Reproductive Apparatus, Gonadic Maturation, and Allometry of Cyclocephala barrerai Martínez (Coleoptera: Melolonthidae: Dynastinae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cyclocephala barrerai Insects
2.2. Reproductive Apparatus Description
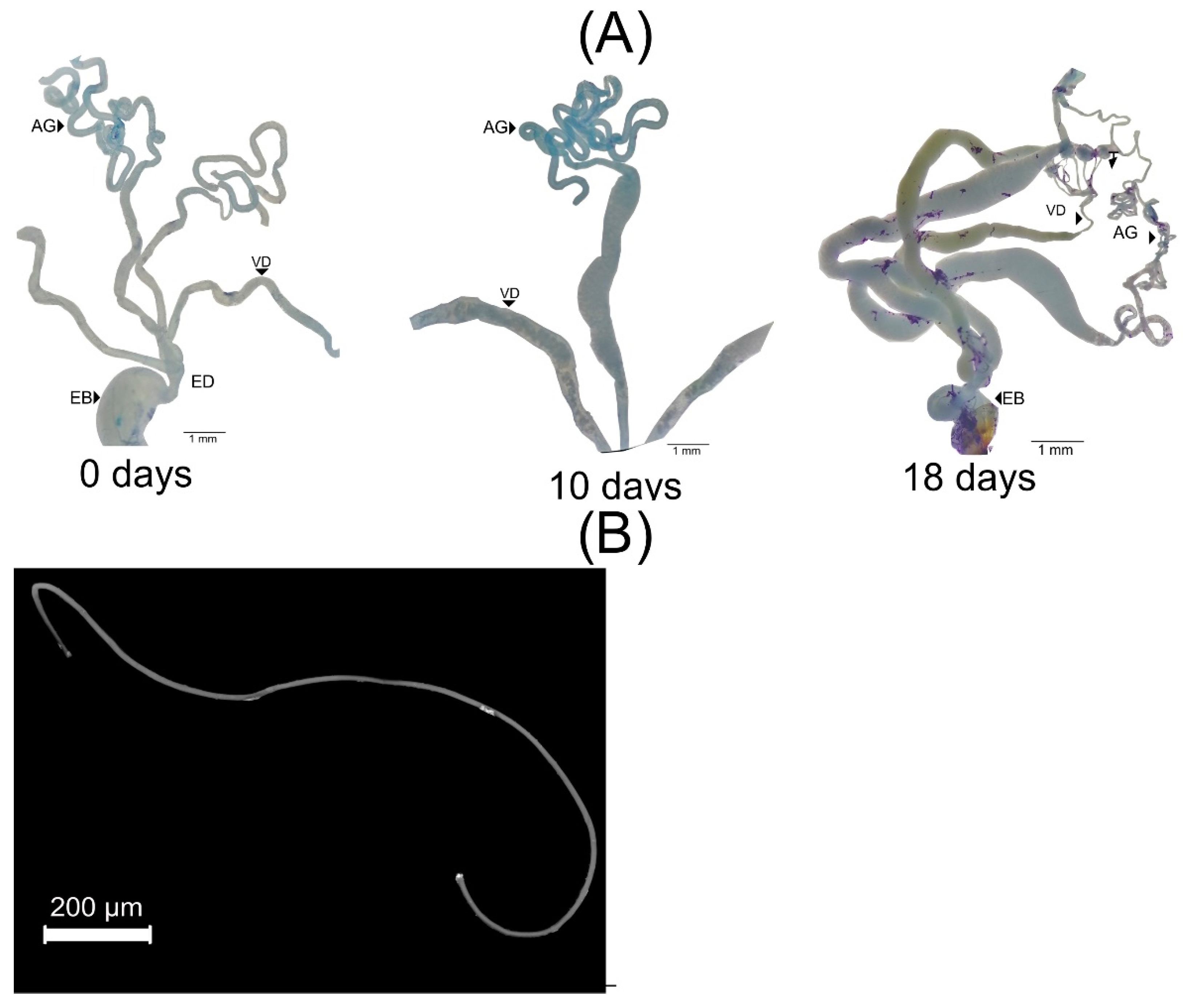
2.3. Gonadic Maturation
2.4. Allometry
Statistical Analysis
3. Results
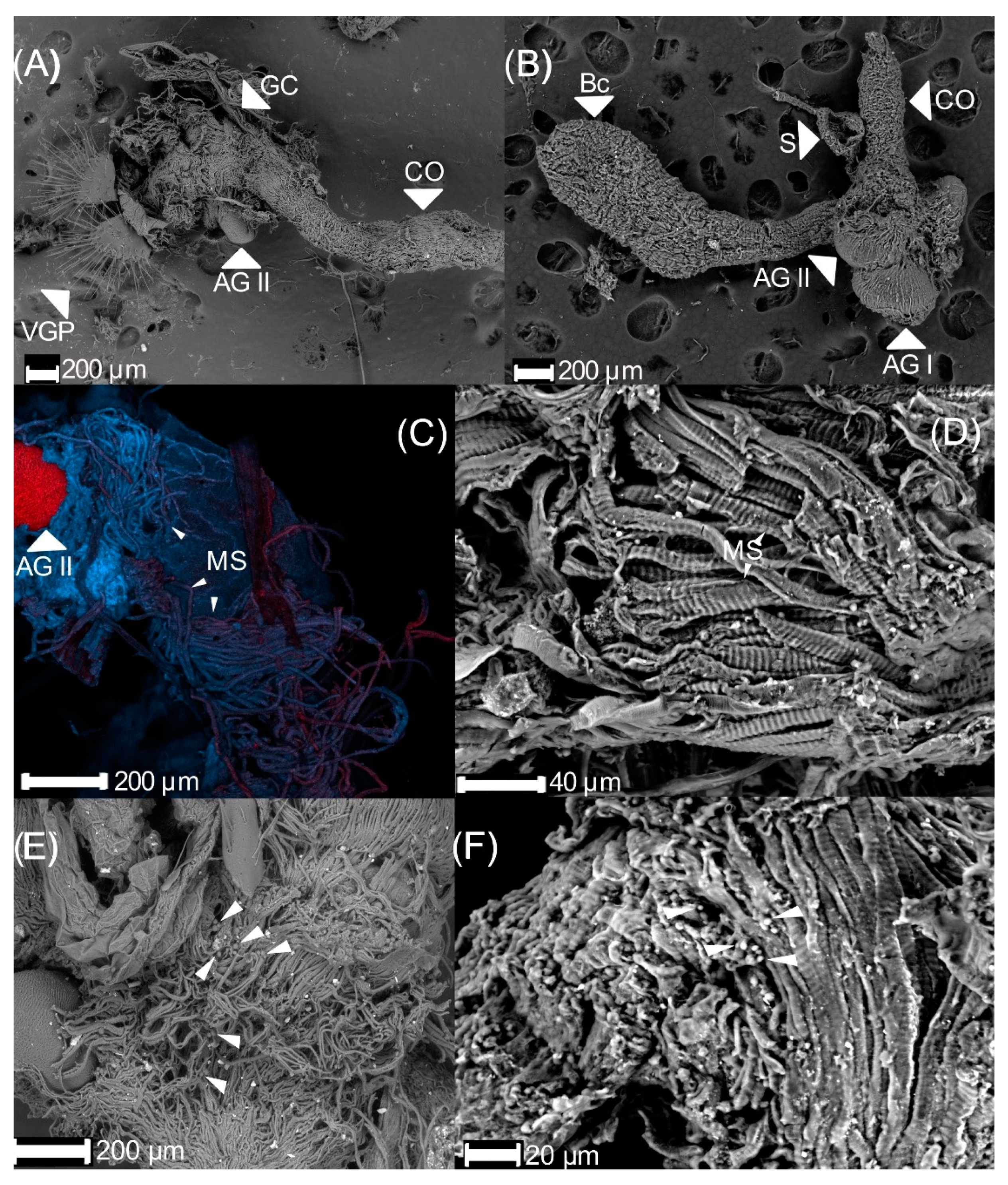
3.1. Male Reproductive Apparatus
- (1)
- The connecting membrane (Figure 3A) is in the ventral zone, connecting the phallobase to parameres.
- (2)
- (3)
- Posterior phallobase and anterior phallobase (Figure 3E). The posterior phallobase is a sub-cylindric structure that involves the aedeagus and connects to the parameres. It is made up of one piece and presents a high diversity of ultrastructure (Figure 3F). The posterior phallobase has four types of ultrastructure: (1) large needle, (2) small needle, (3) pits (Figure 3F), and (4) small seta with a circular base (Figure 3G).
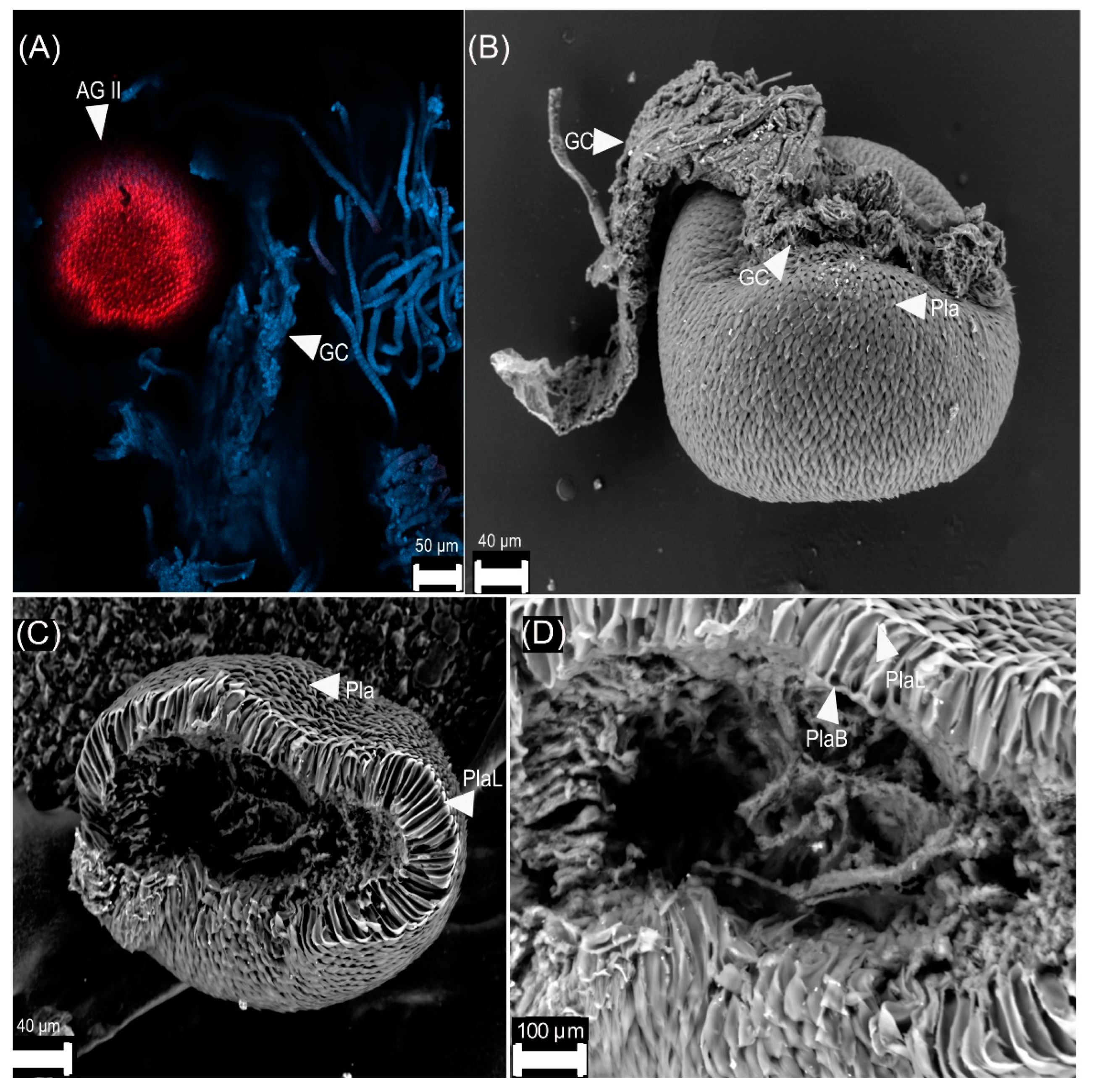
3.2. Female Reproductive Apparatus
- (1)
- Dorsal plates are sclerotized rectangular shape structures. They insert near the accessory glands, around the genital chamber.
- (2)
- (1)
- Type I is oval-shaped, and it inserts in the genital chamber (Figure 5).
- (2)
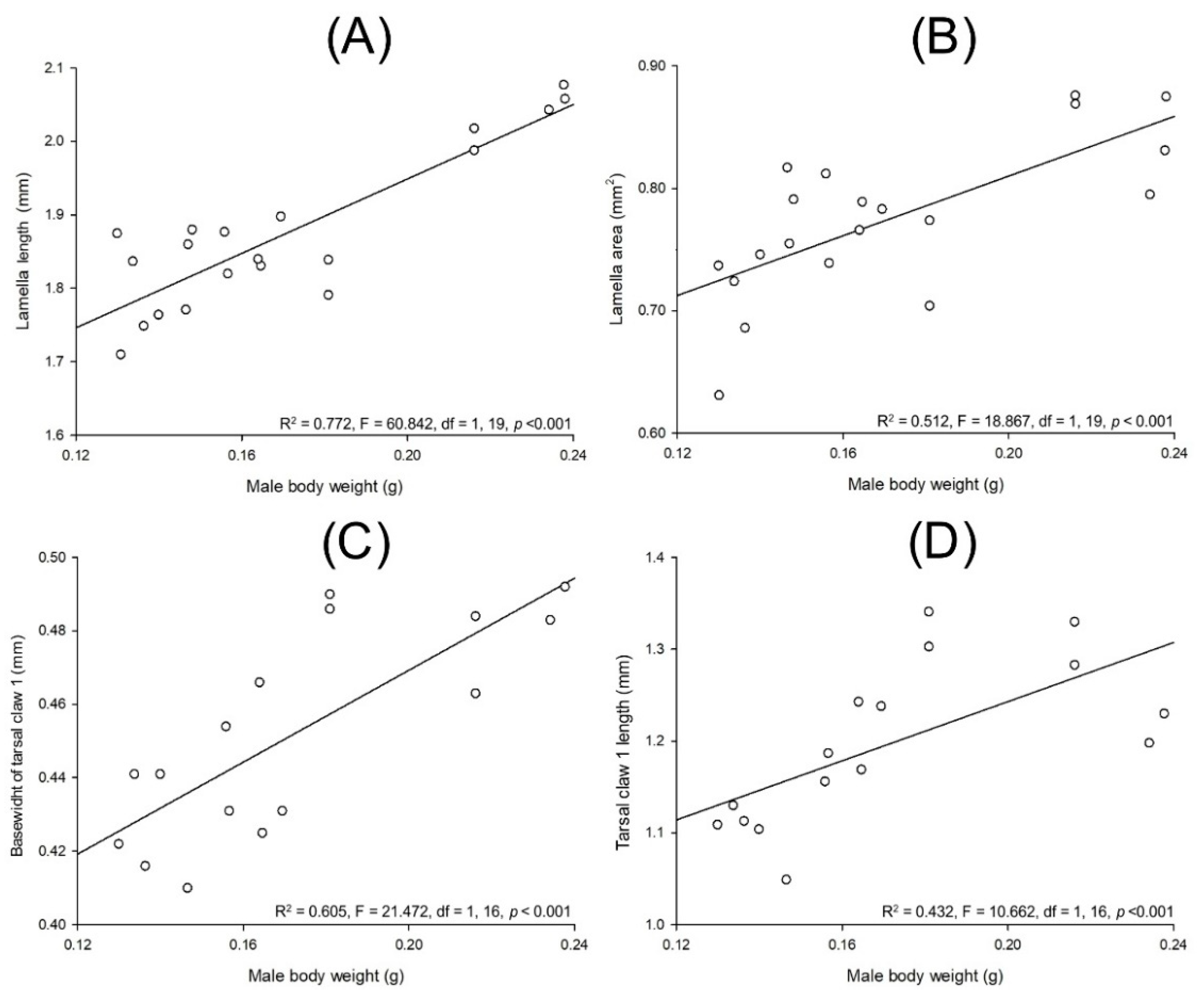
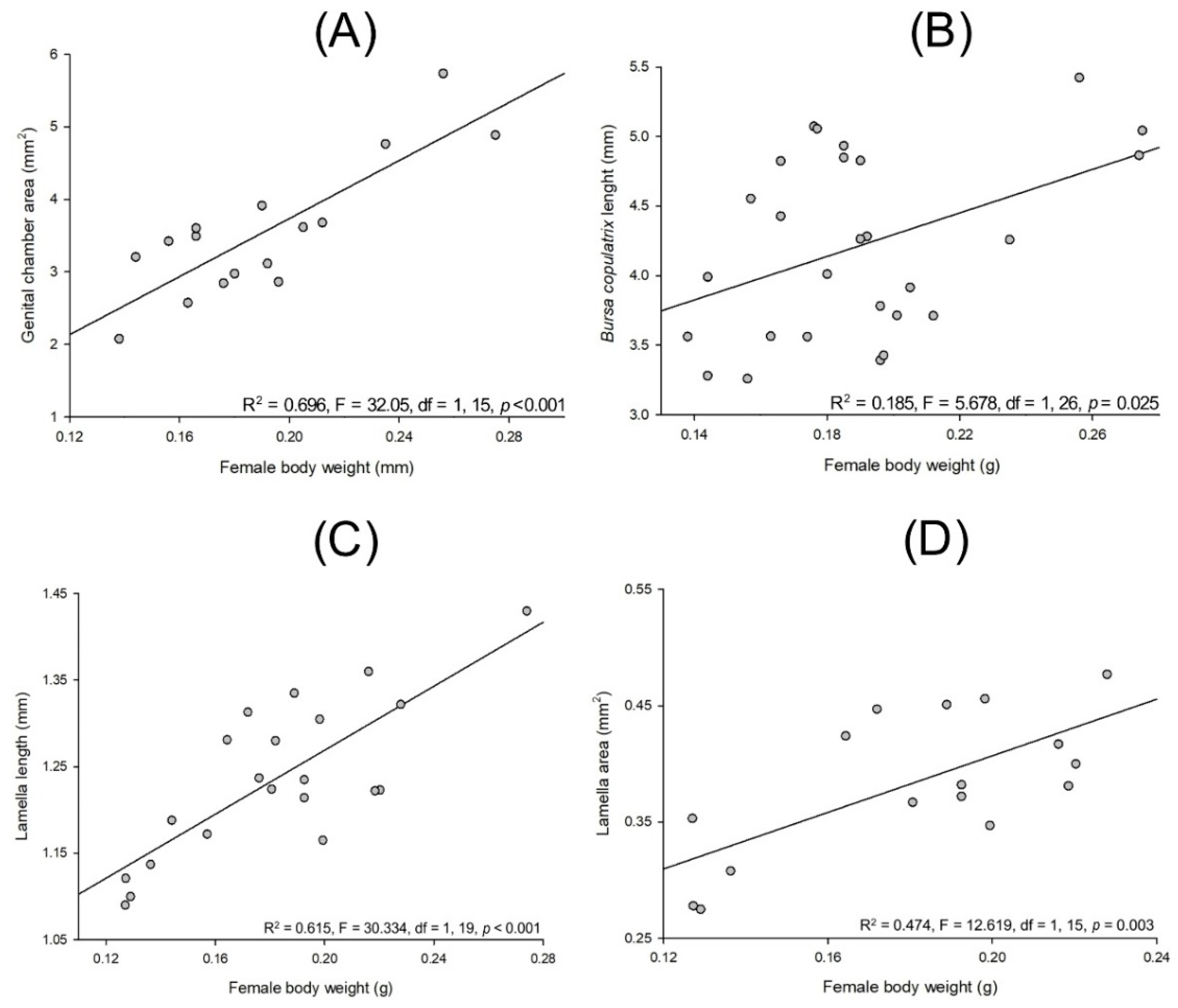
3.3. Allometry in Cyclocephala barrerai
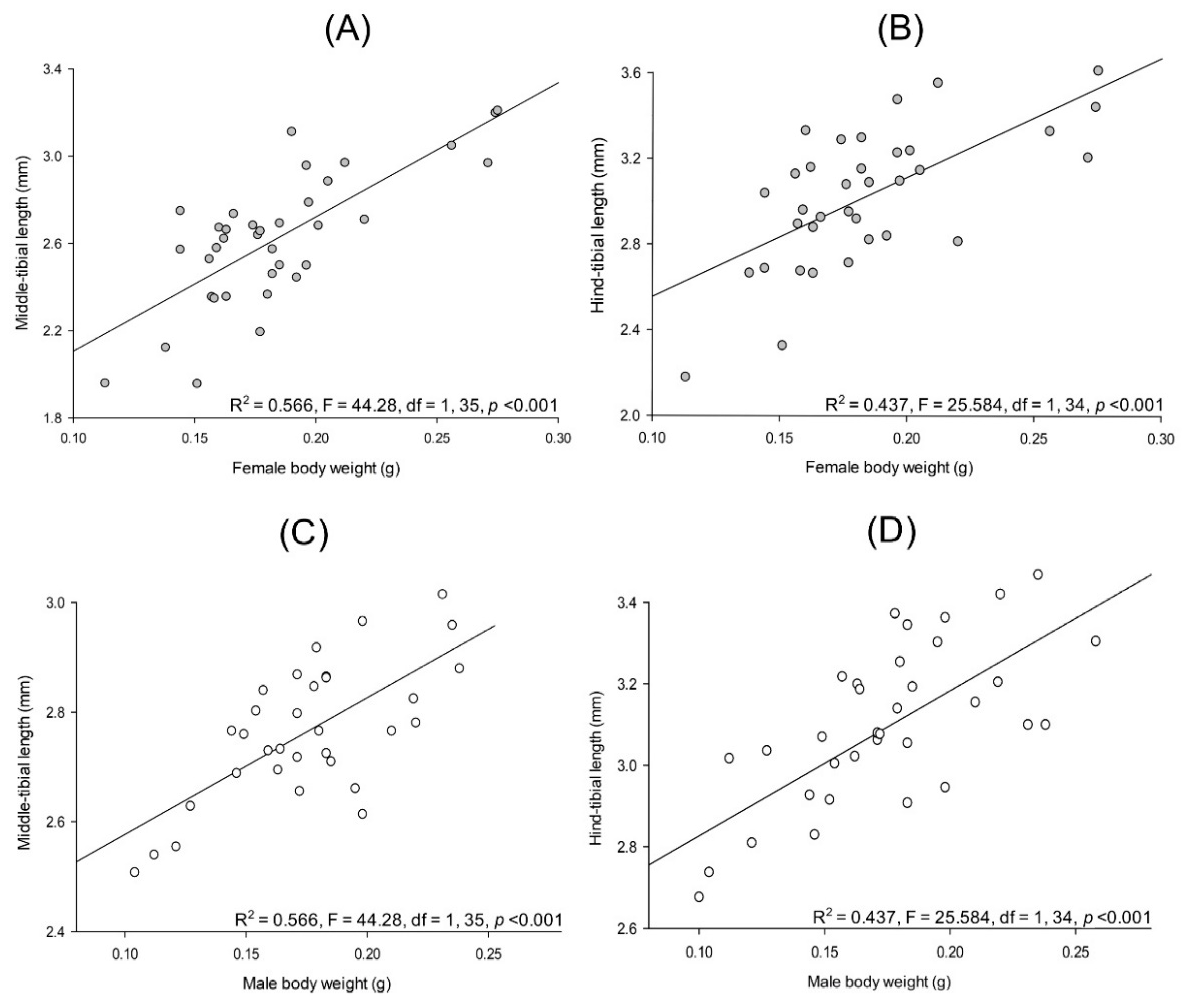
3.3.1. Relationships between Body Parts and Weight
3.3.2. Sexual Dimorphism in Lab-Reared Adults
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thornhill, R.; Alcock, J. The Evolution of Insect Mating Systems; Harvard University Press: London, UK, 2013; pp. 28–50. [Google Scholar]
- Chen, S. Biochemistry of insect male accessory glands. Annu. Rev. Entomol. 1984, 29, 233–255. [Google Scholar] [CrossRef]
- Honěk, A. Intraspecific variation in body size and fecundity in insects: A general relationship. Oikos 1993, 66, 492. [Google Scholar] [CrossRef]
- Roff, D.A. Life History Evolution; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Wilson, D.S.; Knollenberg, W.G. Food discrimination and ovarian development in burying beetles (Coleoptera: Silphidae: Nicrophorus). Ann. Entomol. Soc. Am. 1984, 77, 165–170. [Google Scholar] [CrossRef]
- Gillott, C. Male accessory gland secretions: Modulators of female reproductive physiology and behavior. Annu. Rev. Entomol. 2003, 48, 163–184. [Google Scholar] [CrossRef]
- Zhao, M.-T.; Wang, Y.; Zhou, Z.-S.; Wang, R.; Guo, J.-Y.; Wan, F.-H. Effects of periodically repeated heat events on reproduction and ovary development of Agasicles hygrophila (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2016, 109, 1586–1594. [Google Scholar] [CrossRef]
- Benítez-Herrera, L.N.; Martínez-Morales, I.; Romero-López, A.A. Anatomy of the reproductive system of Macrodactylus mexicanus (Coleoptera: Scarabaeoidea: Melolonthidae) and its possible relationship with sexual chemical communication. Southwest. Entomol. 2015, 40, 189–198. [Google Scholar] [CrossRef]
- Martínez-Morales, I.; Rivera-Gasperín, S.L.; Carrillo-Ruiz, H.; Morón, M.A. Comparative aspects of the internal reproductive system of males in species of Melolonthinae, Dynastinae, and Rutelinae (Coleoptera: Scarabaeoidea) from Mexico. Acta Zool. Mex. 2021, 37, 1–17. [Google Scholar] [CrossRef]
- Cristóvão, J.P.; Vaz-de-Mello, F.Z. The terminalia of the superfamily Scarabaeoidea (Coleoptera): Specific glossary, dissecting methodology, techniques and previously unrecorded sexual dimorphism in some difficult groups. Zool. J. Linn. Soc. 2021, 191, 1001–1043. [Google Scholar] [CrossRef]
- Sanmartín, I.; Martín-Piera, F. First phylogenetic analysis of the subfamily Pachydeminae (Coleoptera, Scarabaeoidea, Melolonthidae): The Palearctic Pachydeminae. J. Zool. Syst. Evol. Res. 2003, 41, 2–46. [Google Scholar] [CrossRef]
- Richmond, M.P.; Park, J.; Henry, C.S. The function and evolution of male and female genitalia in Phyllophaga Harris scarab beetles (Coleoptera: Scarabaeidae). J. Evol. Biol. 2016, 29, 2276–2288. [Google Scholar] [CrossRef] [Green Version]
- Carrillo-Ruiz, H.; Martínez, M.I.; Morón, M.-A. Comparative study of the reproductive system of two species of Hoplia (Coleoptera: Scarabaeidae: Hopliinae). Proc. Entomol. Soc. Wash. 2008, 110, 778–788. [Google Scholar] [CrossRef]
- Romero-López, A.; Arzuffi, R.; Valdez, J.; Sánchez-Espíndola, E.; Morón, M.A. Tissues involved in sex pheromone production in Phyllophaga obsoleta (Coleoptera: Scarabaeoidea: Melolonthidae). Ann. Entomol. Soc. Am. 2011, 104, 960–965. [Google Scholar] [CrossRef] [Green Version]
- Yi-Zhen, L.; Lian-Xin, L.; Yan-Jie, S. Variation in the reproduction of some phytophagous chafers and its bearing on pest control. Acta Entomol. Sin. 1987, 30, 152. [Google Scholar]
- Stringer, I.A.N. The female reproductive system of Costelytra zealandica (White) (Coleoptera: Scarabaeidae: Melolonthinae). N. Z. J. Zool. 1988, 15, 513–533. [Google Scholar] [CrossRef] [Green Version]
- Gayon, J. History of the concept of allometry. Am. Zool. 2000, 40, 748–758. [Google Scholar] [CrossRef] [Green Version]
- Simmons, L.W.; Tomkins, J.L.; Hunt, J. Sperm competition games played by dimorphic male beetles. Proc. R. Soc. B Biol. Sci. 1999, 266, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Okada, K.; Miyatake, T. Sexual dimorphism in mandibles and male aggressive behavior in the presence and absence of females in the beetle Librodor japonicus (Coleoptera: Nitidulidae). Ann. Entomol. Soc. Am. 2004, 97, 1342–1346. [Google Scholar] [CrossRef]
- Okada, K.; Miyatake, T.; Nomura, Y.; Kuroda, K. Fighting, dispersing, and sneaking: Body-size dependent mating tactics by male Librodor japonicus beetles. Ecol. Entomol. 2008, 33, 269–275. [Google Scholar] [CrossRef]
- Cayetano, L.; Maklakov, A.A.; Brooks, R.C.; Bonduriansky, R. Evolution of male and female genitalia following release from sexual selection. Evolution 2011, 65, 2171–2183. [Google Scholar] [CrossRef]
- Rowland, J.M.; Qualls, C.R.; Beaudoin-Ollivier, L. Discrimination of alternative male phenotypes in Scapanes australis (Boisduval) (Coleoptera: Scarabaeidae: Dynastinae). Aust. J. Entomol. 2005, 44, 22–28. [Google Scholar] [CrossRef]
- McCullough, E.L.; Ledger, K.J.; O’Brien, D.M.; Emlen, D.J. Variation in the allometry of exaggerated rhinoceros beetle horns. Anim. Behav. 2015, 109, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, H.; Carrillo-Ruiz, H.; Morón, M. Horns positive allometry in a Mexican population of Strategus aloeus (L.) (Coleoptera: Scarabaeoidea: Dynastinae). Entomotropica 2013, 28, 87–94. [Google Scholar]
- Johnson, J.P. Cyclocephala (ochrosidia) borealis in Connecticut 1. J. Agric. Res. 1941, 62, 79–86. [Google Scholar]
- Maia, A.C.D.; Schlindwein, C. Caladium bicolor (Araceae) and Cyclocephala celata (Coleoptera, Dynastinae): A well-established pollination system in the Northern Atlantic rainforest of Pernambuco, Brazil. Plant Biol. 2006, 8, 529–534. [Google Scholar] [CrossRef]
- Duchini, P.G.; Echeverria, J.R.; Américo, L.F.; Guzatti, G.C.; Cherman, M.A.; Sbrissia, A.F. White grubs (Cyclocephala flavipennis) damaging perennial winter pastures in the south region of Brazil. Ciência Rural 2017, 47, e20160662. [Google Scholar] [CrossRef]
- Parizotto, D.R.; Grossi, P.C. Revisiting pollinating Cyclocephala scarab beetles (Coleoptera: Melolonthidae: Dynastinae) associated with the soursop (Annona muricata, Annonaceae). Neotrop. Entomol. 2019, 48, 415–421. [Google Scholar] [CrossRef]
- Martínez-Morales, I.; Morón, M.A. Female reproductive systems in Melolonthinae, Rutelinae, and Dynastinae (Coleoptera: Scarabaeoidea, Melolonthidae). Southwest. Entomol. 2015, 40, 369–386. [Google Scholar] [CrossRef]
- Sanchez-Cruz, A.; Robledo, N.; Rosete-Enríquez, M.; Romero-López, A.A. Attraction of adults of Cyclocephala lunulata and Cyclocephala barrerai (Coleoptera: Scarabaeoidea: Melolonthidae) towards Bacteria Volatiles Isolated from Their Genital Chambers. Molecules 2020, 25, 4430. [Google Scholar] [CrossRef]
- Aragón, A.; Morón, M.A.; Tapia-Rojas, A.M.; Rojas-García, R. Fauna de Coleoptera Melolonthidae en el rancho “La Joya”, Atlixco, Puebla, México. Acta Zool. Mex. 2001, 164, 143–164. [Google Scholar] [CrossRef]
- Morón, M.A.; Lugo-García, G.A.; Aragón-García, A. Description of the third instar larvae of five species of Cyclocephala (Coleoptera, Melolonthidae, Dynastinae) from Mexico. Rev. Bras. Entomol. 2014, 58, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Moore, M.R.; Cave, R.D.; Branham, M.A. Annotated catalog and bibliography of the cyclocephaline scarab beetles (Coleoptera, Scarabaeidae, Dynastinae, Cyclocephalini). Zookeys 2018, 745, 101–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, S.S.; Tang, X.M. Manipulating biological samples for environmental scanning electron microscopy observation. Scanning 2001, 23, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Kirk, S.E.; Skepper, J.N.; Donald, A.M. Application of environmental scanning electron microscopy to determine biological surface structure. J. Microsc. 2009, 233, 205–224. [Google Scholar] [CrossRef]
- Carson, F.L.; Martin, J.H.; Lynn, J.A. Formalin fixation for electron microscopy: A re-evaluation. Am. J. Clin. Pathol. 1973, 59, 365–373. [Google Scholar] [CrossRef]
- García-Hernández, C.; Rojo-Rubio, R.; Olmedo-Juárez, A.; Zamilpa, A.; Mendoza de Gives, P.; Antonio-Romo, I.A.; Aguilar-Marcelino, L.; Arece-García, J.; Tapia-Maruri, D.; González-Cortazar, M. Galloyl derivatives from Caesalpinia coriaria exhibit in vitro ovicidal activity against cattle gastrointestinal parasitic nematodes. Exp. Parasitol. 2019, 200, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Núñez, E.J.; Zamilpa, A.; González-Cortazar, M.; Olmedo-Juárez, A.; Cardoso-Taketa, A.; Sánchez-Mendoza, E.; Tapia-Maruri, D.; Salinas-Sánchez, D.O.; Mendoza-De Gives, P. Isorhamnetin: A nematocidal flavonoid from Prosopis laevigata leaves against Haemonchus contortus eggs and larvae. Biomolecules 2020, 10, 773. [Google Scholar] [CrossRef]
- Grodowitz, M.J.; Brewer, F.D. Ovarian anatomy and physiological age-grading of the female boll weevil, Anthonomus grandis grandis Boheman (Coleoptera: Curculionidae). Ann. Entomol. Soc. Am. 1987, 80, 642–651. [Google Scholar] [CrossRef]
- Rasband, W.S. ImageJ Version 1.53s, Rasband, IL, USA; US National Institutes of Health: Bethesda, MD, USA, 1997–2009.
- Wang, Q.; Zeng, W.-Y.; Li, J.-S. Reproductive behavior of Paraglenea fortunei (Coleoptera: Cerambycidae). Ann. Entomol. Soc. Am. 1990, 83, 860–866. [Google Scholar] [CrossRef]
- Hoffman, G.D.; Raffa, K.F. Maturation of the male pales weevil (Coleoptera: Curculionidae) reproductive system and its effect on male response to females. Ann. Entomol. Soc. Am. 1992, 85, 571–577. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, G.; Davis, L.K. Reproductive potential and daily reproductive rhythms of Oemona hirta (Coleoptera: Cerambycidae). J. Econ. Entomol. 1998, 91, 1360–1365. [Google Scholar] [CrossRef]
- Breeschoten, T.; Clark, D.R.; Schilthuizen, M. Evolutionary patterns of asymmetric genitalia in the beetle tribe cyclocephalini (Coleoptera: Scarabaeidae: Dynastinae). Contrib. Zool. 2013, 82, 95–106. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Leal, W.S. Ultrastructure of pheromone-detecting sensillum placodeum of the Japanese beetle, Popillia japonica Newmann (Coleoptera: Scarabaeidae). Arthropod Struct. Dev. 2000, 29, 121–128. [Google Scholar] [CrossRef]
- Romero-López, A.; Morón, M.; Valdez, J. Sexual dimorphism in antennal receptors of Phyllophaga ravida Blanchard (Coleoptera: Scarabaeoidea: Melolonthidae). Neotrop. Entomol. 2010, 39, 957–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bal, N.; Özkdikmen, H. Aedeagus structure of Cassida prasina Illiger, 1798 (Coleoptera: Chrysomelidae: Cassidinae) in Scanning Electrone Microscope (SEM). KSU J. Agric. Nat. 2020, 23, 748–753. [Google Scholar] [CrossRef]
- Özdikmen, Ü.; Bal, N.; Amutkan Mutlu, D.; Suludere, Z. A SEM study of the aedeagus and spermatheca of Cassida viridis Linnaeus, 1758 (Coleoptera: Chrysomelidae: Cassidinae) from Turkey. Turk. Entomoloji Derg. 2020, 44, 355–364. [Google Scholar] [CrossRef]
- Özyurt Koçakoğlu, N.; Candan, S.; Güllü, M. Anatomy and histology of reproductive system of adult male mint leaf beetle Chrysolina herbacea (Duftschmid, 1825) (Coleoptera: Chrysomelidae). Microsc. Res. Tech. 2021, 84, 512–520. [Google Scholar] [CrossRef]
- Acebes, A.; Cobb, M.; Ferveur, J.F. Species-specific effects of single sensillum ablation on mating position in Drosophila. J. Exp. Biol. 2003, 206, 3095–3100. [Google Scholar] [CrossRef] [Green Version]
- Schubert, L.F.; Krüger, S.; Moritz, G.B.; Schubert, V. Male reproductive system and spermatogenesis of Limodromus assimilis (Paykull 1790). PLoS ONE 2017, 12, e0180492. [Google Scholar] [CrossRef] [Green Version]
- Pascini, T.V.; Martins, G.F. The insect spermatheca: An overview. Zoology 2017, 121, 56–71. [Google Scholar] [CrossRef]
- Eberhard, W.G. Rapid divergent evolution of sexual morphology: Comparative tests of antagonistic coevolution and traditional female choice. Evolution 2004, 58, 1947–1970. [Google Scholar] [CrossRef]
- Shapiro, A. The lock-and-key hypothesis: Evolutionary and biosystematic interpretation of insect genitalia. Annu. Rev. Entomol. 1989, 34, 231–245. [Google Scholar] [CrossRef]
- Fernadez, M.; Salgado, J.; Pajares, J. The seasonal development of the gonads and fat content of Tomicus minor (Coleoptera Scolytidae). Belg. J. Entomol. 1999, 1, 311–324. [Google Scholar]
- Vega-Petlacalco, M.; Arzuffi, R.; Valdez, J.; Rodríguez-Monroy, M.; Jiménez-Pérez, A.; Robledo, N. Food quality influences ovarian development in Scyphophorus acupunctatus (Coleoptera: Dryophthoridae). Fla. Entomol. 2018, 101, 447–452. [Google Scholar] [CrossRef] [Green Version]
- Sasakawa, K. Diet affects male gonad maturation, female fecundity, and larval development in the granivorous ground beetle Anisodactylus punctatipennis. Ecol. Entomol. 2009, 34, 406–411. [Google Scholar] [CrossRef]
- Ghoneim, K.K.; Abdel-Khaliq, A.A.; Bream, A.S.; Emam, D.M. Effects of food type on the adult performace of black blister beetle Meloe proscaeabaeus (Coleoptera: Meloidae). Int. J. Biol. Sci. 2012, 1, 5–17. [Google Scholar]
- Zhou, P.; Yang, H.; Jin, D.C.; He, X.Z.; Wang, Q. Sex-specific allometry of morphometric and reproductive traits in oriental fruit flies (Diptera: Tephritidae). J. Econ. Entomol. 2016, 109, 1249–1253. [Google Scholar] [CrossRef]
- Carne, P.B. Cyclocephala signaticollis Burmeister, an introduced pasture scarab (Coleoptera). Proc. Linn. Soc. N. S. W. 1957, 81, 217–221. [Google Scholar]
- Stern, D.L.; Emlen, D.J. The developmental basis for allometry in insects. Development 1999, 126, 1091–1101. [Google Scholar] [CrossRef]
- Vera-Cano, D.A.; Álvarez, H.A.; Morón, M.A. Positive allometry of horns in the rhinoceros beetle Golofa xiximeca does not follow breaking-point patterns. Southwest. Entomol. 2017, 42, 933–940. [Google Scholar] [CrossRef]
- Moczek, A.P. Pupal remodeling and the evolution and development of alternative male morphologies in horned beetles. BMC Evol. Biol. 2006, 7, 711–729. [Google Scholar] [CrossRef] [Green Version]
- Knell, R.J.; Pomfret, J.C.; Tomkins, J.L. The limits of elaboration: Curved allometries reveal the constraints on mandible size in stag beetles. Proc. R. Soc. B Biol. Sci. 2004, 271, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Heddleston, J.M.; Aaron, J.S.; Khuon, S.; Chew, T.L. A guide to accurate reporting in digital image acquisition–can anyone replicate your microscopy data? J. Cell Sci. 2021, 134, jcs254144. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.; McNeill, B.; Irwin, M. Nondestructive Imaging of Pin-Mounted Museum Insect Specimens Using the Field-Emission Environmental Scanning Electron Microscope (ESEM-FEG). Microsc. Microanal. 1999, 5, 338–339. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Structure | Lab-Reared Adults Mean ± SEM (n) | |
|---|---|---|
| Female | Male | |
| Pupa weight | 0.308 ± 0.006 g (58) | 0.291 ± 0.006 g (58) |
| Adult weight | 0.188 ± 0.004 g (58) | 0.175 ± 0.004 g (58) |
| Middle-tibial length | 2.625 ± 0.051 mm (36) | 2.770 ± 0.022 mm (33) |
| Hind-tibial length | 3.025 ± 0.054 mm (35) | 3.077 ± 0.048 mm (36) |
| Lamella length | 1.236 ± 0.089 mm (21) | 1.876 ± 0.024 mm (20) |
| Lamella area | 0.383 ± 0.015 mm2 (16) | 0.775 ± 0.014 mm2 (20) |
| Tarsal claw 1 length | - | 1.199 ± 0.0218 mm (16) |
| Tarsal claw 1 width base | - | 0.452 ± 0.007 mm (16) |
| Tarsal claw 2 length | - | 0.839 ± 0.065 mm (15) |
| Tarsal claw 2 width base | - | 0.0388 ± 0.008 mm (15) |
| Genital chamber length | 1.993 ± 0.109 mm (19) | - |
| Genital chamber area | 3.875 ± 0.291 mm2 (16) | - |
| Bursa copulatrix length | 4.223 ± 0.145 mm (27) | - |
| Bursa copulatrix area | 5.089 ± 0.357 mm2 (26) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez-Cruz, A.; Tapia-Maruri, D.; Jiménez-Pérez, A. Reproductive Apparatus, Gonadic Maturation, and Allometry of Cyclocephala barrerai Martínez (Coleoptera: Melolonthidae: Dynastinae). Insects 2022, 13, 638. https://doi.org/10.3390/insects13070638
Sanchez-Cruz A, Tapia-Maruri D, Jiménez-Pérez A. Reproductive Apparatus, Gonadic Maturation, and Allometry of Cyclocephala barrerai Martínez (Coleoptera: Melolonthidae: Dynastinae). Insects. 2022; 13(7):638. https://doi.org/10.3390/insects13070638
Chicago/Turabian StyleSanchez-Cruz, Abraham, Daniel Tapia-Maruri, and Alfredo Jiménez-Pérez. 2022. "Reproductive Apparatus, Gonadic Maturation, and Allometry of Cyclocephala barrerai Martínez (Coleoptera: Melolonthidae: Dynastinae)" Insects 13, no. 7: 638. https://doi.org/10.3390/insects13070638