
Nutritional Composition of Bombyx mori Pupae: A Systematic Review

 , , , , and
, , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
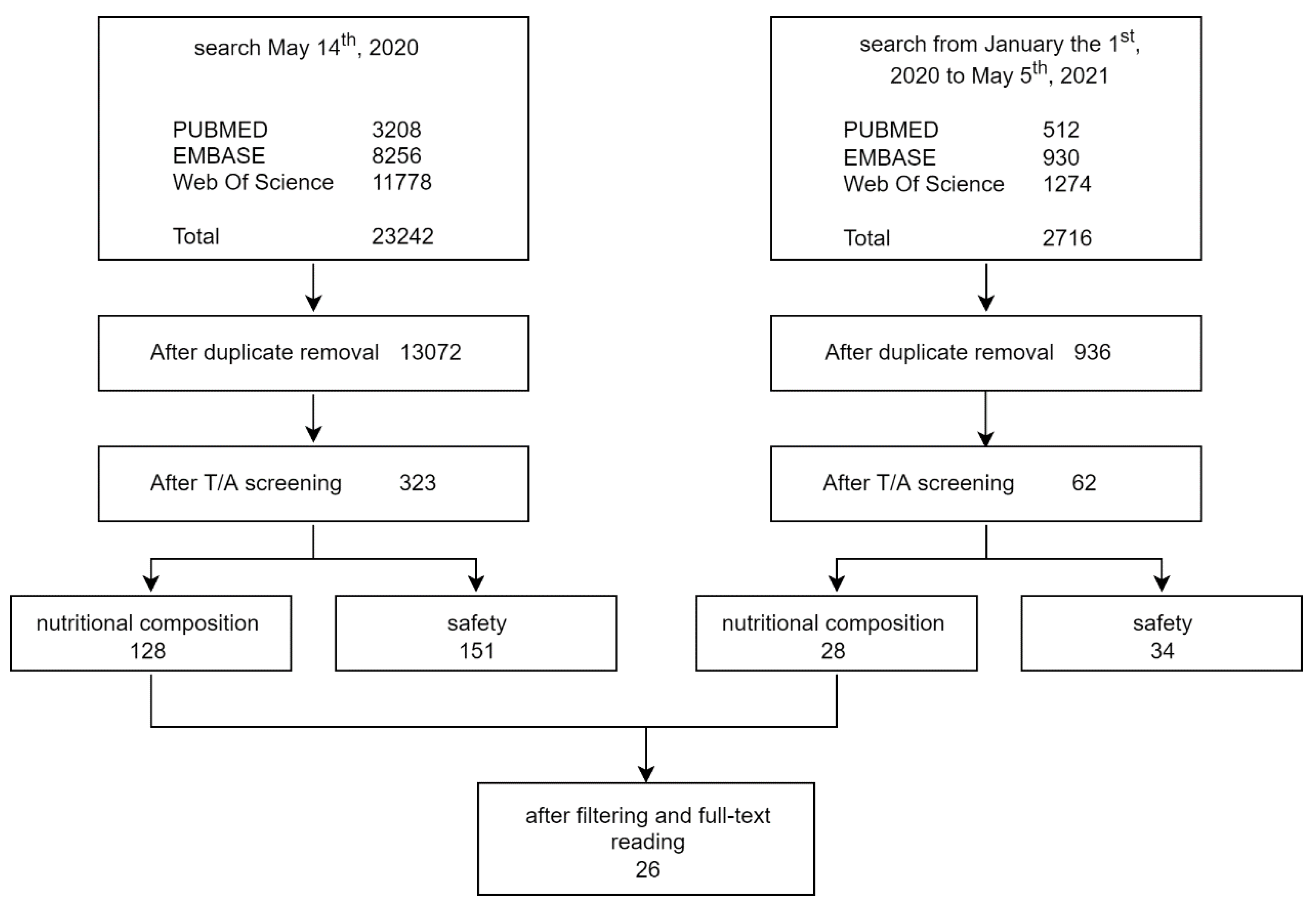
2. Materials and Methods
- Research involving pupae as feed;
- Research describing the composition of the larval stage;
- Research published before the year 2000.
3. Results
3.1. Macronutrients and Ash

{kind=link}
| Reference | Treatment, Strain, Diet | Separation | Protein (Kp = 6.25) | Protein (Kp = 4.76) | Fat | Crude Fiber | NFE (Often Indicates as Carbohydrates in the Original Paper) | Ash | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mishra et al., 2003 [5] | Cocoons were boiled at 100 °C for 30 min. Pupae were taken out and dissected to remove the intestine and waste material. Composition was expressed on wet weight (100 g). | full-fat | 34.38 ± 0.25 | 26.19 ± 0.19 | 57.64 ± 0.22 | 0.54 ± 0.00 | 5.16 ± 0.19 | 2.27 ± 0.17 | |||
| Akande et al., 2020 [18] | Edible insects boiled without seasonings were prepared with 100 g of each of the edible insects with 150 mL of potable water and boiled for 15 min. Data are referred to as boiled silkworm pupae without seasoning (SWOS). | full-fat | 47.17 ± 1.36 | 35.92 ± 1.04 * | 32.16 ± 0.85 | 1.68 ± 0.02 | 28.15 ± 1.23 | 2.12 ± 0.03 | |||
| David-Birman et al., 2019 [19] | Finely milled silkworm pupae flour (SWF) was purchased from JR Unique Foods Ltd. (Udon Thani, Thailand). | full-fat | 53.05 ± 0.09 | 40.4 ± 0.07 * | 29.56 ± 0.08 | n/a | n/a | n/a | |||
| Kim et al., 2016 [20] | Dried, untreated SP flour. The whole insects were cleaned with distilled water, sieved, ground using a hammer mill, and passed through a 20-mesh sieve. | full-fat | 47.87 ± 1.72 | 36.46 ± 1.31 | 30.85 ± 3.81 | 6.38 ± 0.32 ** | n/a | 6.96 ± 0.61 | |||
| The SP flour was then defatted using hexane | defatted | 67.54 ± 0.97 | 51.44 ± 0.74 | 4.75 ± 1.79 | 9.54 ± 0.58 ** | n/a | 9.54 ± 1.22 | ||||
| SP defatted using hexane. The insect flour was also acid-hydrolyzed and dried in the dry oven at 70 °C | defatted | 73.51 ± 0.74 | 55.99 ± 0.56 | 2.05 ± 0.5 | 3.24 ± 0.19 ** | n/a | 8.04 ± 0.18 | ||||
| Anootthato et al., 2019 [21] | Frozen silkworm pupae (−20 °C) were thawed at 4 °C overnight and washed three times before blanching at 95 °C for 10 min. | full-fat | 56.64 ± 0.42 | 43.14 ± 0.32 | 34.07 ± 0.62 | n/a | n/a | n/a | |||
| The sample was further dried at 60 °C for 18 h. Dried samples were ground and packed in an aluminum foil bag until use | full-fat | 52.69 ± 0.46 | 40.13 ± 0.35 | 36.21 ± 0.23 | n/a | n/a | n/a | ||||
| The sample was further dried at 60 °C for 18 h, ground, and packed in an aluminum foil bag until use. Dried samples were mixed and defatted using 99.8% ethanol (1:3 w/v) at 50 °C with stirring for 30 min. Then, each sample was filtered. Defatting was carried out twice before drying in the tray dryer at 50 °C for 3 h. Defatted samples were ground to 150 μm before hydrolysis | defatted | 82.88 ± 0.5 | 63.12 ± 0.38 | 1.47 ± 0.44 | n/a | n/a | n/a | ||||
| Felix et al., 2020 [22] | The silkworm protein concentrate (SPC) used in this study was supplied by FeedStimulants (Amsterdam, The Netherlands). | full-fat | 55.43 ± 1.10 | 42.22 ± 0.84 | 31.72 ± 0.22 | n/a | 5.60 ± 1.42 | 7.24 ± 0.88 | |||
| SPC was defatted using n-hexane | defatted | 81.14 ± 2.06 | 61.80 ± 1.57 | <0.2 | n/a | 8.23 ± 2.04 | 10.63 ± 1.27 | ||||
| Akande et al., 2020 [23] | Freshly harvested mulberry silkworm pupae (SWP) were stifled in an oven at 93 °C for 1 h, cut out of their cocoons and dried in a hot air oven (Gen. Lab. oven) at 40 °C for 8 h. The dried SWP was milled into flour using an electric blender (Binatone, Model No. 51-777) and kept airtight. The data are referred to as silkworm pupae powder (SWP). | full-fat | 60.7 ± 0.2 | 46.23 ± 0.15 | 23.5 ± 0.21 | 1.1 ± 0.16 | 11.3 ± 0.42 | 0.9 ± 0.56 | |||
| Kim et al., 2016 [24] | The dried edible insects were purchased from three vendors located in Chungnam, Korea. Nutrients were originally expressed as g/kg dry weight. | full-fat | 52.58 ± 0.36 | 40.04 ± 0.27 | 19.21 ± 0.34 | n/a | 23.41 ± 0.01 | 4.8 ± 0.71 | |||
| Kuntadi et al., 2018 [25] | The SP were oven dried at 60–70 °C for 12–24 h and then ground. | full-fat | 60.03 | 45.72 | 29.47 | n/a | 0.92 | 5.79 | |||
| Lamberti et al., 2019 [26] | SP were collected 7 days after reaching the cocoon stage. Pupae were lyophilized and stored at −80 °C. | male silkworm reared on an artificial diet | full-fat | 61.45 | 46.80 | n/a | n/a | n/a | 5.68 | ||
| male silkworm reared on leaf | full-fat | 55.02 | 41.90 | n/a | n/a | n/a | 4.62 | ||||
| female silkworm reared on an artificial diet | full-fat | 72.36 | 55.11 | n/a | n/a | n/a | 5.46 | ||||
| female silkworm reared on leaf | full-fat | 56.29 | 42.87 | n/a | n/a | n/a | 5.02 | ||||
| Rodriguez-Ortega et al., 2016 [27] | Dried pupae. Larvae reared on mulberry tree leaves. | full-fat | 64.31 ± 0.00 (true protein = 46.87 ± 0.00) | 48.98 ± 0.00 | 20.63 ± 0.105 | 4.89 ± 0.107 | 3.92 | 6.25 ± 0.017 | |||
| Tomotake et al., 2010 [28] | SP were purchased from Nishiki Food Ltd. (Nagano, Japan). SP were lyophilized followed by grinding into a fine powder. | full-fat | 55.60 | 42.34.00 | 32.20 | n/a | n/a | n/a | |||
| Pereira et al., 2003 [29] | Silkworm pupae fed with younger leaves of mulberry tree, and dried at 130 °C, until 4–5% of moisture. | full-fat | 51.1 ± 1.8 | 38.92 ± 1.37 | 34.4 ± 0.8 | n/a | n/a | 3.64 ± 0.09 | |||
| Ghosh et al., 2020 [30] | Pupae of B. mori were ground in a mortar and pestle and centrifuged adding some distilled water. Then the liquid centrifugate was spray-dried and ground into fine powdered material. | full-fat | 55.87 ± 1.16 | 42.55 ± 0.88 | 23.45 ± 0.28 | 3.77 ± 1.16 | 1.76 ± 1.19 | 5.38 ± 0.07 | |||
| Mishyna et al., 2020 [31] | Insects were stored at −20 °C prior to grinding or drying. After drying, the insects were ground in a food blender and stored in a double plastic bag at −20 °C prior to use, but no longer than 1 month. | freeze-dried | full-fat | 60.29 ± 0.38 | 45.91 ± 0.29 | n/a | n/a | n/a | n/a | ||
| oven-dried | full-fat | 59.33 ± 0.12 | 45.19 ± 0.09 | n/a | n/a | n/a | n/a | ||||
| microwave-dried | full-fat | 61.57 ± 0.21 | 46.89 ± 0.16 | n/a | n/a | n/a | n/a | ||||
| Hirunyophat et al., 2021 [32] | Frozen and stored at −18 °C. | Nangnoi (NN) strain | full-fat | 54.83 ± 0.43 | 41.76 ± 0.33 | 23.61 ± 0.56 | 2.67 ± 0.06 | 14.99 ± 1.05 | 3.99 ± 0.11 | ||
| Siwtui (ST) strain | full-fat | 55.39 ± 0.19 | 42.19 ± 0.14 | 21.26 ± 0.92 | 2.91 ± 0.92 | 17.45 ± 0.83 | 3.12 ± 0.06 | ||||
| Luang Saraburi (LS) strain | full-fat | 50.36 ± 0.21 | 38.35 ± 0.16 | 27.59 ± 0.43 | 3.29 ± 0.07 | 13.42 ± 0.43 | 5.41 ± 0.09 | ||||
| Ubon Ratchathani 60–35 (U 60–35) strain | full-fat | 48.31 ± 0.55 | 36.79 ± 0.42 | 29.93 ± 0.85 | 2.81 ± 0.03 | 14.87 ± 0.08 | 4.23 ± 0.47 | ||||
| Frozen and stored at −18 °C, thawed at 5 °C for 12 h, and dried at 60 °C for 16 h; ground to pass through 177-micron mesh sieve; stored at −18 °C; composition dry weight. | Nangnoi (NN) strain | full-fat | 52.94 ± 0.11 | 40.32 ± 0.08 | 25.53 ± 0.01 | 4.78 ± 0.06 | 12.00 ± 0.09 | 4.69 ± 0.05 | |||
| Siwtui (ST) strain | full-fat | 56.00 ± 0.4 | 42.65 ± 0.3 | 23.95 ± 0.15 | 5.58 ± 0.09 | 10.78 ± 0.7 | 3.94 ± 0.02 | ||||
| Luang Saraburi (LS) strain | full-fat | 49.19 ± 0.26 | 37.46 ± 0.2 | 30.47 ± 0.33 | 5.46 ± 0.14 | 9.93 ± 0.76 | 4.73 ± 0.06 | ||||
| Ubon Ratchathani 60–35 (U 60–35) strain | full-fat | 46.22 ± 0.15 | 35.2 ± 0.11 | 34.69 ± 0.27 | 4.92 ± 0.19 | 9.82 ± 0.03 | 4.25 ± 0.02 | ||||
3.2. Amino Acid Profile
3.3. Fatty Acid Profile
| Reference | Khoeler et al., 2019 [17] | Akande et al., 2020 [18] | Anootthato et al., 2019 [21] | Tomotake et al., 2010 [28] | Kwon et al., 2012 [36] | Shi et al., 2018 [37] | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatment | SH (ready-to-eat, deep-fried in cooking oil. Samples were frozen and freeze-dried for 24 h, powdered and conserved at −80 °C) | SM (non-fried snack, cooked using steam and hot air. Samples were frozen and freeze-dried for 24 h, powdered and conserved at −80 °C) | SP boiled without seasonings, blended | Frozen SP were thawed at 4 °C overnight and washed three times before blanching at 95 °C for 10 min. Then, the sample was dried in a tray dryer at 60 °C for 18 h | SP were lyophilized, followed by grinding into a powder | Frozen SP were extracted and then the protein extract was hydrolyzed with hydrochloric acid at 110 °C for 24 h | SP protein hydrolysate, obtained through enzymatic hydrolysis | |||||
| Pupal age: Day 12–13 | Pupal age: Day D7 | Pupal age: Day D10 | Pupal age: Day D13 | Spray dried | Freeze dried | |||||||
| Amino acid | Val | 5.95 | 5.99 | 4.94 ± 0.04 | 6.18 ± 0.12 | 5.53 | 5.7 | 4.2 | 4 | 2.8 | 5.27 | 4.86 |
| Thr | 4.82 | 4.72 | 4.62 ± 0.06 | 5.08 ± 0.07 | 4.59 | 3.3 | 4.6 | 8.5 | 9.7 | 3.89 | 5.19 | |
| Ile | 4.16 | 4.35 | 4.92 ± 0.09 | 4.84 ± 0.10 | 4 | 2.7 | 2.3 | 1.8 | 2.1 | 4.27 | 2.61 | |
| Leu | 7.44 | 7.39 | 8.16 ± 0.07 | 8.16 ± 0.07 | 7.29 | 3.5 | 2.4 | 2.1 | 2.5 | 7.07 | 6.04 | |
| Lys | 7.4 | 7.19 | 9.00 ± 0.05 | 9.09 ± 0.21 | 7.18 | 4.5 | 2.9 | 3.1 | 3.9 | 7.78 | 7.93 | |
| Met | 3.37 | 4.01 | 2.28 ± 0.22 | - | 4 | 5 | 6.5 | 6.4 | 5 | 2.42 | 2.3 | |
| Phe | 5.04 | 5.71 | 4.21 ± 0.07 | 5.25 ± 0.02 | 5.41 | 3 | 2.6 | 1.4 | 1.6 | 7.2 | 7.33 | |
| His | 2.98 | 3.04 | 2.63 ± 0.06 | 3.25 ± 0.02 | 3.18 | 14.6 | 25.1 | 16 | 13.8 | 3.16 | 3.66 | |
| Trp | 1.49 | 1.55 | 1.28 ± 0.06 | - | 1.76 | - | - | - | - | - | - | |
| Glu | 10.99 | 11.31 | 17.02 ± 0.46 | 13.58 ± 0.21 | 11.18 | 18.3 | 18.9 | 21.5 | 18.1 | 12.44 | 12.68 | |
| Gln | - | - | - | - | - | - | - | - | - | - | - | |
| Gly | 4.6 | 4.18 | 5.16 ± 0.04 | 4.51 ± 0.05 | 4.24 | 7.3 | 5.2 | 7 | 6.1 | 8.73 | 10.09 | |
| Ala | 5.47 | 4.85 | 7.85 ± 0.04 | 5.80 ± 0.07 | 4.59 | 10.2 | 6.4 | 3.8 | 5.1 | 5.14 | 4.77 | |
| Ser | 4.95 | 4.61 | 4.63 ± 0.09 | 5.30 ± 0.10 | 4.35 | 5.2 | 3.4 | 9 | 10.6 | 3.78 | 3.1 | |
| Pro | 4.55 | 4.09 | 3.61 ± 0.05 | 4.87 ± 0.10 | 8.24 | 4.1 | 6.1 | 4.4 | 3.4 | 9.58 | 9.76 | |
| Asp | 11.69 | 11.48 | 8.57 ± 0.04 | - | 10.71 | 1.5 | 1.5 | 2.2 | 6 | 11.23 | 11.04 | |
| Asn | - | - | - | 12.12 ± 0.31 | - | - | - | - | - | - | - | |
| Arg | 5.47 | 5.77 | 6.62 ± 0.06 | 6.61 ± 0.10 | 5.53 | 3.6 | 1.3 | 1.5 | 2.5 | 3.25 | 3.68 | |
| Tyr | 8.23 | 8.46 | 2.62 ± 0.41 | 5.35 ± 0.14 | 6.59 | 6.2 | 6.2 | 6.8 | 6.7 | 3.79 | 4.09 | |
| Cys | 1.4 | 1.31 | 1.90 ± 0.05 | - | 1.65 | 1.5 | 0.2 | 0.6 | 0.4 | 1 | 0.88 | |
| Reference | Treatment | C12:0 | C14:0 | C15:0 | C15:1 | C16:0 | C16:1, C16:1n-7 | C16:2n-6 | C16:3n-3 | C17:0 | C17:1 | C18:0 | C18:1, C18:1n-7, C18:1n-9 | C18:2, C18:2(n-6) | C18:3, C18:3(n-3) | C20:0 | C20:1, C20:1n-9 | C20:3 (and C20:3n-6, C20:3n-3) | C20:4n-6 | C20:5, C20:5n-3 | C22:0 | C24:0 | others | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hu et al., 2017 [4] | BMP were dried in a vacuum drier at 70 °C for 24h to a moisture content of <5% and ground into a fine powder. The powder was sieved and stored at 4 °C. | Microwave assisted extraction, using a mixed solvent consisting of ethanol and n-hexane (1:1, v/v) | - | 0.18 ± 0 | - | - | 23.18 ± 0.52 | 1.07 ± 0.09 | - | - | 0.15 ± 0 | 0.1 ± 0 | 4.69 ± 0.17 | 28.32 ± 0.63 | 3.88 ± 0.13 | 38.25 ± 0.75 | 0.16 | - | - | - | - | - | - | - |
| Soxhlet extraction using n-hexane, at 80 °C for 6 h. The oil was dried at 100 °C ± 5 °C for 15 min. in a drying oven | - | 0.19 ± 0 | - | - | 23.04 ± 0.58 | 1.05 ± 0.07 | - | - | 0.17 ± 0 | nd ± nd | 4.68 ± 0.19 | 28.15 ± 0.54 | 3.85 ± 0.54 | 38.06 ± 0.68 | 0.16 | - | - | - | - | - | - | - | ||
| Tomotake et al., 2010 [28] | Silkworm pupae were lyophilized, followed by grinding into a powder. Lipid content was determined according to the methods of AOAC. | - | 0.1 | - | - | 24.2 | 1.7 | - | - | - | - | 4.5 | 26 | 7.3 | 36.3 | - | - | - | - | - | - | - | - | |
| Pereira et al., 2003 [29] | Chrysalis (worm) toast (originated from the silkworm pupae fed with younger mulberry tree leaves and dried at 130 °C until 4–5% of moisture). Lipids were chemically extracted using chloroform-methanol. | - | 0.164 ± 0.02 | - | - | 24.6 ± 2.1 | 0.656 ± 0.04 | - | - | 0.192 ± 0.06 | - | 7.56 ± 1.54 | 34.8 ± 3.3 | 7.03 ± 1.08 | 24.4 ± 6.7 | - | - | 0.275 ± 0.1 | 0.334 ± 0.08 | - | - | - | - | |
| Wei et al., 2009 [33] | Silkworm pupae were vacuum dried at 60 °C to a moisture content of <5%, then finely ground to powder, and finally oil was extracted by supercritical CO2 extraction. | - | - | - | - | 21.77 | - | - | - | - | - | 7.02 | 33.26 | 7.12 | 27.99 | - | - | - | - | - | - | - | 2.84 | |
| Kwon et al., 2012 [35] | The frozen silkworm pupae were ground, blended, mixed with ethanol and stirred. The sample was filtered, and the particles were extracted with ethyl acetate. The extracts were vacuum-concentrated and dissolved in hexane. Activated carbon was added, and the mixture was heated for 1 h and then cooled and filtered before vacuum-concentration. | Pupal period days 12–13 | - | - | - | - | 19.7 | 2.5 | - | - | - | - | 8.6 | 19.9 | 7.4 | 41.6 | - | - | - | - | 0.3 | - | - | - |
| Pupae at day 7 | - | - | - | - | 22.6 | 2 | - | - | - | - | 9.6 | 21.1 | 8.2 | 36.4 | - | - | - | - | - | - | - | - | ||
| Pupae at day 10 | - | - | - | - | 21 | 1.7 | - | - | - | - | 8.3 | 24.3 | 5.7 | 38.9 | - | - | - | - | 0.1 | - | - | - | ||
| Pupae at day 13 | - | - | - | - | 20 | 1.8 | - | - | - | - | 8.8 | 21.8 | 8.7 | 38.6 | - | - | - | - | 0.3 | - | - | - | ||
| Chieco et al., 2019 [37] | Cocoons were dried for 3 days at 60 °C. The pupae were extracted and stored at −80 °C. Frozen pupae were milled to a fine powder. Lipids were chemically extracted using a chloroform/methanol solution according to the Folch’s method. | White polyhibrid on artificial diet | - | - | - | - | 29.2 ± 0.4 | 1.5 ± 0 | - | - | - | - | 10.9 ± 0.1 | 35.1 ± 0.3 | 11.4 ± 0.5 | 11.7 ± 0.3 | - | - | - | - | - | - | - | - |
| Golden nistari on artificial diet | - | - | - | - | 25.1 ± 0.3 | 1.5 ± 0 | - | - | - | - | 9.3 ± 0.2 | 39.8 ± 0.3 | 11.8 ± 0.1 | 12.3 ± 0.2 | - | - | - | - | - | - | - | - | ||
| White polyhibrid on mulberry leaf of the Florio cultivar | - | - | - | - | 25.2 ± 0.7 | 0.8 ± 0.1 | - | - | - | - | 7.2 ± 0.2 | 35.2 ± 0.3 | 7.1 ± 0.4 | 24.3 ± 0.2 | - | - | - | - | - | - | - | - | ||
| Golden nistari on mulberry leaf of the Florio cultivar | - | - | - | - | 21.6 ± 0.4 | 1.4 ± 0 | - | - | - | - | 4.8 ± 0.1 | 32.3 ± 0.1 | 10.4 ± 0.2 | 29.3 ± 0.2 | - | - | - | - | - | - | - | - | ||
| Tong et al., 2011 [38] | Silkworms were directly frozen on the third day of the 5th instar and then lyophilized. Lipid content was measured using Soxhlet extraction. | - | - | 0.15 | - | 16.03 | 0.39 | - | - | - | - | 9.45 | 28.12 | 12.24 | 31.91 | 0.7 | 0.15 | - | - | - | 0.7 | 0.15 | - | |
| Usub et al., 2008 [39] | Sun-dried pupae. Five grams were grounded and the lipid extracted with 50 mL of chloroform-methanol (2:1, v/v) containing 10 mg L−1 of butylated hydroxytoluene and 0.1 mg L−1 of tricosanoic acid. | 0.1 ± 0 | 0.1 ± 0 | 0.1 ± 0 | 0.1 ± 0 | 17 ± 0.1 | 0.6 ± 0 | - | - | 0.2 ± 0.1 | - | 6.2 ± 0 | 20.4 ± 0.1 | 8.8 ± 0.1 | 45.6 ± 0.2 | 0.4 ± 0 | 0.1 ± 0 | 0.1 ± 0 | 0.1 ± 0 | - | - | - | - | |
| Solar-tunnel dried pupae. Five grams were ground and the lipid extracted with 50 mL of chloroform-methanol (2:1, v/v) containing 10 mg L−1 of butylated hydroxytoluene and 0.1 mg L−1 of tricosanoic acid. | 0.1 ± 0 | 0.1 ± 0 | 0.1 ± 0 | 0.1 ± 0 | 17.5 ± 0.6 | 0.6 ± 0 | - | - | 0.2 ± 0 | - | 6.2 ± 0.1 | 21 ± 0.2 | 8.8 ± 0.1 | 44.6 ± 0.5 | 0.4 ± 0 | 0.1 ± 0 | 0.1 ± 0 | 0.1 ± 0 | - | - | - | - | ||
| Wang et al., 2020 [39] | Silkworm pupae were dried in a vacuum at 60 °C for 24 h to a moisture content of <5% and then slightly ground and sieved through a 60-mesh sieve. Lipid extraction was conducted using the Folch and Sloane method. | - | - | - | - | 22.04 ± 0.49 | 0.92 ± 0.01 | - | - | - | - | 6.84 ± 0.23 | 33.91 ± 0.08 | 5.48 ± 0.14 | 30.81 ± 0.21 | - | - | - | - | - | - | - | - | |
| Yu et al., 2018 [40] | Pupae were frozen in liquid nitrogen and stored at −80 °C before analysis. Pupae were homogenized and oil was chemically extracted using hexane and boron trifluoride-methanol solution. | Three-day old silkworm pupae, strain Dazao, fed with an artificial diet (Silkmate 2S), Table 2 in the original paper | - | - | - | - | 16.6 ± 2.38 | 0.51 ± 0.04 | 0.55 ± 0 | 0.38 ± 0 | - | - | 11.26 ± 0.7 | 28.53 ± 1.83 | 20.54 ± 1.68 | 21.13 ± 1.7 | 0.51 ± 0.12 | - | - | - | - | - | - | - |
| Three-day-old silkworm pupae, strain Dazao, fed with an artificial diet (Silkmate 2S), Table 3 in the original paper | - | - | - | - | 15.61 ± 1.89 | 0.46 ± 0.09 | 0.46 ± 0.1 | 0.23 ± 0.17 | - | - | 12.87 ± 1.4 | 26.14 ± 1.41 | 22.24 ± 1.25 | 21.31 ± 1.21 | 0.67 ± 0.2 | - | - | - | - | - | - | - | ||
| Three-day-old silkworm pupae, strain Dazao, female | - | - | - | - | 17.37 ± 0.53 | 0.53 ± 0.06 | 0.48 ± 0.06 | 0.33 ± 0.05 | - | - | 12.55 ± 1.32 | 26.63 ± 1.4 | 21.32 ± 0.53 | 20.26 ± 0.59 | 0.53 ± 0.19 | - | - | - | - | - | - | - | ||
| Three-day-old silkworm pupae, strain Dazao, male | - | - | - | - | 14.34 ± 1.19 | 0.45 ± 0.08 | 0.55 ± 0.06 | 0.36 ± 0.05 | - | - | 12.54 ± 1.46 | 25.38 ± 1.72 | 23.11 ± 1.09 | 22.57 ± 0.92 | 0.71 ± 0.11 | - | - | - | - | - | - | - | ||
| Three-day-old silkworm pupae, strain 305, female | - | - | - | - | 18.44 ± 1.59 | 1.19 ± 0.57 | 0.53 ± 0.03 | 0.29 ± 0.15 | - | - | 12.93 ± 2.14 | 28.23 ± 2.08 | 18.72 ± 1.05 | 19.14 ± 1.3 | 0.53 ± 0.18 | - | - | - | - | - | - | - | ||
| Three-day-old silkworm pupae, strain 305, male | - | - | - | - | 15.23 ± 0.36 | 1.16 ± 0.58 | 0.57 ± 0.08 | 0.38 ± 0.06 | - | - | 13.12 ± 0.96 | 25.97 ± 1.57 | 21.2 ± 1.3 | 21.54 ± 0.64 | 0.83 ± 0.38 | - | - | - | - | - | - | - | ||
3.4. Mineral Content
| Ref. | Köhler et al., 2019 [17] | Akande et al., 2020 [18] | David-Birman et al., 2019 [19] | Kim et al., 2016 [24] | Kuntadi et al., 2018 [25] | Rodriguez-Ortega et al., 2016 [27] | Pereira et al., 2003 [29] | ||
|---|---|---|---|---|---|---|---|---|---|
| mg/100 g, (Edible Portion) | M ± SD; mg/100 g | M ± SD; mg/100 g (dry weight) | M ± SD; mg/100 g (dry weight) | mg/100 g (dry weight) | Mean ± SD; mg/100 g (dry weight) | M ± SD; mg/100 g (dry weight) | |||
| Street Hawker | Supermarket | ||||||||
| Minerals | Cr | - | - | - | N/D | - | - | - | 0.253 ± 0.023 |
| Hg | <0.005 | <0.005 | - | N/D | - | - | - | - | |
| Pb | 0.0138 | 0.0044 | - | N/D | - | - | - | - | |
| Cd | <0.005 | <0.005 | - | N/D | - | - | - | - | |
| As | 0.0432 | 0.0165 | - | - | - | - | - | - | |
| Se | 0.0534 | 0.2285 | - | - | - | - | - | - | |
| Mo | 0.0151 | 0.0516 | - | - | 0.02 ± 0.00 | - | - | - | |
| B | - | - | - | - | 1.46 ± 0.42 | - | - | - | |
| Cu | 0.711 | 0.943 | - | - | 0.94 ± 0.01 | - | - | 1.52 ± 0.07 | |
| Na | 128.4 | 362.6 | - | 41.86 ± 3.45 | - | - | - | 29.60 ± 2.10 | |
| K | 492.9 | 672 | - | 513.8 ± 1.13 | - | - | - | 477 ± 16 | |
| Mg | 157.7 | 174.7 | - | 280.8 ± 1.13 | 252.29 ± 10.26 | - | - | 89 ± 0 | |
| Fe | 2.83 | 3.18 | 0.142 ± 0.063 | N/D | 4.95 ± 0.05 | 3.54 | - | 3.00 ± 0.88 | |
| Zn | 9.8 | 15.4 | 0.203 ± 0.004 | 1.39 ± 0.07 | 14.70 ± 1.45 | - | - | 24.40 ± 0.96 | |
| Mn | 1.26 | 1.86 | 0.021 ± 0.021 | 1.08 ± 0.05 | 1.68 ± 0.06 | - | - | 2.30 ± 0.17 | |
| Ca | 92.2 | 107.6 | 0.249 ± 0.013 | 137.01 ± 1.34 | 98.72 ± 0.00 | 29.17 | 760 ± 0 | 181 ± 9 | |
| P | 492.9 | 672 | 5.57 ± 0.144 | - | 870.92 ± 15.77 | - | 720 ± 0 | - | |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, S.-Y.; Han, M.-J.; Kang, L.-F.; Li, Z.-W.; Shen, Y.-H.; Zhang, Z. Demographic History and Gene Flow during Silkworm Domestication. BMC Evol. Biol. 2014, 14, 185. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Zhu, Y.; Chen, K. Silkworm: A Promising Model Organism in Life Science. J. Insect Sci. 2017, 17, 97. [Google Scholar] [CrossRef]
- Xu, S.; Wang, F.; Wang, Y.; Wang, R.; Hou, K.; Tian, C.; Ji, Y.; Yang, Q.; Zhao, P.; Xia, Q. A Silkworm Based Silk Gland Bioreactor for High-Efficiency Production of Recombinant Human Lactoferrin with Antibacterial and Anti-Inflammatory Activities. J. Biol. Eng. 2019, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Li, C.; Zhang, Z.; Zhao, Q.; Zhu, Y.; Su, Z.; Chen, Y. Microwave-Assisted Extraction of Silkworm Pupal Oil and Evaluation of Its Fatty Acid Composition, Physicochemical Properties and Antioxidant Activities. Food Chem. 2017, 231, 348–355. [Google Scholar] [CrossRef]
- Mishra, N.; Hazarika, N.C.; Narain, K.; Mahanta, J. Nutritive Value of Non-Mulberry and Mulberry Silkworm Pupae and Consumption Pattern in Assam, India. Nutr. Res. 2003, 23, 1303–1311. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, Z. Study on Edible Insect Resources and Their Exploitation and Utilization. Resour. Sci. 2001, 23, 58–61. [Google Scholar]
- Mitsuhashi, J. Insects as Traditional Foods in Japan. Ecol. Food Nutr. 1997, 36, 187–199. [Google Scholar] [CrossRef]
- Han, R.; Shin, J.T.; Kim, J.; Choi, Y.S.; Kim, Y.W. An Overview of the South Korean Edible Insect Food Industry: Challenges and Future Pricing/Promotion Strategies: S. Korean Edible Insect Food Industry. Entomol. Res. 2017, 47, 141–151. [Google Scholar] [CrossRef]
- Yhoung-Aree, J.; Puwastien, P.; Attig, G.A. Edible Insects in Thailand: An Unconventional Protein Source? Ecol. Food Nutr. 1997, 36, 133–149. [Google Scholar] [CrossRef]
- Aznar-Cervantes, S.D.; Monteagudo Santesteban, B.; Cenis, J.L. Products of Sericulture and Their Hypoglycemic Action Evaluated by Using the Silkworm, Bombyx mori (Lepidoptera: Bombycidae), as a Model. Insects 2021, 12, 1059. [Google Scholar] [CrossRef]
- Ratcliffe, N.A.; Mello, C.B.; Garcia, E.S.; Butt, T.M.; Azambuja, P. Insect Natural Products and Processes: New Treatments for Human Disease. Insect Biochem. Mol. Biol. 2011, 41, 747–769. [Google Scholar] [CrossRef] [PubMed]
- Zimian, D.; Yonghua, Z.; Xiwu, G. Medicinal Insects in China. Ecol. Food Nutr. 1997, 36, 209–220. [Google Scholar] [CrossRef]
- EFSA Scientific Committee. Risk Profile Related to Production and Consumption of Insects as Food and Feed. EFSA J. 2015, 13, 4257. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; He, K.; Velickovic, T.C.; Liu, Z. Nutritional, Functional, and Allergenic Properties of Silkworm Pupae. Food Sci. Nutr. 2021, 9, 4655–4665. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.; Brunton, J.; Graziosi, S. EPPI-Reviewer 4.0: Software for Research Synthesis. EPPI-Centre Software; Social Science Research Unit, Institute of Education, University of London: London, UK, 2010. [Google Scholar]
- Janssen, R.H.; Vincken, J.-P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio Molitor, Alphitobius Diaperinus, and Hermetia Illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Köhler, R.; Kariuki, L.; Lambert, C.; Biesalski, H.K. Protein, Amino Acid and Mineral Composition of Some Edible Insects from Thailand. J. Asia-Pac. Entomol. 2019, 22, 372–378. [Google Scholar] [CrossRef]
- Akande, O.A.; Falade, O.O.; Badejo, A.A.; Adekoya, I. Assessment of Mulberry Silkworm Pupae and African Palm Weevil Larvae as Alternative Protein Sources in Snack Fillings. Heliyon 2020, 6, e03754. [Google Scholar] [CrossRef]
- David-Birman, T.; Moshe, H.; Lesmes, U. Impact of Thermal Processing on Physicochemical Properties of Silk Moth Pupae (Bombyx mori) Flour and in-Vitro Gastrointestinal Proteolysis in Adults and Seniors. Food Res. Int. 2019, 123, 11–19. [Google Scholar] [CrossRef]
- Kim, H.-W.; Setyabrata, D.; Lee, Y.J.; Jones, O.G.; Kim, Y.H.B. Pre-Treated Mealworm Larvae and Silkworm Pupae as a Novel Protein Ingredient in Emulsion Sausages. Innov. Food Sci. Emerg. Technol. 2016, 38, 116–123. [Google Scholar] [CrossRef]
- Anootthato, S.; Therdthai, N.; Ritthiruangdej, P. Characterization of Protein Hydrolysate from Silkworm Pupae (Bombyx mori). J. Food Process. Preserv. 2019, 43. [Google Scholar] [CrossRef]
- Felix, M.; Bascon, C.; Cermeño, M.; FitzGerald, R.J.; de la Fuente, J.; Carrera-Sánchez, C. Interfacial/Foaming Properties and Antioxidant Activity of a Silkworm (Bombyx mori) Pupae Protein Concentrate. Food Hydrocoll. 2020, 103, 105645. [Google Scholar] [CrossRef]
- Akande, A.O.; Jolayemi, O.S.; Adelugba, V.A.; Akande, S.T. Silkworm Pupae (Bombyx mori) and Locusts as Alternative Protein Sources for High-Energy Biscuits. J. Asia-Pac. Entomol. 2020, 23, 234–241. [Google Scholar] [CrossRef]
- Kim, S.-K.; Weaver, C.M.; Choi, M.-K. Proximate Composition and Mineral Content of Five Edible Insects Consumed in Korea. CyTA–J. Food 2016, 15, 143–146. [Google Scholar] [CrossRef]
- Kuntadi, K.; Adalina, Y.; Maharani, K.E. Nutritional Composition of Six Edible Insects in Java. Indones. J. For. Res. 2018, 5, 57–68. [Google Scholar] [CrossRef]
- Lamberti, C.; Gai, F.; Cirrincione, S.; Giribaldi, M.; Purrotti, M.; Manfredi, M.; Marengo, E.; Sicuro, B.; Saviane, A.; Cappellozza, S.; et al. Investigation of the Protein Profile of Silkworm (Bombyx mori) Pupae Reared on a Well-Calibrated Artificial Diet Compared to Mulberry Leaf Diet. PeerJ 2019, 7, e6723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez-Ortega, A.; Pino-Moreno, J.M.; Ángeles-Campos, S.C.; García-Pérez, Á.; Barrón-Yánez, R.M.; Callejas-Hernández, J. Valor Nutritivo de Larvas y Pupas de Gusano de Seda (Bombyx mori) (Lepidoptera: Bombycidae). Rev. Colomb. Entomol. 2016, 42, 69. [Google Scholar] [CrossRef]
- Tomotake, H.; Katagiri, M.; Yamato, M. Silkworm Pupae (Bombyx mori) Are New Sources of High Quality Protein and Lipid. J. Nutr. Sci. Vitaminol. 2010, 56, 446–448. [Google Scholar] [CrossRef] [Green Version]
- Pereira, N.R.; Ferrarese-Filho, O.; Matsushita, M.; de Souza, N.E. Proximate Composition and Fatty Acid Profile of Bombyx mori L. Chrysalis Toast. J. Food Compos. Anal. 2003, 16, 451–457. [Google Scholar] [CrossRef]
- Ghosh, A.; Ray, M.; Gangopadhyay, D. Evaluation of Proximate Composition and Antioxidant Properties in Silk-Industrial Byproduct. LWT 2020, 132, 109900. [Google Scholar] [CrossRef]
- Mishyna, M.; Haber, M.; Benjamin, O.; Martinez, J.J.I.; Chen, J. Drying Methods Differentially Alter Volatile Profiles of Edible Locusts and Silkworms. J. Insects Food Feed 2020, 6, 405–415. [Google Scholar] [CrossRef]
- Hirunyophat, P.; Chalermchaiwat, P.; On-nom, N.; Prinyawiwatkul, W. Selected Nutritional Quality and Physicochemical Properties of Silkworm Pupae (Frozen or Powdered) from Two Species. Int. J. Food Sci. Technol. 2021, 56, 3578–3587. [Google Scholar] [CrossRef]
- Wei, Z.-J.; Liao, A.-M.; Zhang, H.-X.; Liu, J.; Jiang, S.-T. Optimization of Supercritical Carbon Dioxide Extraction of Silkworm Pupal Oil Applying the Response Surface Methodology. Bioresour. Technol. 2009, 100, 4214–4219. [Google Scholar] [CrossRef] [PubMed]
- Walker, V.; Mills, G.A. Quantitative Methods for Amino Acid Analysis in Biological Fluids. Ann. Clin. Biochem. Int. J. Lab. Med. 1995, 32, 28–57. [Google Scholar] [CrossRef] [PubMed]
- Kwon, M.-G.; Kim, D.-S.; Lee, J.-H.; Park, S.-W.; Choo, Y.-K.; Han, Y.-S.; Kim, J.-S.; Hwang, K.-A.; Ko, K.; Ko, K. Isolation and Analysis of Natural Compounds from Silkworm Pupae and Effect of Its Extracts on Alcohol Detoxification: Nutritional Value of Silkworm Component. Entomol. Res. 2012, 42, 55–62. [Google Scholar] [CrossRef]
- Chieco, C.; Morrone, L.; Bertazza, G.; Cappellozza, S.; Saviane, A.; Gai, F.; Di Virgilio, N.; Rossi, F. The Effect of Strain and Rearing Medium on the Chemical Composition, Fatty Acid Profile and Carotenoid Content in Silkworm (Bombyx mori) Pupae. Animals 2019, 9, 103. [Google Scholar] [CrossRef] [Green Version]
- Tong, L.; Yu, X.; Liu, H. Insect Food for Astronauts: Gas Exchange in Silkworms Fed on Mulberry and Lettuce and the Nutritional Value of These Insects for Human Consumption during Deep Space Flights. Bull. Entomol. Res. 2011, 101, 613–622. [Google Scholar] [CrossRef]
- Usub, T.; Lertsatitthanakorn, C.; Poomsa-ad, N.; Wiset, L.; Yang, L.; Siriamornpun, S. Experimental Performance of a Solar Tunnel Dryer for Drying Silkworm Pupae. Biosyst. Eng. 2008, 101, 209–216. [Google Scholar] [CrossRef]
- Wang, W.; Xu, L.; Zou, Y.; Pang, D.; Shi, W.; Mu, L.; Li, E.; Lan, D.; Wang, Y.; Liao, S. Comprehensive Identification of Principal Lipid Classes and Tocochromanols in Silkworm (Antheraea pernyi and Bombyx mori) Pupae Oils. Eur. J. Lipid Sci. Technol. 2020, 122, 1900280. [Google Scholar] [CrossRef]
- Yu, X.; Shen, Y.; Cui, Q.; Chen, Y.; Sun, W.; Huang, X.; Zhu, Y. Silkworm (Bombyx mori) Has the Capability to Accumulate C20 and C22 Polyunsaturated Fatty Acids. Eur. J. Lipid Sci. Technol. 2018, 120, 1700268. [Google Scholar] [CrossRef]
- Ghosh, S.; Lee, S.-M.; Jung, C.; Meyer-Rochow, V.B. Nutritional Composition of Five Commercial Edible Insects in South Korea. J. Asia-Pac. Entomol. 2017, 20, 686–694. [Google Scholar] [CrossRef]
- CREA Centro Alimenti e Nutrizione Tabelle Di Composizione Degli Alimenti, Aggiornamento. 2019. Available online: https://www.alimentinutrizione.it/tabelle-nutrizionali (accessed on 15 April 2022).
- World Health Organization; Food and Agriculture Organization of the United Nations. Nitrogen and Protein Content Measurement and Nitrogen to Protein Conversion Factors for Dairy and Soy Protein-Based Foods: A Systematic Review and Modelling Analysis; World Health Organization: Geneva, Switzerland, 2019; ISBN 978-92-4-151698-3. [Google Scholar]
- Cappellozza, L.; Cappellozza, S.; Saviane, A.; Sbrenna, G. Artificial Diet Rearing System for the Silkworm Bombyx mori (Lepidoptera: Bombycidae): Effect of Vitamin C Deprivation on Larval Growth and Cocoon Production. Appl. Entomol. Zool. 2005, 40, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, C.; Zha, X. Midgut and Head Transcriptomic Analysis of Silkworms Reveals the Physiological Effects of Artificial Diets. Insects 2022, 13, 291. [Google Scholar] [CrossRef] [PubMed]
- Horie, Y.; Nakasone, S. Effects of the Levels of Fatty Acids and Carbohydrates in a Diet on the Biosynthesis of Fatty Acids in Larvae of the Silkworm, Bombyx mori. J. Insect Physiol. 1971, 17, 1441–1450. [Google Scholar] [CrossRef]
- Patel, A.; Desai, S.S.; Mane, V.K.; Enman, J.; Rova, U.; Christakopoulos, P.; Matsakas, L. Futuristic Food Fortification with a Balanced Ratio of Dietary ω-3/ω-6 Omega Fatty Acids for the Prevention of Lifestyle Diseases. Trends Food Sci. Technol. 2022, 120, 140–153. [Google Scholar] [CrossRef]
- Simopoulos, A. An Increase in the Omega-6/Omega-3 Fatty Acid Ratio Increases the Risk for Obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Lupette, J.; Benning, C. Human Health Benefits of Very-Long-Chain Polyunsaturated Fatty Acids from Microalgae. Biochimie 2020, 178, 15–25. [Google Scholar] [CrossRef]
- Ibrahim, D.; El-Sayed, R.; Khater, S.I.; Said, E.N.; El-Mandrawy, S.A.M. Changing Dietary N-6:N-3 Ratio Using Different Oil Sources Affects Performance, Behavior, Cytokines MRNA Expression and Meat Fatty Acid Profile of Broiler Chickens. Anim. Nutr. 2018, 4, 44–51. [Google Scholar] [CrossRef]
- Wood, J.D.; Enser, M.; Fisher, A.V.; Nute, G.R.; Sheard, P.R.; Richardson, R.I.; Hughes, S.I.; Whittington, F.M. Fat Deposition, Fatty Acid Composition and Meat Quality: A Review. Meat Sci. 2008, 78, 343–358. [Google Scholar] [CrossRef]
- Trembecka, L.; Hascik, P.; Cubon, J.; Bobko, M.; Pavelkova, A. Fatty Acids Profile of Breast and Thigh Muscles of Broiler Chickens Fed Diets with Propolis and Probiotics. J. Cent. Eur. Agric. 2016, 17, 1179–1193. [Google Scholar] [CrossRef] [Green Version]
- Strobel, C.; Jahreis, G.; Kuhnt, K. Survey of N- 3 and n-6 Polyunsaturated Fatty Acids in Fish and Fish Products. Lipids Health Dis. 2012, 11, 144. [Google Scholar] [CrossRef] [Green Version]
- Zeng, P.; Guo, Z.; Xiao, X.; Peng, C.; Liu, L.; Yan, D.; He, Y. Physiological Stress Responses, Mineral Element Uptake and Phytoremediation Potential of Morus alba L. in Cadmium-Contaminated Soil. Ecotoxicol. Environ. Saf. 2020, 189, 109973. [Google Scholar] [CrossRef] [PubMed]
- Levickienė, D.; Vaitkevičienė, N.; Jarienė, E.; Mažeika, R. The Content of Macroelements in White Mulberry (Morus alba L.) Leaves. Žemės Ūkio Moksl. 2019, 25. [Google Scholar] [CrossRef]
- Saviane, A.; Tassoni, L.; Naviglio, D.; Lupi, D.; Savoldelli, S.; Bianchi, G.; Cortellino, G.; Bondioli, P.; Folegatti, L.; Casartelli, M.; et al. Mechanical Processing of Hermetia Illucens Larvae and Bombyx mori Pupae Produces Oils with Antimicrobial Activity. Animals 2021, 11, 783. [Google Scholar] [CrossRef] [PubMed]

| Keywords (Title/Abstract) | ||||
|---|---|---|---|---|
| Bombyx OR Silkworm OR “silk worms” OR silkmoth OR “silk moths” | AND | nutrition OR composition OR centesimal OR nutrient OR nutrients OR protein OR proteins peptide OR peptides OR aminoacid OR aminoacids OR “amino acid” OR “amino acids” OR acid OR acids OR polypeptide OR polypeptides OR fat OR fats OR lipid OR lipids OR “fatty acid” OR “fatty acids” OR “fatty alcohols” OR sugar OR sugars OR carbohydrate OR carbohydrates OR disaccharide OR disaccharides OR monosaccharide OR monosaccharides OR polysaccharide OR polysaccharides OR ash OR mineral OR minerals OR macronutrient OR macronutrients OR micronutrient OR micronutrients OR oligoelement OR oligoelements OR microelement OR microelements OR vitamin OR vitamins OR oil OR oils OR “trace element” OR “trace elements” | OR | safety OR hazard OR hazards OR risk OR risks OR microorganism OR microorganisms OR pathogen OR pathogens OR contaminant OR contaminants OR contamination OR contaminations OR chemical OR chemicals OR toxic OR toxics OR toxicity OR metal OR metals OR toxin OR toxins OR allergy OR allergies OR allergen OR allergens OR allergic OR allergenic OR sensitization OR sensitisation OR cross-reactivity OR anaphylactic OR anaphylaxis OR poisoning OR poison OR compound OR compounds OR pesticide OR pesticides OR residual OR residue OR residues OR antibiotic OR antibiotics OR antiparasitic OR antiparasitics OR mycotoxin OR mycotoxins OR dioxin OR dioxins OR polluting OR pollutant OR pollutants |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tassoni, L.; Cappellozza, S.; Dalle Zotte, A.; Belluco, S.; Antonelli, P.; Marzoli, F.; Saviane, A. Nutritional Composition of Bombyx mori Pupae: A Systematic Review. Insects 2022, 13, 644. https://doi.org/10.3390/insects13070644
Tassoni L, Cappellozza S, Dalle Zotte A, Belluco S, Antonelli P, Marzoli F, Saviane A. Nutritional Composition of Bombyx mori Pupae: A Systematic Review. Insects. 2022; 13(7):644. https://doi.org/10.3390/insects13070644
Chicago/Turabian StyleTassoni, Luca, Silvia Cappellozza, Antonella Dalle Zotte, Simone Belluco, Pietro Antonelli, Filippo Marzoli, and Alessio Saviane. 2022. "Nutritional Composition of Bombyx mori Pupae: A Systematic Review" Insects 13, no. 7: 644. https://doi.org/10.3390/insects13070644