
Predatory and Parasitic Insects Associated with Urophora cardui L. (Diptera: Tephritidae) Galls on Canada Thistle, Cirsium arvense L. (Asterales, Asteraceae) in North Dakota

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Site Description and Sampling Methods
2.2. Statistical Analyses
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- White, I.M.; Korneyev, V.A. A revision of the western Palaearctic species of Urophora Robineau-Desvoidy (Diptera: Tephritidae). Syst. Entomol. 1989, 14, 327–374. [Google Scholar] [CrossRef] [PubMed]
- Korneyev, V.A.; White, I.M. Fruit flies of the genus Urophora R.-D. (Diptera, Tephritidae) of eastern Palaearctics. II. Review of species of the subgenus Urophora s. str. Communication 3. Entomol. Rev. 1996, 76, 499–513. [Google Scholar]
- Frenzel, M.; Eber, S.; Klotz, S.T.; Brandl, R. Ecological comparisons across geographical distributions: The thistle gall fly Urophora cardui (Diptera: Tephritidae) on two different Cirsium hosts. Eur. J. Entomol. 2000, 97, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Cripps, M.G.; Gassmann, A.; Fowler, S.V.; Bourdȏt, G.W.; McClay, A.S.; Edwards, G.R. Classical biological control of Cirsium arvense: Lessons from the past. Biol. Control. 2011, 57, 165–174. [Google Scholar] [CrossRef]
- Winston, R.L.; Schwarzländer, M.; Hinz, H.L.; Day, M.D.; Cock, M.J.W.; Julien, M.H. Biological Control of Weeds: A World Catalogue of Agents and Their Target Weeds, 5th ed.; FHTET-2014-04; USDA Forest Service, Forest Health Technology Enterprise Team: Morgantown, WV, USA, 2014. [Google Scholar]
- Peschken, D.P.; Finnamore, D.B.; Watson, A.K. Biocontrol of the weed Canada thistle (Cirsium arvense): Releases and development of the gall fly Urophora cardui (Diptera: Tephritidae) in Canada. Can. Entomol. 1982, 114, 349–357. [Google Scholar] [CrossRef]
- Piper, G.L.; Andres, L.A. Canada thistle. In Biological Control in the Western United States: Accomplishments and Benefits of Regional Research Project W-84, 1964–1989; Nechols, J.R., Andres, L.A., Beardsley, J.W., Goeden, R.D., Jackson, C.G., Eds.; Publication 3361; University of California, Division of Agriculture and Natural Resources: Oakland, CA, USA, 1995; pp. 233–237. [Google Scholar]
- Peschken, D.P.; Harris, P. Host specificity and biology of Urophora cardui (Diptera: Tephritidae). A biocontrol agent for Canada thistle (Cirsium arvense). Can. Entomol. 1975, 107, 1101–1110. [Google Scholar] [CrossRef]
- Eber, S.; Brandl, R. Ecological and genetic spatial patterns of Urophora cardui (Diptera: Tephritidae) as evidence for population structure and biogeographical processes. J. Anim. Ecol. 1994, 63, 187–199. [Google Scholar] [CrossRef]
- Price, J.R. Post-Release Assessment of Classical Biological Control of Canada Thistle (Cirsium arvense) in the Western United States. Master’s Thesis, University of Idaho, Moscow, ID, USA, 2014. [Google Scholar]
- Harris, P.; Shorthouse, J.D. Effectiveness of gall inducers in weed biological control. Can. Entomol. 1996, 128, 1021–1055. [Google Scholar] [CrossRef] [Green Version]
- Lalonde, R.G.; Shorthouse, J.D. Developmental morphology of the gall of Urophora cardui (Diptera, Tephritidae) in the stems of Canada thistle (Cirsium arvense). Can. J. Bot. 1984, 62, 1372–1384. [Google Scholar] [CrossRef]
- Lalonde, R.G.; Shorthouse, J.D. Exit strategy of Urophora cardui (Diptera: Tephritidae) from its gall on Canada thistle. Can. Entomol. 1982, 114, 873–878. [Google Scholar] [CrossRef]
- Freese, G.; Zwölfer, H. The problem of optimal clutch size in a tritrophic system: The oviposition strategy of the thistle gallfly Urophora cardui (Diptera: Tephritidae). Oecologia 1996, 108, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Shorthouse, J.D.; Lalonde, R.G. Role of Urophora cardui (L.) (Diptera: Tephritidae) in growth and development of its gall on stems of Canada thistle. Can. Entomol. 1988, 120, 639–646. [Google Scholar] [CrossRef]
- Lalonde, R.G.; Shorthouse, J.D. Growth and development of larvae and galls of Urophora cardui (Diptera: Tephritidae) on Cirsium arvense (Compositae). Oecologia 1985, 65, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Zwölfer, H.; Arnold-Rinehart, J. The evolution of interactions and diversity in plant–insect systems: The Urophora Eurytoma food web in galls on Palearctic Cardueae. In Biodiversity and Ecosystem Function; Schulze, E.D., Mooney, H.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1994; pp. 211–229. [Google Scholar]
- Johannesen, J.; Seitz, A. Larval distributions of the ectoparasitoid wasp Eurytoma robusta relative to the host tephritid gall fly Urophora cardui. Entomol. Exp. Appl. 2003, 107, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Price, P.W.; Wilson Fernandes, G.; Waring, G.L. Adaptive nature of insect galls. Environ. Entomol. 1987, 16, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Peschken, D.P.; Derby, J.L. Establishment of Urophora cardui (Diptera: Tephritidae) on Canada thistle, Cirsium arvense (Asteraceae), and colony development in relation to habitat and parasitoids in Canada. In Ecological Studies; Vertical Food Web Interactions; Dettner, K., Bauer, G., Völkl, W., Eds.; Springer: Berlin/Heidelberg, Germany, 1997; Volume 130, pp. 53–66. [Google Scholar] [CrossRef]
- Zwölfer, H.; Böheim, M.; Beck, E. Eurytoma serratulae and E. robusta (Hymenoptera, Eurytomidae): Complementary host exploitation strategies of coexisting parasitoids and their impact on the host Urophora cardui. J. Hymenopt. Res. 2015, 42, 47–62. [Google Scholar] [CrossRef] [Green Version]
- Prischmann-Voldseth, D.A.; Gramig, G.G.; Burns, E.E. Home on the range: Establishment of a Canada thistle biocontrol agent. Rangel 2012, 34, 2–5. [Google Scholar] [CrossRef] [Green Version]
- Prischmann-Voldseth, D.A.; Burns, E.E.; Swenson, S.; Gramig, G.G. Life history and phenology of an endophagous stem-mining herbivore (Coleoptera: Curculionidae, Hadroplontus litura) of a clonal weed. Ann. Entomol. Soc. Am. 2016, 109, 12–21. [Google Scholar] [CrossRef]
- Klimmek, F.; Baur, H. An interactive key to Central European species of the Pteromalus albipennis species group and other species of the genus (Hymenoptera: Chalcidoidea: Pteromalidae), with the description of a new species. Biodivers. Data J. 2018, 6, e27722. [Google Scholar] [CrossRef]
- Gibson, G.A.P.; Huber, J.T.; Woolley, J.B. Annotated Keys to the Genera of Nearctic Chalcidoidea (Hymenoptera); NRC Research Press: Ottawa, ON, Canada, 1997. [Google Scholar]
- Talebi, A.A.; Farahani, S.; Pirouzeh, F.Z. Six new record species of the genus Schizoprymnus Förster, 1862 (Hymenoptera: Braconidae, Brachistinae). Iran. Entomofauna 2018, 39, 325–333. [Google Scholar]
- Knull, J.N. The checkered beetles of Ohio (Coleoptera: Cleridae). Ohio Biol. Surv. Bull. 1951, 8, 268–350. [Google Scholar]
- Opitz, W. Family 73. Cleridae Latreille 1804. In American Beetles, Polyphaga: Scarabaeoidea through Curculionoidea; Arnett, R.H., Jr., Thomas, M.C., Skelley, P.E., Frank, J.H., Eds.; CRC Press: Boca Raton, FL, USA, 2002; Volume 2, pp. 267–280. [Google Scholar]
- Critchlow, D.E.; Fligner, M.A. On distribution-free multiple comparisons in the one-way analysis of variance. Commun. Stat.-Theor. M. 1991, 20, 127–139. [Google Scholar] [CrossRef]
- Neuhäuser, M.; Bretz, F. Nonparametric all-pairs multiple comparisons. Biometr. J. 2001, 43, 571–580. [Google Scholar] [CrossRef]
- Böving, A.G.; Champlain, A.B. Larvae of North American beetles of the family Cleridae. U. S. Natl. Mus. Proc. 1920, 57, 575–649. [Google Scholar] [CrossRef] [Green Version]
- Sabrosky, C.W. Notes on the larvae and larval habit of Isohydnocera curtipennis (Newmn.) (Coleoptera: Cleridae). J. Kans. Entomol. Soc. 1934, 7, 65–68. [Google Scholar]
- Clausen, C.P. Entomophagous Insects; McGraw-Hill Book Co.: New York, NY, USA, 1940. [Google Scholar]
- Morrill, W.L.; Weaver, D.K.; Irish, N.J.; Barr, W.F. Phyllobaenus dubius (Wolcott) (Coleoptera: Cleridae), a new record of a predator of the wheat stem sawfly (Hymenoptera: Cephidae). J. Kans. Entomol. Soc. 2001, 74, 181–183. [Google Scholar] [CrossRef]
- Osten Sacken, R. On the Cynipidae of the North American oaks and their galls. Proc. Entomol. Soc. Phil. 1861, 1, 47–72. [Google Scholar]
- Leavengood, J.M., Jr. The Checkered Beetles (Coleoptera: Cleridae) of Florida. Master’s Thesis, University of Florida, Gainesville, FL, USA, 2008. [Google Scholar]
- Baker, C.F. Biological notes’ on some Colorado Coleoptera. Entomol. News. 1895, 6, 27–29. [Google Scholar]
- De Smet-Moens, H. The Insect Fauna of Canada Thistle, Cirsium arvense (L.) Scop. in Southern Montana. Master’s Thesis, Montana State University, Bozeman, MT, USA, 1982. [Google Scholar]
- SCAN (Symbiota Collections of Arthropods Network). Available online: www.scan-bugs.org (accessed on 8 June 2021).
- GBIF.org. Global Biodiversity Information Facility. GBIF Occurrence Download. Available online: https://doi.org/10.15468/dl.89asaz (accessed on 7 June 2021).
- GBIF Secretariat. Isohydnocera tabida (LeConte, 1849). GBIF Backbone Taxonomy. Checklist Dataset. Available online: https://doi.org/10.15468/39omei (accessed on 7 June 2021).
- Chapin, E.A. Studies in the Genus Hydnocera Newman: The Species of the New England States. Master’s Thesis, University of Massachusetts, Amherst, MA, USA, 1917. [Google Scholar]
- Wolcott, A.B. Catalogue of North American beetles of the family Cleridae. Fieldiana Zool. 1947, 32, 61–105. [Google Scholar] [CrossRef]
- Davis, L.V.; Gray, I.E. Zonal and seasonal distribution of insects in North Carolina salt marshes. Ecol. Monogr. 1966, 36, 275–295. [Google Scholar] [CrossRef]
- Rykken, J.J.; Farrell, B.D. Boston Harbor Islands all taxa biodiversity inventory: Discovering the “microwilderness” of an urban island park. In Natural Resource Technical Report NPS/BOHA/NRTR—2013/746; National Park Service: Fort Collins, CO, USA, 2013. [Google Scholar]
- Dorshorst, J.J.; Young, D.K. An annotated checklist of Wisconsin checkered beetles (Coleoptera: Cleridae and Thanerocleridae). Gr. Lakes Entomol. 2008, 41, 169–184. [Google Scholar]
- Mawdsley, J.R. Ecological notes on species of Cleridae (Insecta: Coleoptera) associated with the prairie flora of central North America. Gt. Lakes Entomol. 2002, 35, 15–22. [Google Scholar]
- Wilhelm, G.; Rericha, L. Inventory and Assessment Plan for Stewardship and Monitoring at Hitchcock Nature Center; Hitchcock Nature Center: Honey Creek, IA, USA, 2012. [Google Scholar]
- Dailey, P.J.; Graves, R.C.; Kingsolver, J.M. Survey of Coleoptera collected on the common milkweed, Asclepias syriaca, at one site in Ohio. Coleopt. Bull. 1978, 32, 223–229. [Google Scholar]
- Böving, A.G.; Craighead, F.C. An Illustrated Synopsis of the Principal Larval Forms of the Order Coleoptera; The Brooklyn Entomological Society: Brooklyn, NY, USA, 1931. [Google Scholar] [CrossRef]
- Leng, C.W.; Davis, W.T. List of the Coleoptera of Staten Island, New York. In Proceedings of the Staten Island Institute Arts Sciences; Lancaster Press Incorporated: Lancaster, PA, USA, 1924; Volume 2, pp. 1–82. [Google Scholar]
- Tilden, J.W. Notes on the habits of Phyllobaenus scabra (Le Conte) (Cleridae). Coleopt. Bull. 1950, 4, 17–19. [Google Scholar]
- László, Z.; Sólyom, K.; Prázsmári, H.; Barta, Z.; Tóthmérész, B. Predation on rose galls: Parasitoids and predators determine gall size through directional selection. PLoS ONE 2014, 9, e99806. [Google Scholar] [CrossRef] [Green Version]
- Tscharntke, T. Cascade effects among four trophic levels: Bird predation on galls affects density-dependent parasitism. Ecology 1992, 73, 1689–1698. [Google Scholar] [CrossRef]
- Hoebeke, E.R.; Wheeler, A.G., Jr. Pteromalus elevatus (Walker) (Hymenoptera: Pteromalidae): North American records of an immigrant parasitoid of the gall fly Urophora jaceana (Diptera: Tephritidae). Proc. Entomol. Soc. Wash. 1996, 98, 87–92. [Google Scholar]
- Jones, T.H.; Godfray, H.C.J.; Hassell, M.P. Relative movement patterns of a tephritid fly and its parasitoid wasps. Oecologia 1996, 106, 317–324. [Google Scholar] [CrossRef]
- Duan, J.J.; Messing, R.H. Effect of Tetrastichus giffardianus (Hymenoptera: Eulophidae) on nontarget flowerhead-feeding tephritids (Diptera: Tephritidae). Environ. Entomol. 1998, 27, 1022–1028. [Google Scholar] [CrossRef]
- Vikberg, V. Orakärpänen (Urophora cardui) ja sen kiilukaisloiset Etelä-Suomessa (Hymenoptera: Chalcidoidea: Eulophidae, Eupelmidae, Eurytomidae, Pteromalidae, Torymidae). Sahlbergia 2005, 10, 30–49. [Google Scholar]
- Johannesen, J. Tracing the history and ecological context of Wolbachia double infection in a specialist host (Urophora cardui)—parasitoid (Eurytoma serratulae) system. Ecol. Evol. 2017, 7, 986–996. [Google Scholar] [CrossRef] [PubMed]
- Story, J.M.; DeSmet-Moens, H.; Morrill, W.L. Phytophagous insects associated with Canada thistle, Cirsium arvense (L.) Scop., in southern Montana. J. Kans. Entomol. Soc. 1985, 58, 472–478. [Google Scholar]
- Zwölfer, H.; Englert, W.; Pattullo, W. Investigations on the Biology, Population Ecology and the Distribution of Urophora cardui L. Weed Projects for Canada, Progress Report XXVII; Commonwealth Institute of Biological Control: Delémont, Switzerland, 1970. [Google Scholar]




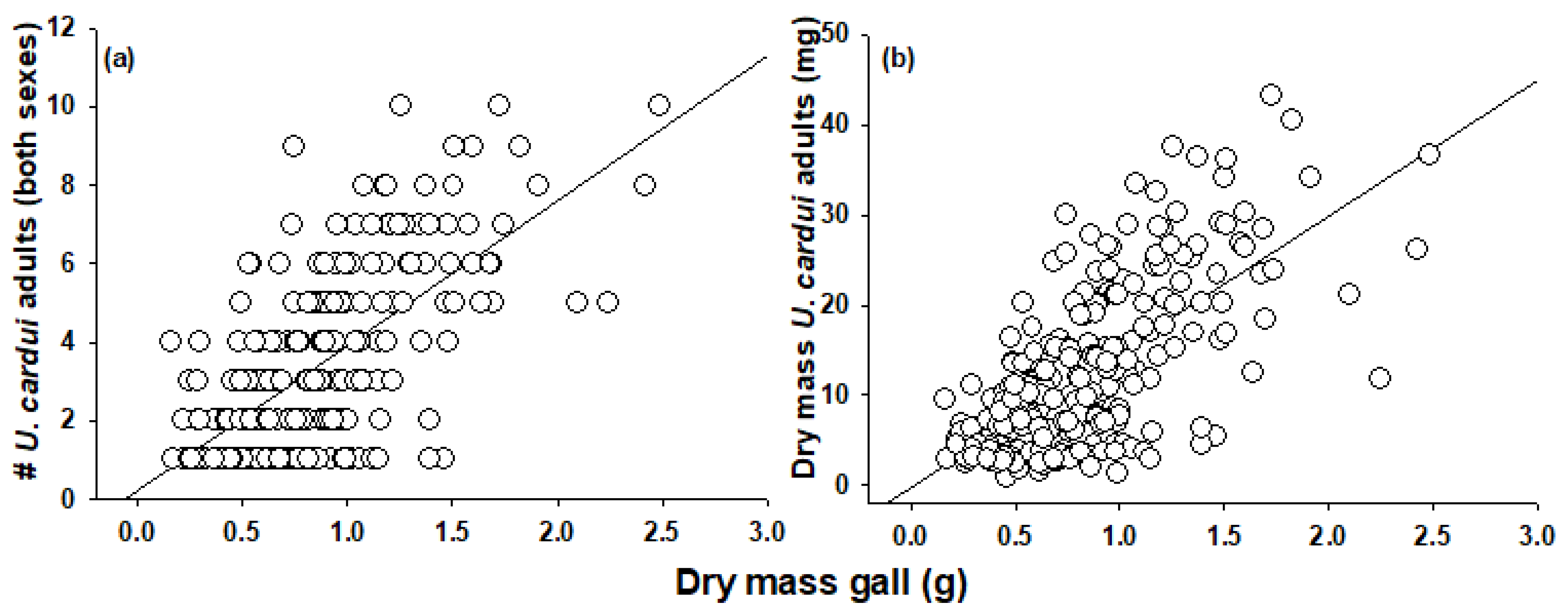{kind=link}
{kind=link}
{kind=link}
| Site 1 | ND 2 County | Latitude N, Longitude W (Decimal Degrees) | NDAWN 3 Weather Station | Avg. Air Temp. (°C) | Avg. Precipitation (cm) | Date Galls Collected 4 | ||
|---|---|---|---|---|---|---|---|---|
| May | Yearly | May | Yearly | |||||
| Tr_a | Trail | 47.577, 96.859 | Hillsboro ND | 13.3 | 5 | 6.86 | 54.99 | 4 May 2012 |
| Tr_b | Trail | 47.312904, 97.409602 | Mayville ND | 12.7 | 4.4 | 6.83 | 54.79 | 4 May 2012 |
| Ra | Ransom | 46.350056, 97.933539 | Lisbon ND | 13.9 | 6.1 | 7.49 | 53.75 | 2 May 2012 |
| Gr | Griggs | 47.514863, 98.366467 | Cooperstown ND | 12.2 | 3.9 | 7.42 | 54.89 | 4 May 2012 |
| St_a | Stark | 46.848512, 102.892616 | Dickinson ND | 11.7 | 5.6 | 5.89 | 42.49 | 10 May 2012 20 May 2013 |
| St_b | Stark | 46.849122, 102.892973 | Dickinson ND | 11.7 | 5.6 | 5.89 | 42.49 | |
| St_c | Stark | 46.854804, 102.893682 | Dickinson ND | 11.7 | 5.6 | 5.89 | 42.49 | |
| Sp | Slope | 46.294698, 103.634989 | Bowman ND | 11.7 | 6.1 | 6.27 | 39.65 | 11 May 2012 |
| Bo | Bowman | 46.223459, 103.702719 | Bowman ND | 11.7 | 6.1 | 6.27 | 39.65 | 11 May 2012 21 May 2013 |
| Site 1 | # Galls 2 | # Adult Flies Per Gall (Mean ± SE) 3 | Fly Biomass Per Gall (Mean ± SE mg dry wt.) 3 | # P. elevatus Per Gall (Mean ± SE) 4 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | V | +I | +U | +P | M and F | M | F | M and F | M and F | M | F | |
| 2012 | ||||||||||||
| Tr_a | 8 | 8 | 0 | 8 | 0 | 2.13 ± 0.52b | 1.00 ± 0.42ab | 1.13 ± 0.35ab | 6.36 ± 1.49b | n/a | n/a | n/a |
| Tr_b | 54 | 37 | 0 | 34 | 9 | 2.59 ± 0.33b | 1.44 ± 0.22ab | 1.15 ± 0.19b | 8.30 ± 1.09b | 1.64 ± 0.20a | 1.25 ± 0.16a | 1.20 ± 0.20a |
| Ra | 41 | 12 | 0 | 11 | 1 | 2.09 ± 0.39b | 0.46 ± 0.16b | 1.64 ± 0.34ab | 5.92 ± 1.08b | 4.00 ± 5a | 1.00 ± 5a | n/a |
| Gr | 38 | 37 | 0 | 36 | 1 | 4.64 ± 0.37a | 2.36 ± 0.26a | 2.28 ± 0.24a | 18.23 ± 1.49a | 2.00 ± 5a | 1.00 ± 5a | 1.00 ± 5a |
| St-a | 62 | 59 | 5 | 51 | 25 | 3.02 ± 0.34b | 1.29 ± 0.19b | 1.73 ± 0.24ab | 10.84 ± 1.27b | 1.81 ± 0.23a | 1.32 ± 0.19a | 1.31 ± 0.15a |
| St-b | 34 | 27 | 0 | 24 | 8 | 3.04 ± 0.41ab | 1.75 ± 0.27ab | 1.29 ± 0.22ab | 10.54 ± 1.42b | 1.75 ± 0.25a | 1.13 ± 0.13a | 1.25 ± 0.25a |
| St-c | 62 | 51 | 5 | 41 | 21 | 2.41 ± 0.25b | 0.81 ± 0.15b | 1.61 ± 0.22ab | 8.64 ± 0.90b | 1.86 ± 0.26a | 1.35 ± 0.17a | 1.23 ± 0.12a |
| Sp | 25 | 22 | 0 | 17 | 9 | 2.59 ± 0.49b | 1.44 ± 0.22ab | 1.24 ± 0.27ab | 9.51 ± 2.01b | 2.56 ± 0.50a | 1.78 ± 0.32a | 1.17 ± 0.17a |
| Bo | 44 | 41 | 1 | 41 | 3 | 4.66 ± 0.37a | 2.51 ± 0.30a | 2.15 ± 0.22a | 19.31 ± 1.58a | 1.00 ± 0.00a | 1.00 ± 0.00a | 1.00 ± 5a |
| 2013 | ||||||||||||
| St-a-c | 480 | 124 | 17 | 82 | 37 | 1.83 ± 0.15 | n/a | n/a | n/a | 1.71 ± 0.23 | n/a | n/a |
| Bo | 211 | 196 | 23 | 160 | 65 | 2.74 ± 0.15 | n/a | n/a | n/a | 1.92 ± 0.17 | n/a | n/a |
| State 1 | County | Location | Date | Year | Collector/Identifier | Other Label Data 3 |
|---|---|---|---|---|---|---|
| ND | Barnes | S. of V[alley] city | 6-vi | 1963 | D.[G.] Aarhus | - |
| ND | Ransom | - | 6-vi | 1963 | R.L. Post/D.E. Foster | - |
| ND | Sargent | Tewaukon NWR 2 | 4-vi | 1964 | D.G. Aarhus | - |
| SD | Brookings | Brookings | 28-vi | 1965 | V.M. Kirk | S[weeping], 2 spp. |
| SD | Brookings | White | 18 vii | 1966 | V.M. Kirk | - |
| SD | Union | Elk Point | 20-vi | 1968 | V.M. Kirk/W.F. Barr | - |
| SC | Charleston | Kiawah | 10-v | 1963 | V.M. Kirk | 2 spp. |
| SC | Charleston | Seabrook | 8-x | 1962 | V.M. Kirk/[J.N.] Knull | - |
| SC | Charleston/Colleton | Edisto Is. | 9-x | 1962 | V.M. Kirk | Sweeping beach grass, 2 spp. |
| SC | Georgetown | Pawley’s Is. | 7-viii | 1962 | V.M. Kirk | S[weeping], 2 spp. ‘159’ [?] |
| SC | Sumter | Sumter | 15-vii | 1964 | V.M. Kirk | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swenson, S.J.; Bell-Clement, J.; Schroeder, S.; Prischmann-Voldseth, D.A. Predatory and Parasitic Insects Associated with Urophora cardui L. (Diptera: Tephritidae) Galls on Canada Thistle, Cirsium arvense L. (Asterales, Asteraceae) in North Dakota. Insects 2022, 13, 646. https://doi.org/10.3390/insects13070646
Swenson SJ, Bell-Clement J, Schroeder S, Prischmann-Voldseth DA. Predatory and Parasitic Insects Associated with Urophora cardui L. (Diptera: Tephritidae) Galls on Canada Thistle, Cirsium arvense L. (Asterales, Asteraceae) in North Dakota. Insects. 2022; 13(7):646. https://doi.org/10.3390/insects13070646
Chicago/Turabian StyleSwenson, Stephanie J., Jonathan Bell-Clement, Sarah Schroeder, and Deirdre A. Prischmann-Voldseth. 2022. "Predatory and Parasitic Insects Associated with Urophora cardui L. (Diptera: Tephritidae) Galls on Canada Thistle, Cirsium arvense L. (Asterales, Asteraceae) in North Dakota" Insects 13, no. 7: 646. https://doi.org/10.3390/insects13070646