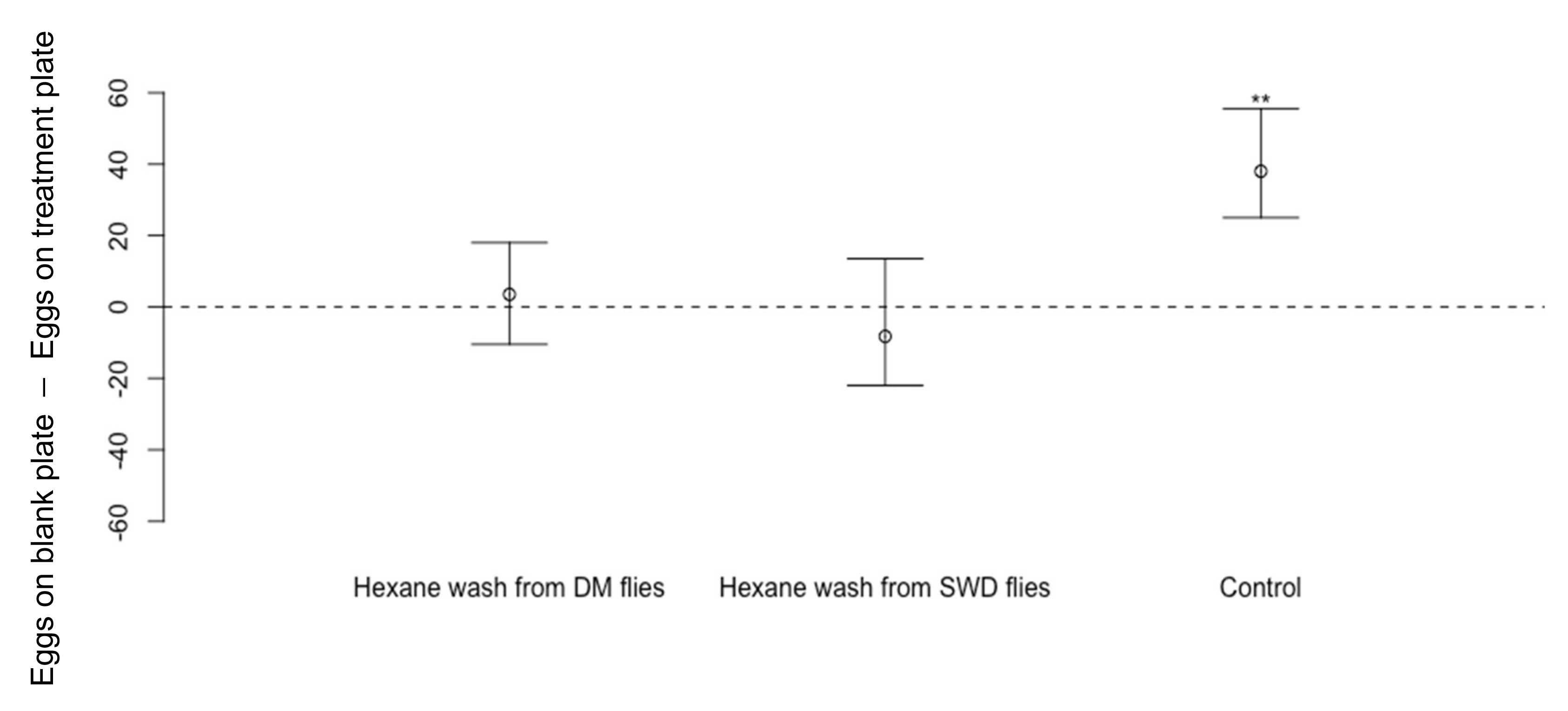
2.5. Effect of Cuticular Hydrocarbons (CHCs) from Adults or Larvae of D. melanogaster on Oviposition by D. suzukii
2.6. Statistical Analyses
3. Results
3.1. Experiments to Determine Life Stage of Drosophila melanogaster Responsible for Reduction in Oviposition by D. suzukii
3.1.1. Effect of Adult Drosophila melanogaster on Oviposition by D. suzukii
3.1.2. Effect of Drosophila melanogaster Eggs on Oviposition by D. suzukii
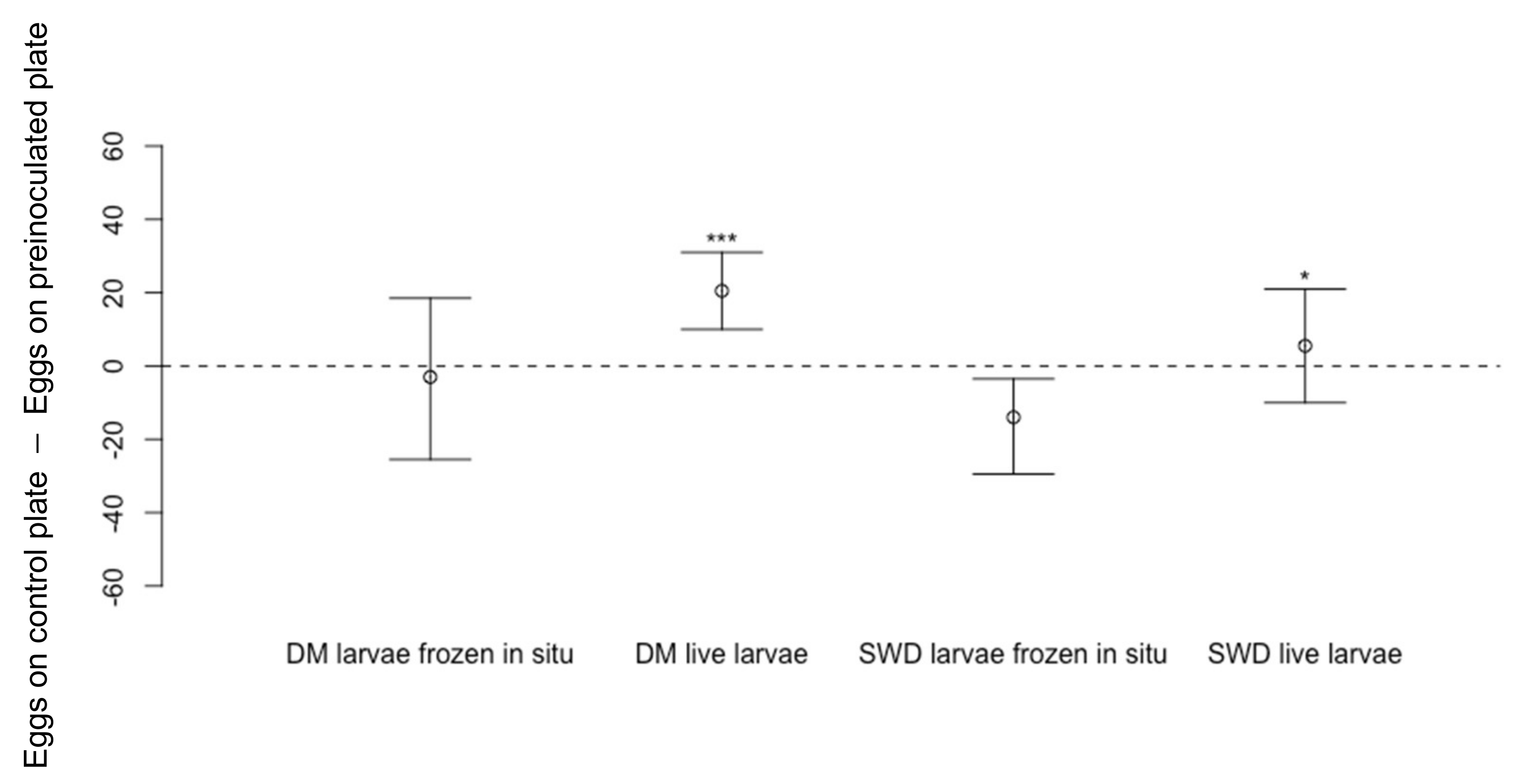
3.1.3. Effect of Drosophila melanogaster Larvae on Oviposition by D. suzukii
3.1.4. Effect of D. melanogaster Larval Feeding Activity on Oviposition by D. suzukii
3.2. Effect of Cuticular Hydrocarbons (CHCs) on Oviposition by Drosophila suzukii
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive Pest of Ripening Soft Fruit Expanding Its Geographic Range and Damage Potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Harris, A.L.; Shaw, B. First record of Drosophila suzukii Matsumura (Diptera: Drosophilidae) in Great Britain. Dipter. Dig. 2014, 21, 8. [Google Scholar]
- Andreazza, F.; Bernardi, D.; dos Santos, R.S.S.; Garcia, F.R.M.; Oliveira, E.E.; Botton, M.; Nava, D.E. Drosophila suzukii in Southern Neotropical Region: Current Status and Future Perspectives. Neotrop. Entomol. 2017, 46, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Hassani, I.M.; Behrman, E.L.; Prigent, S.R.; Gidaszewski, N.; Raveloson Ravaomanarivo, L.H.; Suwalski, A.; Debat, V.; David, J.R.; Yassin, A. First Occurrence of the Pest Drosophila suzukii (Diptera: Drosophilidae) in the Comoros Archipelago (Western Indian Ocean). Afr. Entomol. 2020, 28, 78–83. [Google Scholar] [CrossRef]
- Atallah, J.; Teixeira, L.; Salazar, R.; Zaragoza, G.; Kopp, A. The Making of a Pest: The Evolution of a Fruit-Penetrating Ovipositor in Drosophila suzukii and Related Species. Proc. R. Soc. B Biol. Sci. 2014, 281, 20132840. [Google Scholar] [CrossRef] [Green Version]
- Rombaut, A.; Guilhot, R.; Xuéreb, A.; Benoit, L.; Chapuis, M.P.; Gibert, P.; Fellous, S. Invasive Drosophila suzukii Facilitates Drosophila melanogaster Infestation and Sour Rot Outbreaks in the Vineyards. R. Soc. Open Sci. 2017, 4, 170117. [Google Scholar] [CrossRef] [Green Version]
- Ioriatti, C.; Guzzon, R.; Anfora, G.; Ghidoni, F.; Mazzoni, V.; Villegas, T.R.; Dalton, D.T.; Walton, V.M. Drosophila suzukii (Diptera: Drosophilidae) Contributes to the Development of Sour Rot in Grape. J. Econ. Entomol. 2018, 111, 283–292. [Google Scholar] [CrossRef]
- Emiljanowicz, L.M.; Ryan, G.D.; Langille, A.; Newman, J. Development, Reproductive Output and Population Growth of the Fruit Fly Pest Drosophila suzukii (Diptera: Drosophilidae) on Artificial Diet. J. Econ. Entomol. 2014, 107, 1392–1398. [Google Scholar] [CrossRef] [Green Version]
- Tochen, S.; Dalton, D.T.; Wiman, N.; Hamm, C.; Shearer, P.W.; Walton, V.M. Temperature-Related Development and Population Parameters for Drosophila suzukii (Diptera: Drosophilidae) on Cherry and Blueberry. Environ. Entomol. 2014, 43, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Little, C.M.; Chapman, T.W.; Hillier, N.K. Plasticity Is Key to Success of Drosophila suzukii (Diptera: Drosophilidae) Invasion. J. Insect Sci. 2020, 20, 5. [Google Scholar] [CrossRef]
- Van Timmeren, S.; Isaacs, R. Control of Spotted Wing Drosophila, Drosophila suzukii, by Specific Insecticides and by Conventional and Organic Crop Protection Programs. Crop Prot. 2013, 54, 126–133. [Google Scholar] [CrossRef]
- Haye, T.; Girod, P.; Cuthbertson, A.G.S.; Wang, X.G.; Daane, K.M.; Hoelmer, K.A.; Baroffio, C.; Zhang, J.P.; Desneux, N. Current SWD IPM Tactics and Their Practical Implementation in Fruit Crops across Different Regions around the World. J. Pest Sci. 2016, 89, 643–651. [Google Scholar] [CrossRef]
- Noble, R.; Dobrovin-Pennington, A.; Shaw, B.; Buss, D.S.; Cross, J.V.; Fountain, M.T. Fermentation for Disinfesting Fruit Waste from Drosophila Species (Diptera: Drosophilidae). Environ. Entomol. 2017, 46, 939–945. [Google Scholar] [CrossRef]
- Schöneberg, T.; Lewis, M.T.; Burrack, H.J.; Grieshop, M.; Isaacs, R.; Rendon, D.; Rogers, M.; Rothwell, N.; Sial, A.A.; Walton, V.M.; et al. Cultural Control of Drosophila suzukii in Small Fruit—Current and Pending Tactics in the U.S. Insects 2021, 12, 172. [Google Scholar] [CrossRef]
- Van Timmeren, S.; Fanning, P.D.; Schöneberg, T.; Hamby, K.; Lee, J.; Isaacs, R. Exploring the Efficacy and Mechanisms of a Crop Sterilant for Reducing Infestation by Spotted-Wing Drosophila (Diptera: Drosophilidae). J. Econ. Entomol. 2020, 113, 288–298. [Google Scholar] [CrossRef]
- Disi, J.O.; Van Timmeren, S.; Gress, B.; Zalom, F.; Isaacs, R.; Sial, A. Insecticide Residue Longevity for On-site Screening of Drosophila suzukii (Matsumura) Resistance. Pest Manag. Sci. 2020, 76, 2918–2924. [Google Scholar] [CrossRef]
- Gress, B.E.; Zalom, F.G. Identification and Risk Assessment of Spinosad Resistance in a California Population of Drosophila suzukii. Pest Manag. Sci. 2019, 75, 1270–1276. [Google Scholar] [CrossRef]
- Civolani, S.; Vaccari, G.; Caruso, S.; Finetti, L.; Bernacchia, G.; Chicca, M.; Cassanelli, S. Evaluation of Insecticide Efficacy and Insecticide Adaptive Response in Italian Populations of Drosophila suzukii. Bull. Insectol. 2021, 74, 103–114. [Google Scholar]
- Kienzle, R.; Groß, L.B.; Caughman, S.; Rohlfs, M. Resource Use by Individual Drosophila suzukii Reveals a Flexible Preference for Oviposition into Healthy Fruits. Sci. Rep. 2020, 10, 3132. [Google Scholar] [CrossRef] [Green Version]
- Little, C.M.; Chapman, T.W.; Moreau, D.L.; Hillier, N.K. Susceptibility of Selected Boreal Fruits and Berries to the Invasive Pest Drosophila suzukii (Diptera: Drosophilidae). Pest Manag. Sci. 2017, 73, 160–166. [Google Scholar] [CrossRef]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The Wide Potential Trophic Niche of the Asiatic Fruit Fly Drosophila suzukii: The Key of Its Invasion Success in Temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef] [Green Version]
- Shaw, B.; Brain, P.; Wijnen, H.; Fountain, M.T. Reducing Drosophila suzukii Emergence through Inter-Species Competition: Reducing D. suzukii Emergence through Competition. Pest Manag. Sci. 2018, 74, 1466–1471. [Google Scholar] [CrossRef] [Green Version]
- Kidera, H.; Takahashi, K.H. Chemical Cues from Competitors Change the Oviposition Preference of Drosophila suzukii. Entomol. Exp. Appl. 2020, 168, 304–310. [Google Scholar] [CrossRef]
- Dancau, T.; Stemberger, T.L.M.; Clarke, P.; Gillespie, D.R. Can Competition Be Superior to Parasitism for Biological Control? The Case of Spotted Wing Drosophila (Drosophila suzukii), Drosophila melanogaster and Pachycrepoideus Vindemmiae. Biocontrol Sci. Technol. 2017, 27, 3–16. [Google Scholar] [CrossRef]
- Gao, H.-H.; Zhai, Y.-F.; Chen, H.; Wang, Y.-M.; Liu, Q.; Hu, Q.-L.; Ren, F.-S.; Yu, Y. Ecological Niche Difference Associated with Varied Ethanol Tolerance between Drosophila suzukii and Drosophila melanogaster (Diptera: Drosophilidae). Fla. Entomol. 2018, 101, 498–504. [Google Scholar] [CrossRef]
- Yew, J.Y.; Dreisewerd, K.; Luftmann, H.; Müthing, J.; Pohlentz, G.; Kravitz, E.A. A New Male Sex Pheromone and Novel Cuticular Cues for Chemical Communication in Drosophila. Curr. Biol. 2009, 19, 1245–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, R.W.; Blomquist, G.J. Ecological, behavioral, and biochemical aspects of insect hydrocarbons. Ann. Rev. 2005, 50, 371–393. [Google Scholar] [CrossRef] [PubMed]
- Ferveur, J.-F. Cuticular Hydrocarbons: Their Evolution and Roles in Drosophila Pheromonal Communication. Behav. Genet. 2005, 35, 279–295. [Google Scholar] [CrossRef]
- Farine, J.-P.; Ferveur, J.-F.; Everaerts, C. Volatile Drosophila Cuticular Pheromones Are Affected by Social but Not Sexual Experience. PLoS ONE 2012, 7, e40396. [Google Scholar] [CrossRef]
- Dekker, T.; Revadi, S.; Mansourian, S.; Ramasamy, S.; Lebreton, S.; Becher, P.G.; Angeli, S.; Rota-Stabelli, O.; Anfora, G. Loss of Drosophila Pheromone Reverses Its Role in Sexual Communication in Drosophila suzukii. Proc. R. Soc. B Biol. Sci. 2015, 282, 20143018. [Google Scholar] [CrossRef] [Green Version]
- Snellings, Y.; Herrera, B.; Wildermann, B.; Beelen, M.; Zwarts, L.; Wenseleers, T.; Callaerts, P. The role of cuticular hydrocarbons in mate recognition in Drosophila suzukii. Sci. Rep. 2018, 8, 4996. [Google Scholar] [CrossRef]
- Beevor, P.S.; Hall, D.R.; Hendarsih, M.S. Identification and field evaluation of the female sex pheromone of the white stem borer, Scirpophaga innotata. In Proceedings of the 4th International Conference on Plant Protection in the Tropics, Kuala Lumpur, Malaysia, 28–31 March 1994; pp. 140–142, ISBN 967-9942-17-1. [Google Scholar]
- Billeter, J.-C.; Atallah, J.; Krupp, J.J.; Millar, J.G.; Levine, J.D. Specialized cells tag sexual and species identity in Drosophila melanogaster. Nat. Lett. 2009, 461, 987–991. [Google Scholar] [CrossRef]
- Lebreton, S.; Trona, S.; Borrero-Echeverry, F.; Bilz, F.; Grabe, V.; Becher, P.G.; Carlsson, M.A.; Nässel, D.R.; Hansson, B.S.; Sachse, S.; et al. Feeding regulates sex pheromone attraction and courtship in Drosophila females. Sci. Rep. 2015, 5, 13132. [Google Scholar] [CrossRef] [Green Version]
- Everaerts, C.; Farine, J.-P.; Cobb, M.; Ferveur, J.-F. Drosophila Cuticular Hydrocarbons Revisited: Mating Status Alters Cuticular Profiles. PLoS ONE 2010, 5, e9607. [Google Scholar] [CrossRef]
- Dweck, H.K.M.; Ebrahim, S.A.M.; Thoma, M.; Mohamed, A.A.M.; Keesey, I.W.; Trona, F.; Lavista-Llanos, S.; Svatoš, A.; Sachse, S.; Knaden, M.; et al. Pheromones Mediating Copulation and Attraction in Drosophila. Proc. Natl. Acad. Sci. USA 2015, 112, E2829–E2835. [Google Scholar] [CrossRef] [Green Version]
- Kienzle, R.; Rohlfs, M. Mind the Wound!—Fruit Injury Ranks Higher than, and Interacts with, Heterospecific Cues for Drosophila suzukii Oviposition. Insects 2021, 12, 424. [Google Scholar] [CrossRef]
- Henry, Y.; Tarapacki, P.; Colinet, H. Larval density affects phenotype and surrounding bacterial community without altering gut microbiota in Drosophila melanogaster. FEMS Microbiol. Ecol. 2020, 96, fiaa055. [Google Scholar] [CrossRef]
- Keesey, I.W.; Koerte, S.; Retzke, T.; Haverkamp, A.; Hansson, B.S.; Knaden, M. Adult Frass Provides a Pheromone Signature for Drosophila Feeding and Aggregation. J. Chem. Ecol. 2016, 42, 739–747. [Google Scholar] [CrossRef] [Green Version]
- Chandler, J.A.; Morgan Lang, J.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial Communities of Diverse Drosophila Species: Ecological Context of a Host–Microbe Model System. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef]
- Chandler, J.A.; Eisen, J.A.; Kopp, A. Yeast Communities of Diverse Drosophila Species: Comparison of Two Symbiont Groups in the Same Hosts. Appl. Environ. Microbiol. 2012, 78, 7327–7336. [Google Scholar] [CrossRef] [Green Version]
- Adair, K.L.; Bost, A.; Bueno, E.; Kaunisto, S.; Kortet, R.; Peters-Schulze, G.; Martinson, V.G.; Douglas, A.E. Host Determinants of Among-Species Variation in Microbiome Composition in Drosophilid Flies. ISME J. 2020, 14, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.; Fountain, M.T.; Günther, C.S.; Eady, P.E.; Goddard, M.R. Separate and Combined Hanseniaspora Uvarum and Metschnikowia Pulcherrima Metabolic Volatiles Are Attractive to Drosophila suzukii in the Laboratory and Field. Sci. Rep. 2021, 11, 1201. [Google Scholar] [CrossRef] [PubMed]
- Wong, A.C.-N.; Wang, Q.-P.; Morimoto, J.; Senior, A.M.; Lihoreau, M.; Neely, G.G.; Simpson, S.J.; Ponton, F. Gut Microbiota Modifies Olfactory-Guided Microbial Preferences and Foraging Decisions in Drosophila. Curr. Biol. 2017, 27, 2397–2404.e4. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Keesey, I.W.; Hansson, B.S.; Knaden, M. Gut Microbiota Affects Development and Olfactory Behavior in Drosophila melanogaster. J. Exp. Biol. 2019, 222, jeb.192500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, A.; Tanaka, K.M.; Yew, J.Y.; Takahashi, A. Drosophila suzukii Avoidance of Microbes in Oviposition Choice. R. Soc. Open Sci. 2021, 8, 201601. [Google Scholar] [CrossRef]
- Scheidler, N.H.; Liu, C.; Hamby, K.A.; Zalom, F.G.; Syed, Z. Volatile Codes: Correlation of Olfactory Signals and Reception in Drosophila-Yeast Chemical Communication. Sci. Rep. 2015, 5, 14059. [Google Scholar] [CrossRef] [Green Version]
- Mazzetto, F.; Gonella, E.; Crotti, E.; Vacchini, V.; Syrpas, M.; Pontini, M.; Mangelinckx, S.; Daffonchio, D.; Alma, A. Olfactory Attraction of Drosophila suzukii by Symbiotic Acetic Acid Bacteria. J. Pest Sci. 2016, 89, 783–792. [Google Scholar] [CrossRef]
- Bolton, L.G.; Piñero, J.C.; Barrett, B.A. Electrophysiological and Behavioral Responses of Drosophila suzukii (Diptera: Drosophilidae) Towards the Leaf Volatile β-Cyclocitral and Selected Fruit-Ripening Volatiles. Environ. Entomol. 2019, 48, 1049–1055. [Google Scholar] [CrossRef]
- Bolton, L.G.; Piñero, J.C.; Barrett, B.A. Olfactory Cues From Host- and Non-Host Plant Odor Influence the Behavioral Responses of Adult Drosophila suzukii (Diptera: Drosophilidae) to Visual Cues. Environ. Entomol. 2021, 50, 571–579. [Google Scholar] [CrossRef]
- Tait, G.; Park, K.; Nieri, R.; Crava, M.C.; Mermer, S.; Clappa, E.; Boyer, G.; Dalton, D.T.; Carlin, S.; Brewer, L.; et al. Reproductive Site Selection: Evidence of an Oviposition Cue in a Highly Adaptive Dipteran, Drosophila suzukii (Diptera: Drosophilidae). Environ. Entomol. 2020, 49, 355–363. [Google Scholar] [CrossRef]
- Elsensohn, J.E.; Aly, M.F.K.; Schal, C.; Burrack, H.J. Social Signals Mediate Oviposition Site Selection in Drosophila suzukii. Sci. Rep. 2021, 11, 3796. [Google Scholar] [CrossRef]
- Durisko, Z.; Kemp, R.; Mubasher, R.; Dukas, R. Dynamics of Social Behavior in Fruit Fly Larvae. PLoS ONE 2014, 9, e95495. [Google Scholar] [CrossRef]
- Durisko, Z.; Dukas, R. Attraction to and Learning from Social Cues in Fruitfly Larvae. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131398. [Google Scholar] [CrossRef] [Green Version]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tungadi, T.D.; Shaw, B.; Powell, G.; Hall, D.R.; Bray, D.P.; Harte, S.J.; Farman, D.I.; Wijnen, H.; Fountain, M.T. Live Drosophila melanogaster Larvae Deter Oviposition by Drosophila suzukii. Insects 2022, 13, 688. https://doi.org/10.3390/insects13080688
Tungadi TD, Shaw B, Powell G, Hall DR, Bray DP, Harte SJ, Farman DI, Wijnen H, Fountain MT. Live Drosophila melanogaster Larvae Deter Oviposition by Drosophila suzukii. Insects. 2022; 13(8):688. https://doi.org/10.3390/insects13080688
Chicago/Turabian StyleTungadi, Trisna D., Bethan Shaw, Glen Powell, David R. Hall, Daniel P. Bray, Steven J. Harte, Dudley I. Farman, Herman Wijnen, and Michelle T. Fountain. 2022. "Live Drosophila melanogaster Larvae Deter Oviposition by Drosophila suzukii" Insects 13, no. 8: 688. https://doi.org/10.3390/insects13080688