
Changes in Diversity and Structure of Thrips (Thysanoptera) Assemblages in the Spruce Forest Stands of High Tatra Mts. after a Windthrow Calamity

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling
2.3. Data Analysis
3. Results
3.1. Vertical and Horizontal Stratification of Thrips
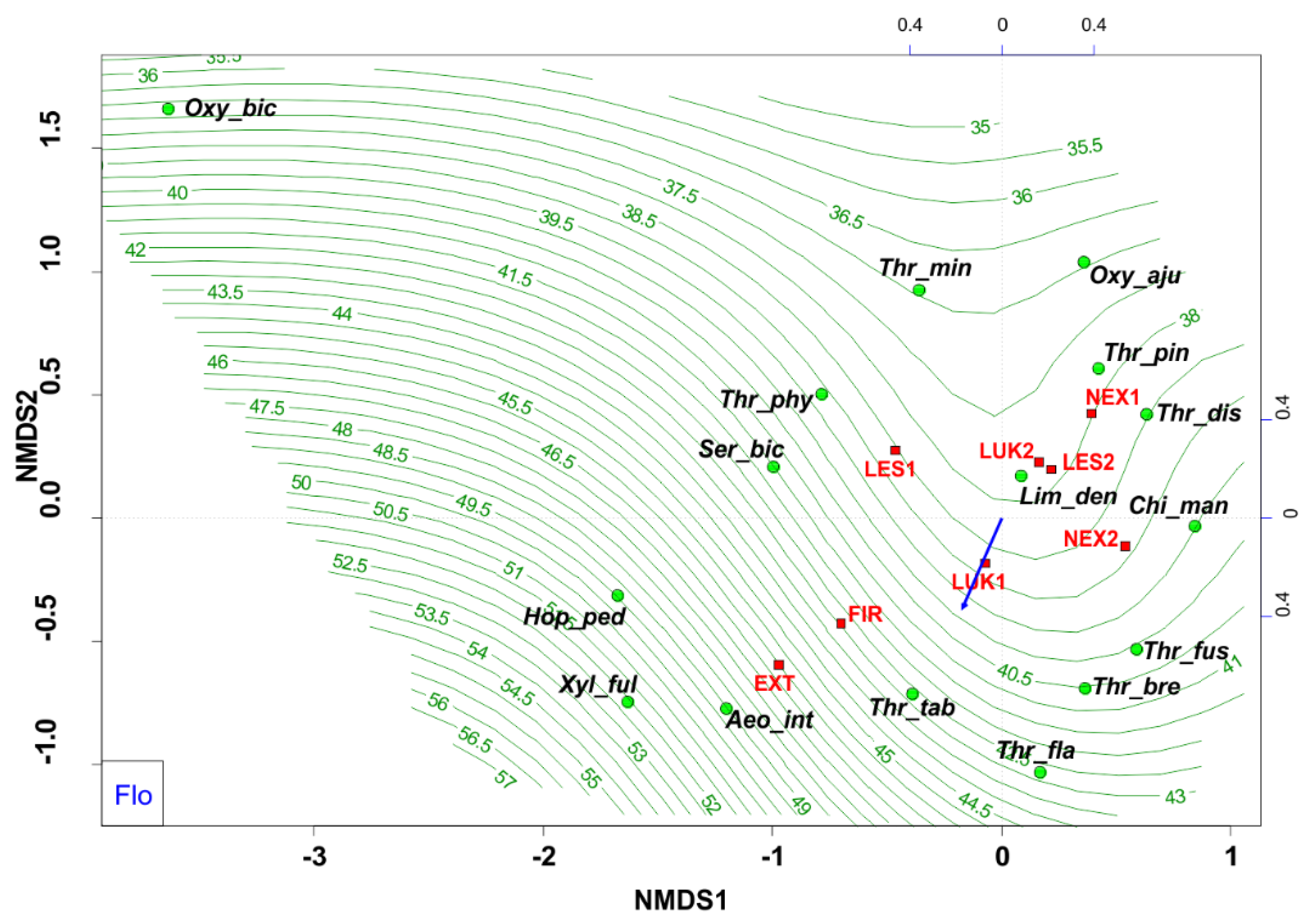
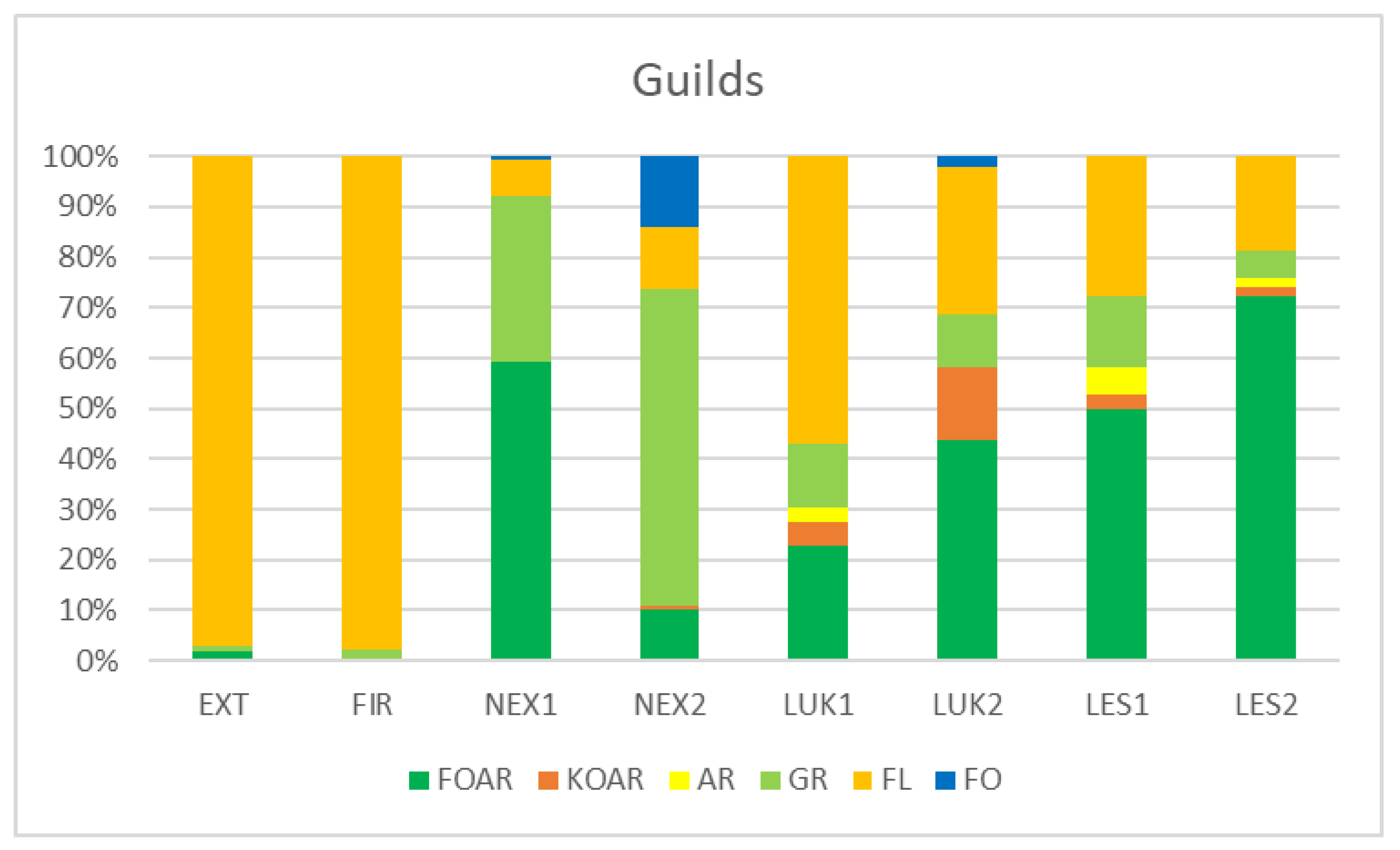
3.2. Functional Structure of Thrips Assemblages
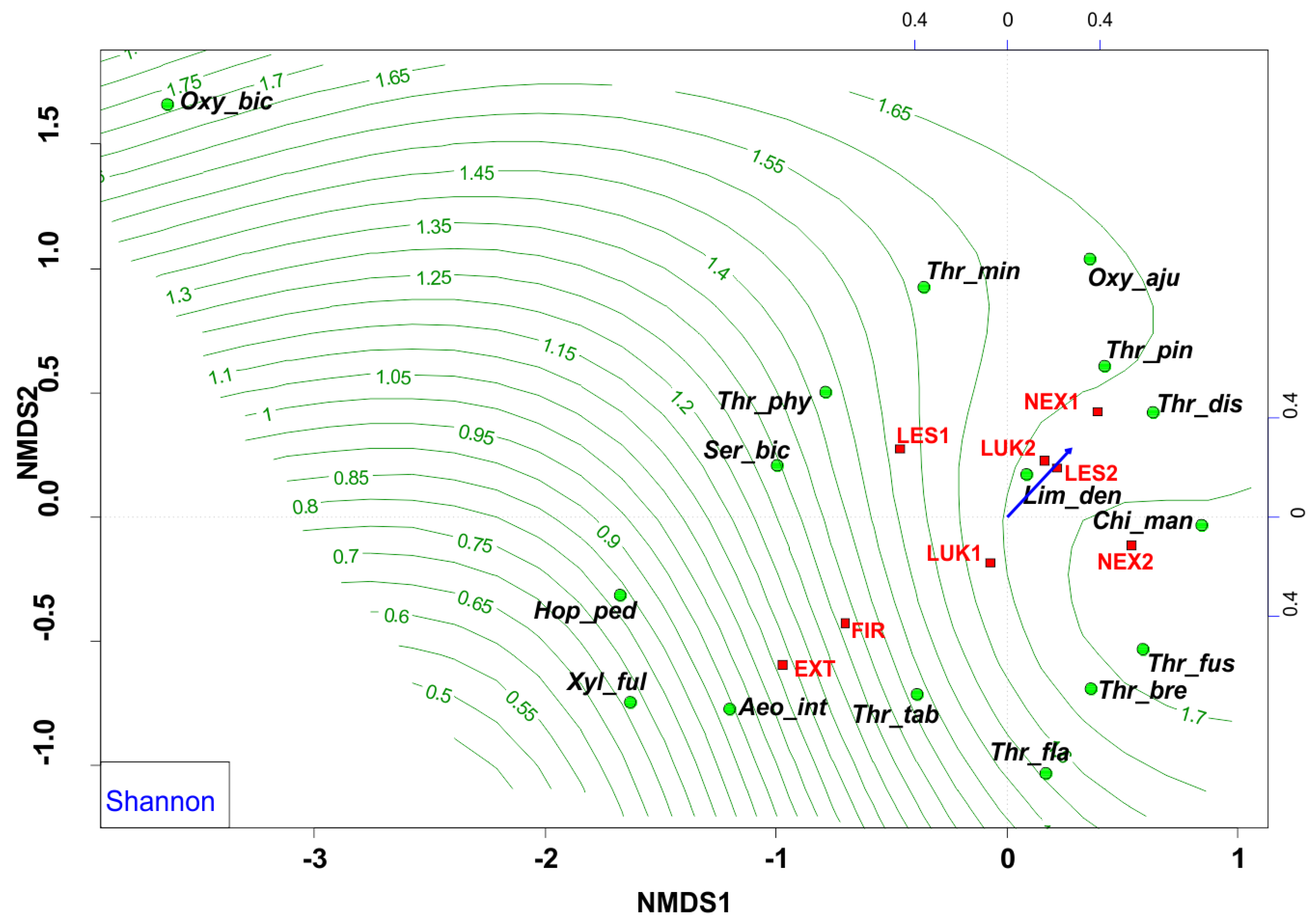
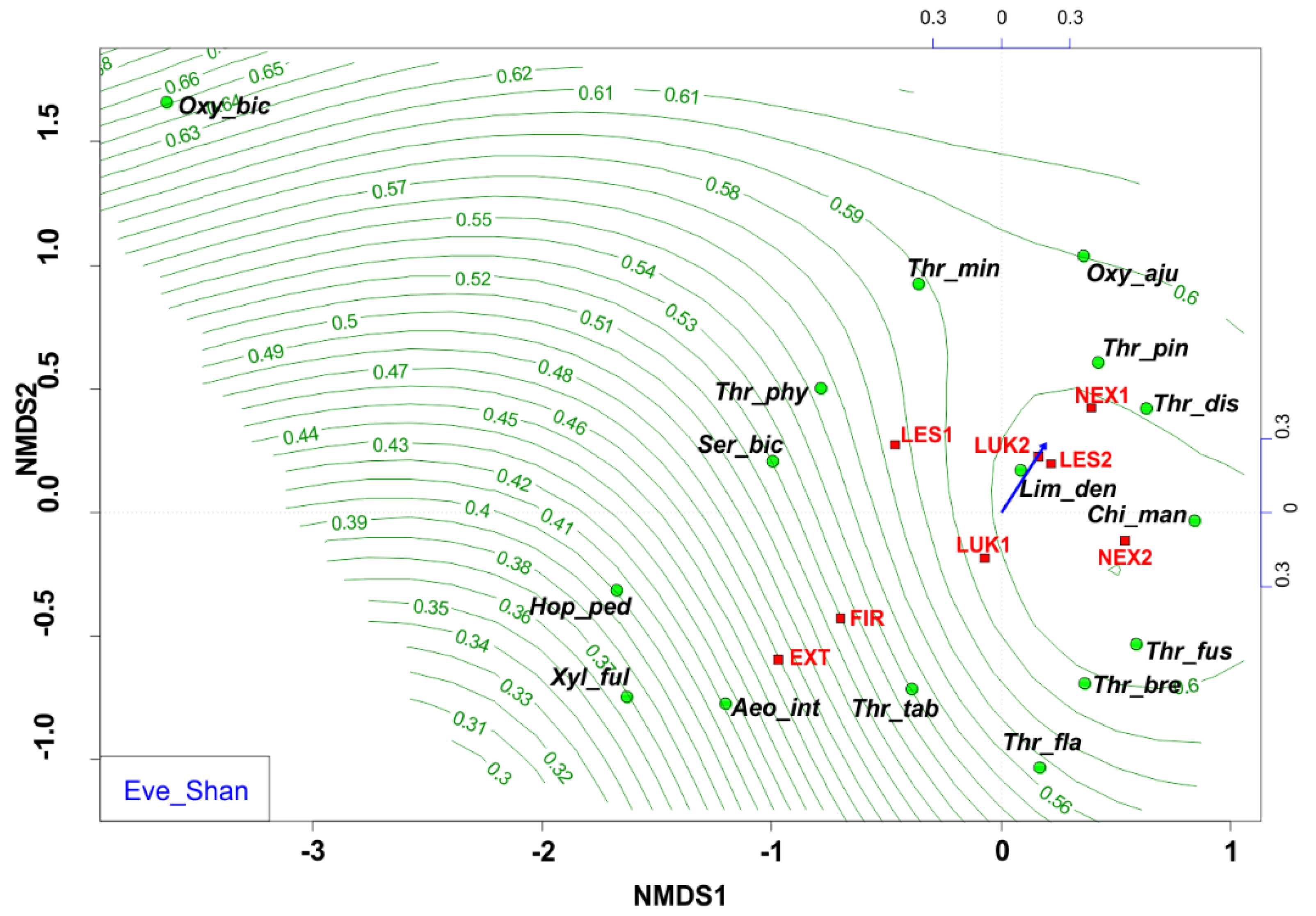
3.3. Species Richness and Diversity of Thrips Assemblages
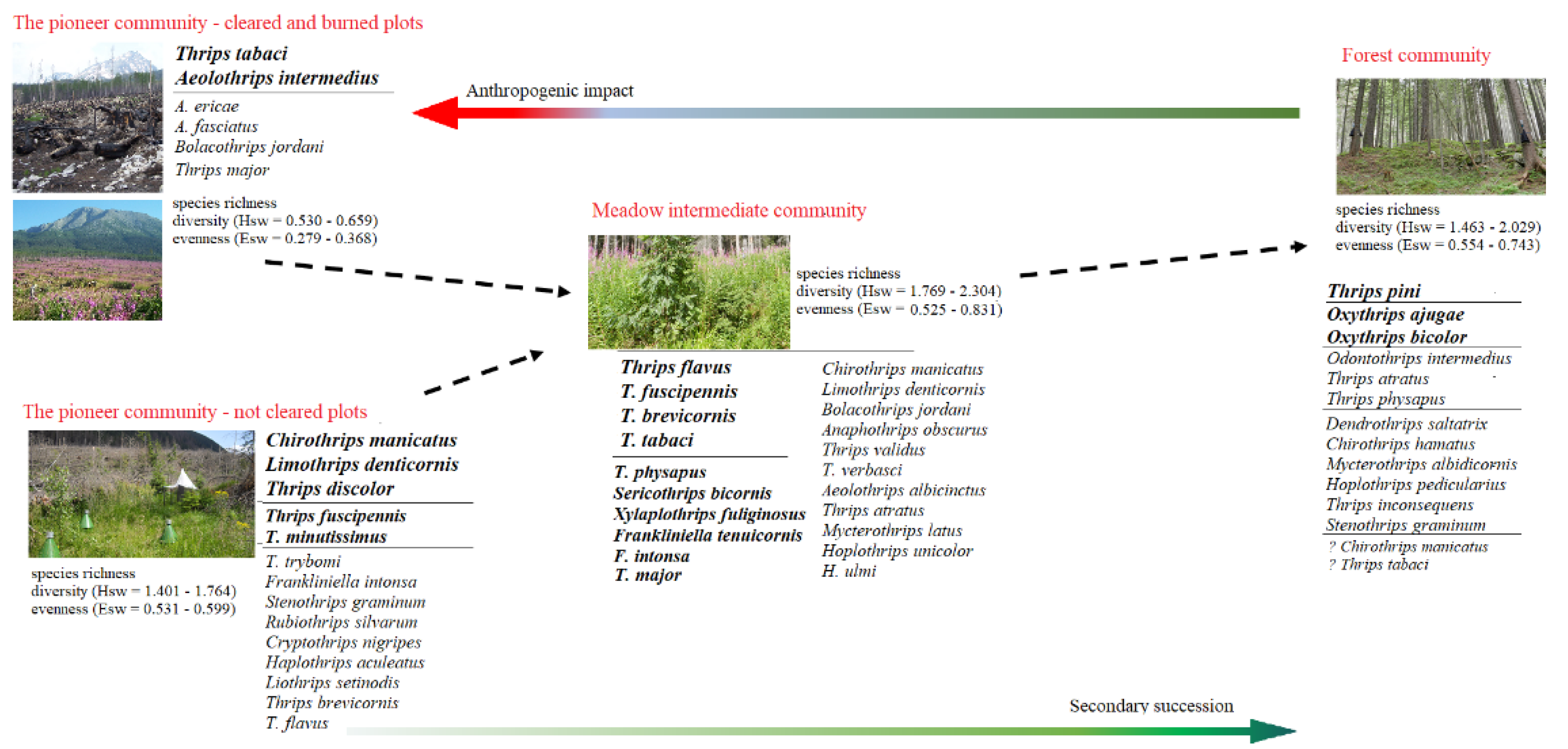
3.4. Anthropogenic Impact
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lässig, R.; Močalov, S.A. Frequency and characteristics of severe storms in the Urals and their influence on the development, structure and management of the boreal forests. For. Ecol. Manag. 2000, 135, 179–194. [Google Scholar] [CrossRef]
- Peltonen, M. Windthrows and dead standing trees as bark beetle breeding material at forest clearcut edge. Scand. J. For. Res. 1999, 14, 505–511. [Google Scholar] [CrossRef]
- Bouget, C.; Duelli, P. The effects of windthrow on forest insect communities: A literature review. Biol. Conserv. 2004, 118, 281–299. [Google Scholar] [CrossRef]
- Holeksa, J.; Jaloviar, P.; Kucbel, S.; Saniga, M.; Svoboda, M.; Szewcyk, J.; Szwagrzyk, J.; Zielonka, T.; Žywiec, M. Models of disturbance driven dynamics in the West Carpathian spruce forests. For. Ecol. Manag. 2017, 388, 79–89. [Google Scholar] [CrossRef]
- Janda, P.; Bače, R.; Trotsiuk, V.; Mikoláš, M.; Mrhalová, H.; Morrisey, R.C.; Matějů, L.; Lábusová, J.; Nagel, T.; Kucbel, S.; et al. The historical disturbance regime of mountain Norway spruce forests in the Western Carpathians and its influence on current forest structure and composition. For. Ecol. Manag. 2017, 388, 67–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wermelinger, B.; Moretti, M.; Duelli, P.; Lachat, T.; Pezzatti, G.B.; Obrist, M.K. Impact of windthrow and salvage-logging on taxonomic and functional diversity of forest arthropods. For. Ecol. Manag. 2017, 391, 9–18. [Google Scholar] [CrossRef]
- Faliński, J.B. Vegetation dynamics in temperate lowland primeval forest. Ecological studies in Bialowieza forest. Geobotany 1986, 8, 1±537. [Google Scholar]
- Schelhaas, M.J.; Nabuurs, G.J.; Schuck, A. Natural disturbances in the European forests in the 19th and 20th centuries. Glob. Change Biol. 2003, 9, 1620–1633. [Google Scholar] [CrossRef]
- Zackrisson, O. Influence of forest fires on the North Swedish boreal forest. Oikos 1977, 29, 22–32. [Google Scholar] [CrossRef]
- Perlin, J. A Forest Journey: The Role of Wood in the Development of Civilization; Norton: New York, NY, USA, 1989. [Google Scholar]
- Gossner, M.M.; Floren, A.; Weisser, W.; Linsenmair, K.E. Effect of dead wood enrichment in the canopy and on the forest floor on beetle guild composition. For. Ecol. Manag. 2013, 302, 404–413. [Google Scholar] [CrossRef]
- Leidinger, J.; Seibold, S.; Weisser, W.W.; Lange, M.; Schall, P.; Turke, M.; Gossner, M.M. Effects of forest management on herbivorous insects in temperate Europe. For. Ecol. Manag. 2019, 437, 232–245. [Google Scholar] [CrossRef]
- Johansson, T.; Hjältén, J.; Gibb, H.; Hilszczanski, J.; Stenlidb, J.; Balla, J.P.; Alinvi, O.; Danell, K. Variable response of different functional groups of saproxylic beetles to substrate manipulation and forest management: Implications for conservation strategies. For. Ecol. Manag. 2007, 242, 496–510. [Google Scholar] [CrossRef]
- Overbeck, M.; Schmidt, M. Modelling infestation risk of Norway spruce by Ips typographus (L.) in the Lower Saxon Harz Mountains (Germany). For. Ecol. Manag. 2012, 266, 115–125. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Lexer, M.J. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Change Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Seidl, R.; Thom, D.; Kautz, M.; Martin-Benito, D.; Peltoniemi, M.; Vacchiano, G.; Wild, J.; Ascoli, D.; Petr, M.; Honkaniemi, J.; et al. Forest disturbances under climate change. Nat. Clim. Change 2017, 7, 395–402. [Google Scholar] [CrossRef] [Green Version]
- Davis, F.W.; Moritz, M. Mechanisms of disturbance. In Encyclopedia of Biodiversity; Levin, S.A., Ed.; Academic Press: San Diego, CA, USA, 2001; pp. 153–160. [Google Scholar]
- Koreň, M. Vetrová kalamita 19.11.2004, nové pohľady a konsekvencie. Tatry 2005, 44, 6–29. [Google Scholar]
- Fleisher, P. Windfall research and monitoring in the High Tatra Mts., objectives, principles, methods, and current status. Contrib. Geophys. Geod. 2008, 38, 233–248. [Google Scholar]
- Fleisher, P. Pokalamitný výskum vo Vysokých Tatrách–ciele, metódy a charakteristika lokalít. Štúdie O Tatranskom Národnom Parku 2011, 10, 7–12. [Google Scholar]
- Hanajík, P.; Fritze, H. Effects of forest management on soil properties at windthrow area in Tatra National Park. Acta Environ. Univ. Comen. 2009, 17, 36–46. [Google Scholar]
- Budzáková, M.; Galvánek, D.; Littera, P.; Šibík, J. The wind and fire disturbance in Central European mountain spruce forests: The regeneration after four years. Acta Soc. Bot. Pol. 2013, 82, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Marini, L.; Lindelöw, Å.; Jönsson, A.M.; Wulff, S.; Schroeder, L.M. Population dynamics of the spruce bark beetle: A long-term study. Oikos 2013, 122, 1768–1776. [Google Scholar] [CrossRef]
- Wermelinger, B.; Obrist, M.K.; Baur, H.; Jakoby, O.; Duelli, P. Synchronous rise and fall of bark beetle and parasitoid populations in windthrow areas. Agric. For. Entomol. 2013, 15, 301–309. [Google Scholar] [CrossRef]
- Duelli, P.; Obrist, M.K.; Wermelinger, B. Windthrow-induced changes in faunistic biodiversity in alpine spruce forests. For. Snow Landsc. Res. 2002, 77, 117–131. [Google Scholar]
- Bellmann, H. Arthropoden der Kraut-und Strauchschicht [Herb and shrub arthropoda]. In The Development of Biotic Communities after Windthrow; Fischer, A., Ed.; Ecomed: Landsberg, Germany, 1998; pp. 280–281. [Google Scholar]
- Hilszczański, J.; Gibb, H.; Hjältén, J.; Atlegrim, O.; Johansson, T.; Pettersson, R.B.; Ball, J.P.; Danell, K. Parasitoids (Hymenoptera, Ichneumonoidea) of Saproxylic beetles are a Vected by forest successional stage and dead wood characteristics in boreal spruce forest. Biol. Conserv. 2005, 126, 456–464. [Google Scholar] [CrossRef]
- Lewis, T. Thrips. Their Biology, Ecology, and Economic Importance; Academic Press: London, UK, 1973. [Google Scholar]
- Pelikán, J. Kpoznání třásněnek Jeseníku II. Thysanoptera. Přírodovědecký Sborník Ostrav. Kraj. 1950, 11, 345–354. [Google Scholar]
- Oettingen, H. Beiträge zur Thysanopterenfauna Schwedens. Entomol. Tidskr. 1954, 75, 134–150. [Google Scholar]
- Pelikán, J.; Fedor, P.; Krumpál, M.; Cyprich, D. Thrips (Thysanoptera) in nests of birds and mammals in Slovakia. Ekológia 2002, 21, 275–282. [Google Scholar]
- Vasiliu-Oromulu, L. Niche and overlap in thrips populations. In Proceedings of the Third International Symposium on Thysanoptera, Kazimierz Dolny, Poland, 11–16 June 1990; pp. 57–63. [Google Scholar]
- Masarovič, R.; Doričová, M.; Prokop, P.; Fedor, P. “Testing the limits”—An interesting record of the exotic banded greenhouse thrips Hercinothrips femoralis (Thysanoptera: Thripidae: Panchaetothripinae) at high Carpathian mountain altitudes. Biologia 2014, 69, 1631–1634. [Google Scholar] [CrossRef]
- Dubovský, M.; Fedor, P.; Kucharczyk, H.; Masarovič, R.; Balkovič, J. Assemblages of bark−dwelling thrips (Thysanoptera) of uneven−aged oak forests in Slovakia. Sylwan 2010, 154, 659–668. [Google Scholar]
- Zvaríková, M.; Kiseľák, J.; Zvarík, M.; Masarovič, R.; Prokop, P.; Fedor, P. Ecological gradients and environmental impact in the forest dwelling Haplothrips subtilissimus (Thysanoptera: Phlaeothripidae) phenotypic variability. Ecol. Indic. 2016, 66, 313–320. [Google Scholar] [CrossRef]
- Vasiliu-Oromulu, L. Ecological investigations on Thysanoptera in the Garbova-Massif Romania. Folia Entomol. Hung. 1989, 50, 157–163. [Google Scholar]
- Vasiliu-Oromulu, L. The study of thrips fauna from the romanian carpathians (Insecta: Thysanoptera). Acta Zool. Univ. Comen. 2004, 46, 95–106. [Google Scholar]
- Vasiliu-Oromulu, L. Thysanoptera (Insecta: Thysanoptera) populations, bioindicators on mining spoil material sites in the Retezat and Tarcu-Godeanu Massifs. Entomol. Rom. 2007, 12, 325–341. [Google Scholar]
- Mound, L.A.; Kibby, G. Thysanoptera: An Identification Guide; CAB international: Wallingford, NY, USA, 1998. [Google Scholar]
- Zur Strassen, R. Die Terebranten Thysanopteren Europas und des Mittelmer Gebietes; Goecke and Evers: Keltern, Germany, 2003. [Google Scholar]
- Schliephake, G.; Klimt, K. Thysanoptera. Die Tierwald Deutschland; G. Fischer Verlag: Jena, Germany, 1979. [Google Scholar]
- Kobro, S.; Rafoss, T. Identification of adult males and females of Hoplothrips species (Thysanoptera: Tubulifera) known from Norway, and some deductions on their life history. Entomol. Fenn. 2006, 17, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Spellerberg, I.F.; Fedor, P.J. A tribute to Claude Shannon (1916–2001) and a plea for more rigorous use of species richness, species diversity and the ‘Shannon–Wiener’ Index. Glob. Ecol. Biogeogr. 2003, 12, 177–179. [Google Scholar] [CrossRef] [Green Version]
- Fedor, P.; Zvaríková, M. Biodiversity indices. Encycl. Ecol. 2019, 2, 337–346. [Google Scholar]
- Oksanen, J. Multivariate Analysis of Ecological Communities in R: Vegan Tutorial 2015. Available online: https://www.mooreecology.com/uploads/2/4/2/1/24213970/vegantutor.pdf (accessed on 30 June 2022).
- Wohlgemuth, T.; Kull, P.; Wüthrich, H. Disturbance of microsites and early tree regeneration after windthrow in Swiss mountain forests due to the winter storm Vivian 1990. For. Snow Landsc. Res. 2002, 77, 17–47. [Google Scholar]
- Connell, J.H. Intermediate-Disturbance Hypothesis. Science 1979, 204, 1345. [Google Scholar] [CrossRef]
- Duelli, P. Mosaikkonzept und Inseltheorie in der Kulturlandschaft. Verh. Ges. Ökol. 1992, 21, 379–384. [Google Scholar]
- Čuchta, P.; Miklisová, D.; Kováč, Ľ. A three-year study of soil Collembola communities in spruce forest stands of the High Tatra Mts (Slovakia) after a catastrophic windthrow event. Eur. J. Soil Biol. 2012, 50, 151–158. [Google Scholar] [CrossRef]
- Čuchta, P.; Miklisová, D.; Kováč, Ľ. The succession of soil Collembola communities in spruce forests of the High Tatra Mountains five years after a windthrow and clear–cut logging. For. Ecol. Manag. 2019, 433, 504–513. [Google Scholar] [CrossRef]
- Lošková, J.; Ľuptáčik, P.; Miklisová, D.; Kováč, Ľ. The effect of clear-cutting and wildfire on soil Oribatida (Acari) in windthrown stands of the High Tatra Mountains (Slovakia). Eur. J. Soil Biol. 2013, 55, 131–138. [Google Scholar] [CrossRef]
- Urbanovičová, V.; Miklisová, D.; Kováč, Ľ. Forest Disturbance Enhanced the Activity of Epedaphic Collembola in Windthrown Stands of the High Tatra Mountains. J. Mt. Sci. 2014, 11, 449–463. [Google Scholar] [CrossRef]
- Kováč, Ľ.; Urbanovičová, V. Activity of epigeic arthropods in differently managed windthrown forest stands in the High Tatra Mts. North-West. J. Zool. 2014, 10, 337–345. [Google Scholar]
- Wermelinger, B.; Duelli, P.; Obrist, M.K. Dynamics of saproxylic beetles (Coleoptera) in windthrow areas in alpine spruce forests. For. Snow Landsc. Res. 2002, 77, 133–148. [Google Scholar]
- Thorn, S.; Bässler, C.; Gottschalk, T.; Hothorn, T.; Bussler, H.; Raffa, K.; Müller, J. New Insights into the Consequences of Post-Windthrow Salvage Logging Revealed by Functional Structure of Saproxylic Beetles Assemblages. PLoS ONE 2014, 9, e101757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wermelinger, B.; Duelli, P.; Obrist, M.K. Windthrow stimulates arthropod biodiversity in forests. In Dead Wood: A Key to Biodiversity, Proceedings of the International Symposium, Mantova, Italy, 29–31 May 2003; WSL Swiss Federal Research Institute: Birmensdorf, Switzerland, 2003; pp. 79–82. [Google Scholar]
- Sierka, W.; Halgoš, J. Thrips (Insecta, Thysanoptera) of the NPR Šúr near Bratislava. Entomofauna Carpath. 2003, 15, 14–19. [Google Scholar]
- Pelikán, J. Vertical distribution of alpine Thysanoptera. Folia Entomol. Hung. 1996, 57, 121–125. [Google Scholar]
- Conti, B. Notes on the presence of Aeolothrips intermedius in northwestern Tuscany and on its development under laboratory conditions. Bull. Insectology 2009, 62, 107–112. [Google Scholar]
- Kucharczyk, H.; Zawirska, I. Materiały do poznania wciornastków (Thysanoptera, Insecta) wyżyny luelskiej i niecki nidziańskej. Rocz. Świętokrzyski. Ser. B Nauk. Przyr. 1997, 24, 61–73. [Google Scholar]
- Den Belder, E.; Elderson, J.; Van Den Brink, W.J.; Schelling, G. Effect of woodlots on thrips density in leek fields: A landscape analysis. Agric. Ecosyst. Environ. 2002, 91, 139–145. [Google Scholar] [CrossRef]
- Trdan, S.; Valič, N.; Žnidarčič, D. Field efficacy of deltamethrin in reducing damage caused by Thrips tabaci Lindeman (Thysanoptera: Thripidae) on early white cabbage. J. Pest Sci. 2007, 80, 217–223. [Google Scholar] [CrossRef]
- Jenser, G. Studies on the vertical distribution of some Thysanoptera species in an oak forest. Int. J. Pure Appl. Zool. 1993, 4, 233–238. [Google Scholar]
- Pelikán, J. Thysanoptera. Terrestrial Invertebrates of the Pálava Biosphere Reserve of UNESCO. Folia Fac. Sci. Nat. Univ. Masaryk. Brun. Biológia 1995, 92, 137–146. [Google Scholar]
- Kucharczyk, H.; Zawirska, I.; Malczewska, E. Thrips (Thysanoptera, Insecta) of the Babia Góra Massif (Western Carpathians, Poland). Acta Phytopathol. Et Entomol. Hung. 2008, 43, 307–315. [Google Scholar] [CrossRef]
- Pelikán, J. Poznámky k znalosti třásněnek Rejvízských rašelinišť. Entomol. Listy 1947, 10, 1–16. [Google Scholar]
- Jenser, G.; Vasiliu-Oromulu, L.; Orbán, K.; Syénasi, A. Thysanoptera from Transylvania (Insecta: Thysanoptera). Entomol. Rom. 2003, 7, 62–78. [Google Scholar]
- Kucharczyk, H. Wciornastki (Insecta: Thysanoptera) jako element monitoringu ekologicznego w puszczy Białowieskiej. Leśne Pr. Badaw. 2004, 3, 85–94. [Google Scholar]
- Kucharczyk, H. Materiały do poznania wciornastków Puszczy Białowieskiej. Parki Nar. I Rezerw. Przyr. 1999, 18, 87–92. [Google Scholar]
- Kettunen, J.; Kobro, S.; Martikainen, P. Thrips (Thysanoptera) from dead aspen (Populus tremula) trees in Eastern Finland. Entomol. Fenn. 2005, 16, 246–250. [Google Scholar] [CrossRef]
- Kratochvíl, J.; Farský, O. Das Absterben der diesjahrigen terminalen Larchentriebe. Z. Für Angew. Entomol. 1941, 29, 177–218. [Google Scholar] [CrossRef]
- Lewis, T. Thrips as Crop Pests; CAB International: Oxford, UK, 1997. [Google Scholar]
- Maksymov, J.K. Die Überwinterung des Lärchenblasenfusses Taeniothrips laricivorus Kratochvil und Farsky. Mitt. Schweiz. Anst. Für D. Forstl. Vers. 1965, 41, 1–17. [Google Scholar]
- Zenther-Møller, O. Taeniothrips laricivorus (Krat.) (Thripidae, Thysanoptera) in Danish stands of Larix decidua (Mill.). Oikos 1965, 16, 58–59. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EXT | FIR | NEX1 | NEX2 | LUK1 | LUK2 | LES1 | LES2 | |
|---|---|---|---|---|---|---|---|---|
| Coordinates | 49°07′15.60″ N | 49°08′10.86″ N | 49°09′41.34″ N | 49°10′40.20″ N | 49°10′30.02″ N | 49°10′27.08″ N | 49°10′29.56″ N | 49°10′30.51″ N |
| 20°09′49.68″ E | 20°11′57.00″ E | 19°55′26.88″ E | 19°55′02.34″ E | 20°14′48.64″ E | 20°14′49.52″ E | 20°14′46.81″ E | 20°14′49.64″ E | |
| Altitude (m.a.s.l.) | 1060 | 1080 | 1006 | 1052 | 1050 | 1050 | 1050 | 1050 |
| Area (ha) | 93 | 90 | 11 | 21.4 | 0.2 | 0.2 | 66 | 1 |
| Age | 4 | 4 | 5 | 5 | 40 | 40 | 140 | 140 |
| Management | cleared | fire + cleared | not cleared | not cleared | mow | mow | no | no |
| Fallen trees | 100% | 100% | 40% | 90% | 0% | 0% | 0% | 0% |
| Affected by windstorm | Not affected by windstorm | |||||||
| Wood decay | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 |
| Same-age trees | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
| Management | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 |
| Spruce % | 1 | 1 | 1 | 1 | 0 | 0 | 0 | 0 |
| Fire | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 |
| Conservation | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 1 |
| Anthropogenic impact | 5 | 6 | 3 | 3 | 3 | 3 | 0 | 1 |
| Variable | Description | NMDS1 | NMDS2 | r2 | Pr(>r) | Stress | Code |
|---|---|---|---|---|---|---|---|
| Diversity | |||||||
| Abundance | the number of thrips specimen per study plot | 0.9378 | −0.3473 | 0.0401 | 0.180 | 0.1271 | |
| Eve_Shan | equitability based on the Shannon index | 0.5661 | 0.8243 | 0.1125 | 0.008 | 0.1271 | ** |
| Eve_Simp | equitability based on the Simpson index | −0.9558 | 0.2939 | 0.0441 | 0.149 | 0.12722 | |
| Shannon | Shannon–Wiener index of diversity | 0.7051 | 0.7091 | 0.1492 | 0.002 | 0.1272 | ** |
| Simpson | a reciprocal Simpson diversity | 0.5362 | 0.8441 | 0.0330 | 0.245 | 0.1271 | |
| Spe_rich | the number of thrips species per study plot | 0.8963 | 0.4435 | 0.2559 | 0.001 | 0.1273 | *** |
| Guild ecology | |||||||
| Arb | percentage representation of species belonging to the tree-living species | −0.6883 | 0.7254 | 0.0495 | 0.118 | 0.1271 | |
| Cor_arb | percentage representation of species belonging to the bark-dwelling species | 0.7861 | 0.6181 | 0.0065 | 0.757 | 0.1271 | |
| Flo | percentage representation of species belonging to the flower-living species | −0.4184 | −0.9083 | 0.1618 | 0.001 | 0.1282 | *** |
| Fol | percentage representation of species belonging to the leaf-living species | 0.8700 | 0.4930 | 0.0611 | 0.073 | 0.1271 | |
| Fol_arb | percentage representation of species belonging to the leaf-living species on trees | 0.6562 | 0.7546 | 0.1446 | 0.003 | 0.1273 | ** |
| Gra | percentage representation of species belonging to the grass-living species | −0.8320 | 0.5547 | 0.0084 | 0.690 | 0.1274 | |
| Nm_guilds | the number of guilds per study plot | 0.6213 | 0.7836 | 0.0285 | 0.299 | 0.1280 | |
| Stand variables and anthropogenic impact | |||||||
| Age | the age of the stand expressed in years | −0.4476 | 0.8942 | 0.0457 | 0.139 | 0.1277 | |
| Area | area of study plot (in hectares) | −0.5999 | −0.8000 | 0.3327 | 0.001 | 0.1272 | *** |
| Calamity | the presence of windthrow (1-present, 0-absent) | 0.5430 | −0.8398 | 0.0217 | 0.407 | 0.1279 | |
| Defoliation_NFC | the degree of defoliation according to National Forestry Centre | 0.7180 | −0.6960 | 0.0118 | 0.617 | 0.1271 | |
| E1 | the cover of herbs | 0.3671 | −0.9302 | 0.0498 | 0.116 | 0.1270 | |
| E2 | the cover of shrubs | 0.8569 | 0.5154 | 0.3691 | 0.001 | 0.1274 | *** |
| E3 | the cover of trees | −0.0790 | 0.9969 | 0.1067 | 0.010 | 0.1272 | ** |
| Edge_effect | distance (in meters) of the trap from the neighboring habitat | −0.6497 | −0.7602 | 0.3879 | 0.001 | 0.1273 | *** |
| Exposure | exposure to the sun | −0.8814 | −0.4724 | 0.3491 | 0.001 | 0.1272 | *** |
| Health_NFC | the degree of forest health according to National Forestry Centrum | −0.1813 | −0.9834 | 0.0561 | 0.089 | 0.1275 | |
| Heterogeneity | habitat heterogeneity (0-weak to 1-greater) | 0.6704 | 0.7420 | 0.3975 | 0.001 | 0.1276 | *** |
| Plant_com | the type of the plant community | −0.4693 | 0.8831 | 0.0454 | 0.140 | 0.1273 | |
| Plant_height | the height of the stand in the study plot | 0.1812 | 0.9835 | 0.2013 | 0.001 | 0.1274 | *** |
| Slope | slope of the study plot (in degrees) | −0.9988 | 0.0488 | 0.1221 | 0.004 | 0.1271 | ** |
| Spe_rich_tree | the number of tree species per study plot | −0.6485 | −0.7612 | 0.0566 | 0.087 | 0.1274 | |
| Tree_thick | the thickness of trees at the study plot | 0.1374 | 0.9905 | 0.1823 | 0.001 | 0.1280 | *** |
| Ant_imp | anthropogenic impact based on six negative points of anthropogenic activity | −0.6534 | −0.7570 | 0.1718 | 0.001 | 0.1270 | *** |
| Climatic factors | |||||||
| C | clouds amount per study plot | 0.9972 | 0.0747 | 0.0253 | 0.329 | 0.1273 | |
| H | average relative air humidity during the vegetation season per study plot | 0.5979 | 0.8016 | 0.2723 | 0.001 | 0.1272 | *** |
| P | the average precipitation during the growing season per study plot | 0.8202 | −0.5721 | 0.0142 | 0.548 | 0.1273 | |
| T | average air temperature during the vegetation season per study plot | −0.8266 | −0.5628 | 0.3532 | 0.001 | 0.1275 | *** |
| Species/Study Plot | EXT | FIR | NEX1 | NEX2 | LUK1 | LUK2 | LES1 | LES2 |
|---|---|---|---|---|---|---|---|---|
| Aeolothrips albicinctus | + | |||||||
| Aeolothrips ericae | + | |||||||
| Aeolothrips intermedius | + | + | + | + | ||||
| Aeolothrips vittatus | + | |||||||
| Anaphothrips obscurus | + | + | ||||||
| Aptinothrips stylifer | + | |||||||
| Bolacothrips jordani | + | + | + | |||||
| Dendrothrips saltatrix | + | |||||||
| Frankliniella intonsa | + | + | ||||||
| Frankliniella tenuicornis | + | + | ||||||
| Chirothrips hamatus | + | |||||||
| Chirothrips manicatus | + | + | + | + | + | + | + | |
| Limothrips denticornis | + | + | + | + | ||||
| Mycterothrips albidicornis | + | |||||||
| Mycterothrips latus | + | |||||||
| Neohydatothrips gracilicornis | + | |||||||
| Odontothrips intermedius | + | |||||||
| Odontothrips loti | + | + | ||||||
| Oxythrips ajugae | + | + | + | + | + | |||
| Oxythrips bicolor | + | + | ||||||
| Rubiothrips silvarum | + | |||||||
| Sericothrips bicornis | + | + | ||||||
| Stenothrips graminum | + | + | ||||||
| Taeniothrips inconsequens | + | |||||||
| Taeniothrips picipes | + | + | ||||||
| Thrips angusticeps | + | + | ||||||
| Thrips atratus | + | + | ||||||
| Thrips brevicornis | + | + | + | |||||
| Thrips discolor | + | + | ||||||
| Thrips flavus | + | + | + | + | + | + | ||
| Thrips fuscipennis | + | + | + | + | ||||
| Thrips major | + | + | ||||||
| Thrips minutissimus | + | + | + | + | + | |||
| Thrips physapus | + | + | + | + | ||||
| Thrips pini | + | + | + | + | + | + | ||
| Thrips praetermissus | + | |||||||
| Thrips tabaci | + | + | + | + | + | + | + | + |
| Thrips trybomi | + | |||||||
| Thrips validus | + | |||||||
| Thrips vulgatissimus | + | |||||||
| Cryptothrips nigripes | + | |||||||
| Haplothrips aculeatus | + | |||||||
| Hoplothrips pedicularius | + | + | + | + | ||||
| Hoplothrips polystici | + | |||||||
| Hoplothrips ulmi | + | |||||||
| Hoplothrips unicolor | + | |||||||
| Liothrips setinodis | + | |||||||
| Xylaplothrips fuliginosus | + | + |
| EXT | FIR | NEX1 | NEX2 | LUK1 | LUK2 | LES1 | LES2 | |
|---|---|---|---|---|---|---|---|---|
| Spe_rich | 6 | 7 | 14 | 20 | 20 | 16 | 14 | 15 |
| Shannon | 0.6584 | 0.5300 | 1.4008 | 1.7642 | 2.3046 | 2.3042 | 1.9610 | 1.4630 |
| Simpson | 1.4146 | 1.3179 | 2.7297 | 2.8810 | 6.4968 | 7.1111 | 4.1538 | 2.3180 |
| Eve_Shannon | 0.3675 | 0.2724 | 0.5308 | 0.5889 | 0.7693 | 0.8311 | 0.7431 | 0.5544 |
| Eve_Simpson | 0.2358 | 0.1883 | 0.1950 | 0.1441 | 0.3248 | 0.4444 | 0.2967 | 0.1656 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masarovič, R.; Zvaríková, M.; Zvarík, M.; Majzlan, O.; Prokop, P.; Fedor, P. Changes in Diversity and Structure of Thrips (Thysanoptera) Assemblages in the Spruce Forest Stands of High Tatra Mts. after a Windthrow Calamity. Insects 2022, 13, 670. https://doi.org/10.3390/insects13080670
Masarovič R, Zvaríková M, Zvarík M, Majzlan O, Prokop P, Fedor P. Changes in Diversity and Structure of Thrips (Thysanoptera) Assemblages in the Spruce Forest Stands of High Tatra Mts. after a Windthrow Calamity. Insects. 2022; 13(8):670. https://doi.org/10.3390/insects13080670
Chicago/Turabian StyleMasarovič, Rudolf, Martina Zvaríková, Milan Zvarík, Oto Majzlan, Pavol Prokop, and Peter Fedor. 2022. "Changes in Diversity and Structure of Thrips (Thysanoptera) Assemblages in the Spruce Forest Stands of High Tatra Mts. after a Windthrow Calamity" Insects 13, no. 8: 670. https://doi.org/10.3390/insects13080670