
Odonata Assemblages as a Tool to Assess the Conservation Value of Intermittent Rivers in the Mediterranean


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Odonata Survey
2.3. Data Analyses
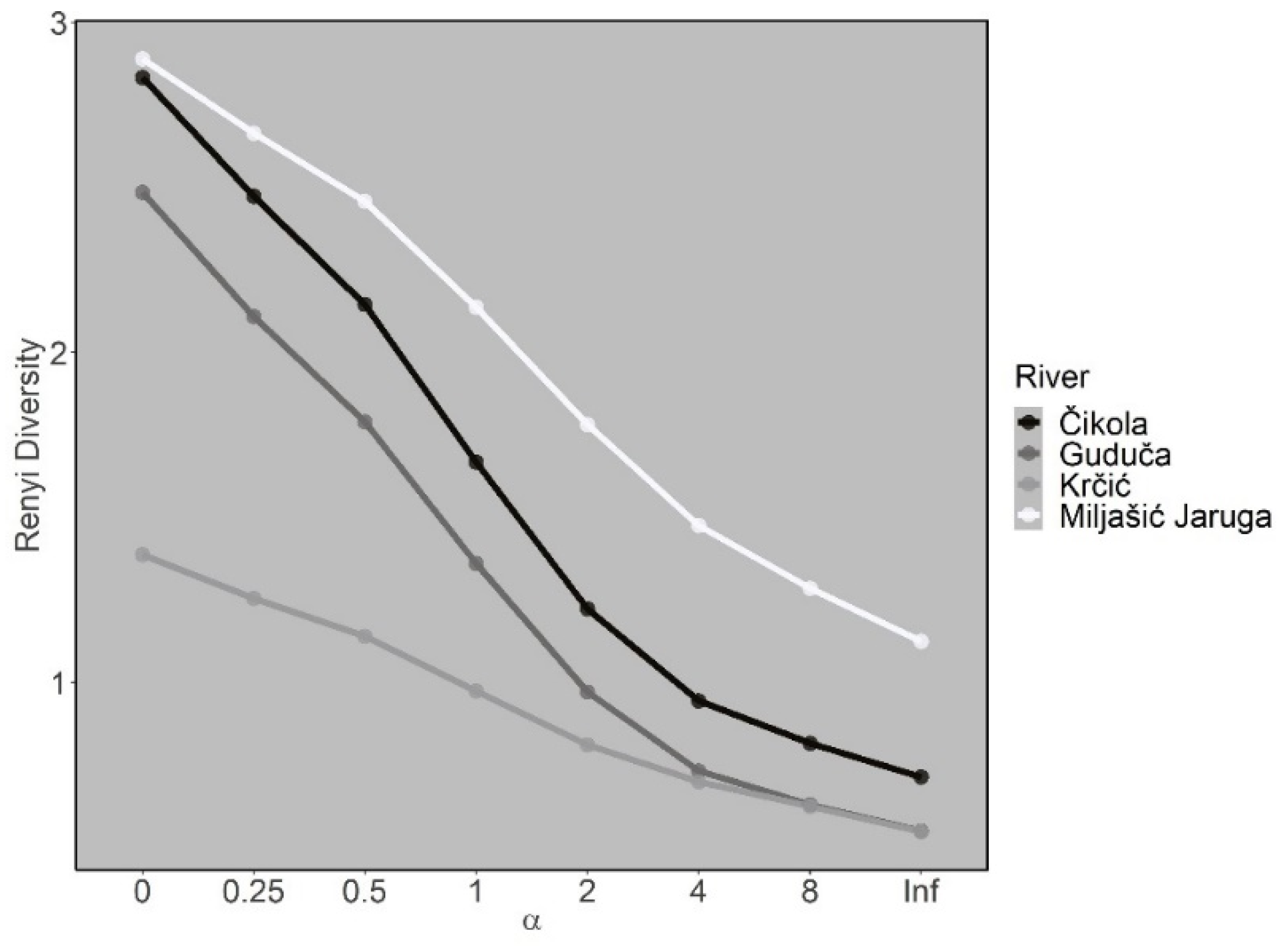
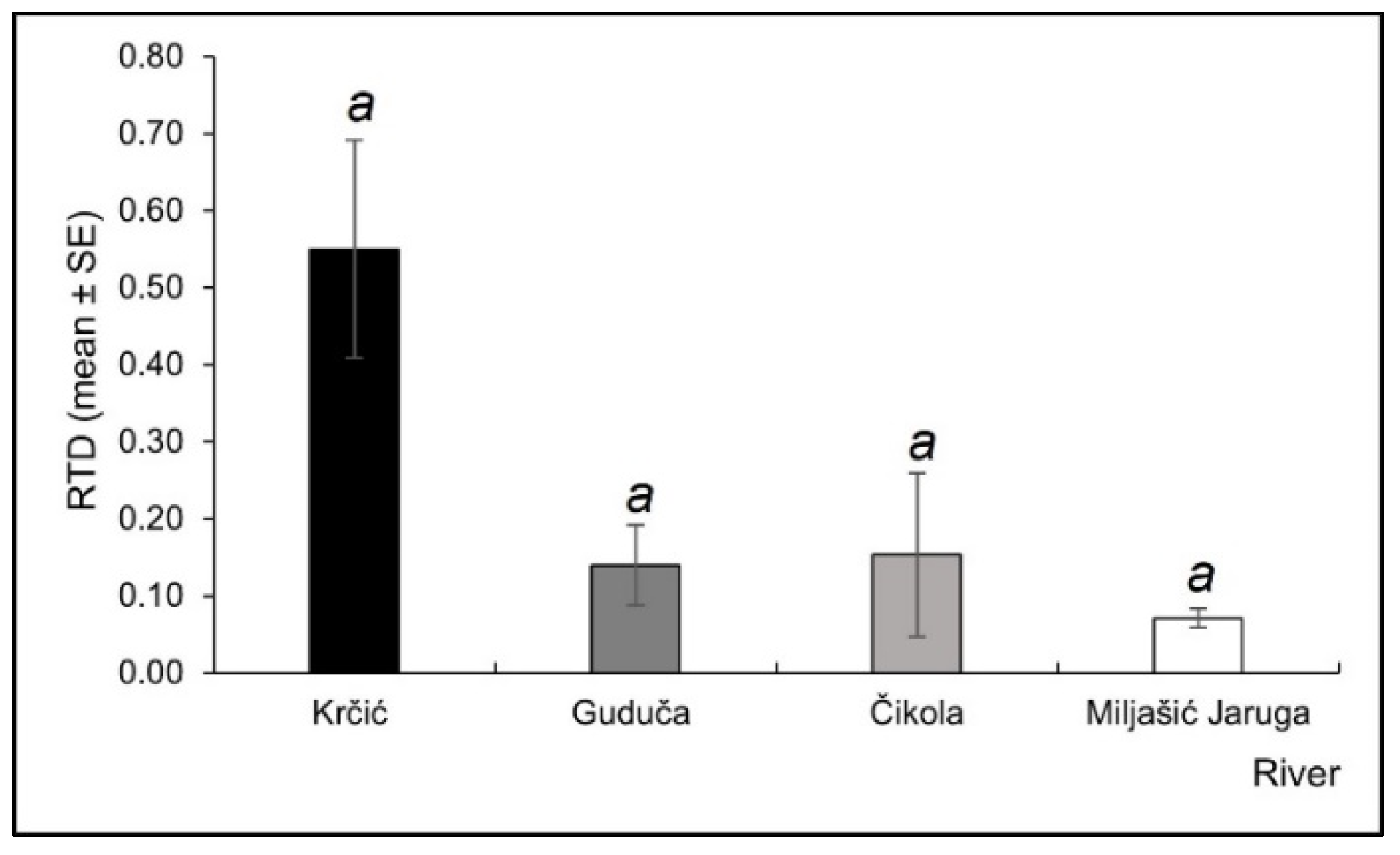
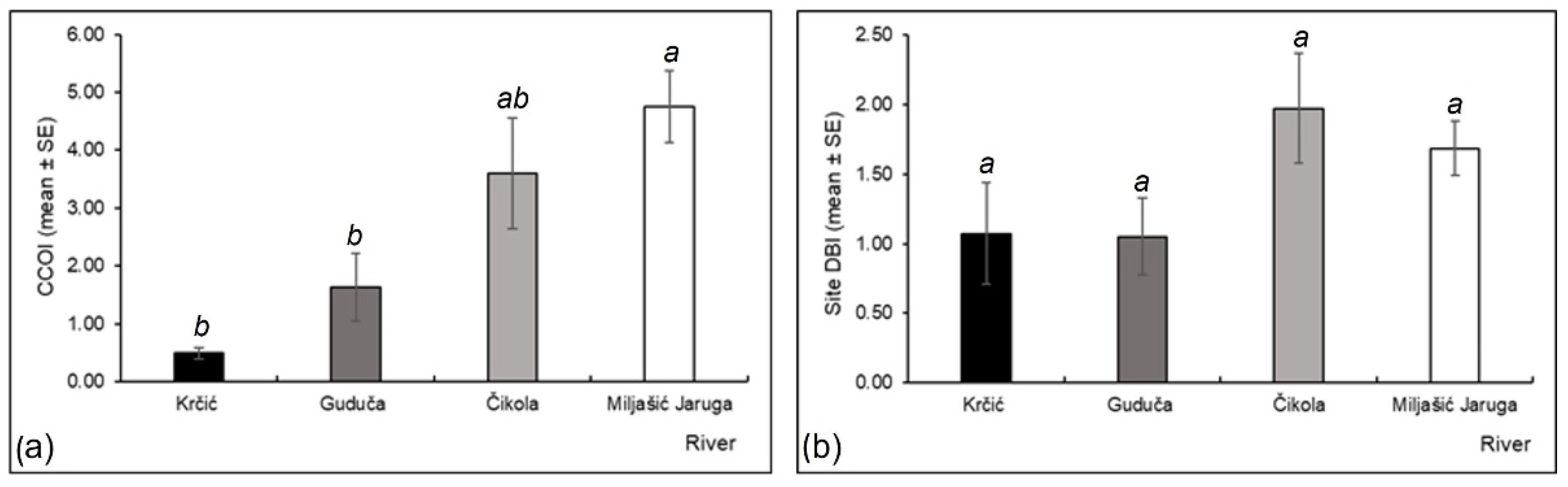
3. Results
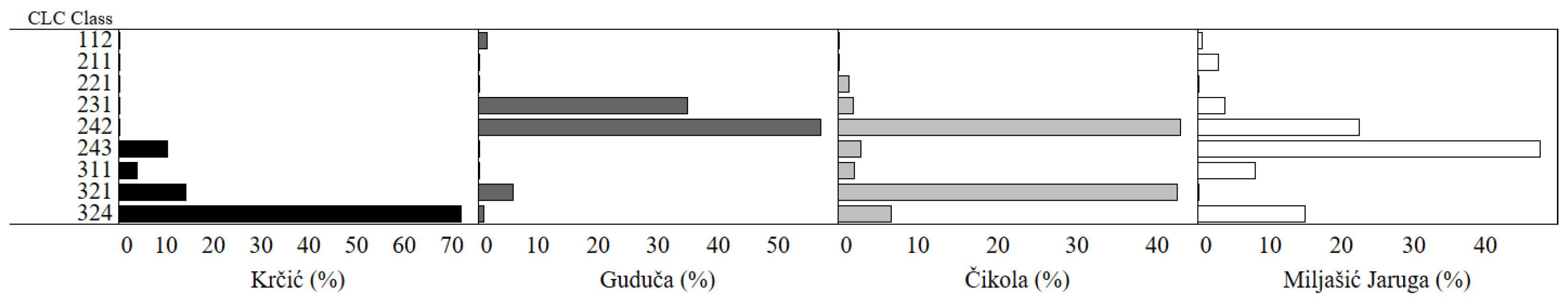
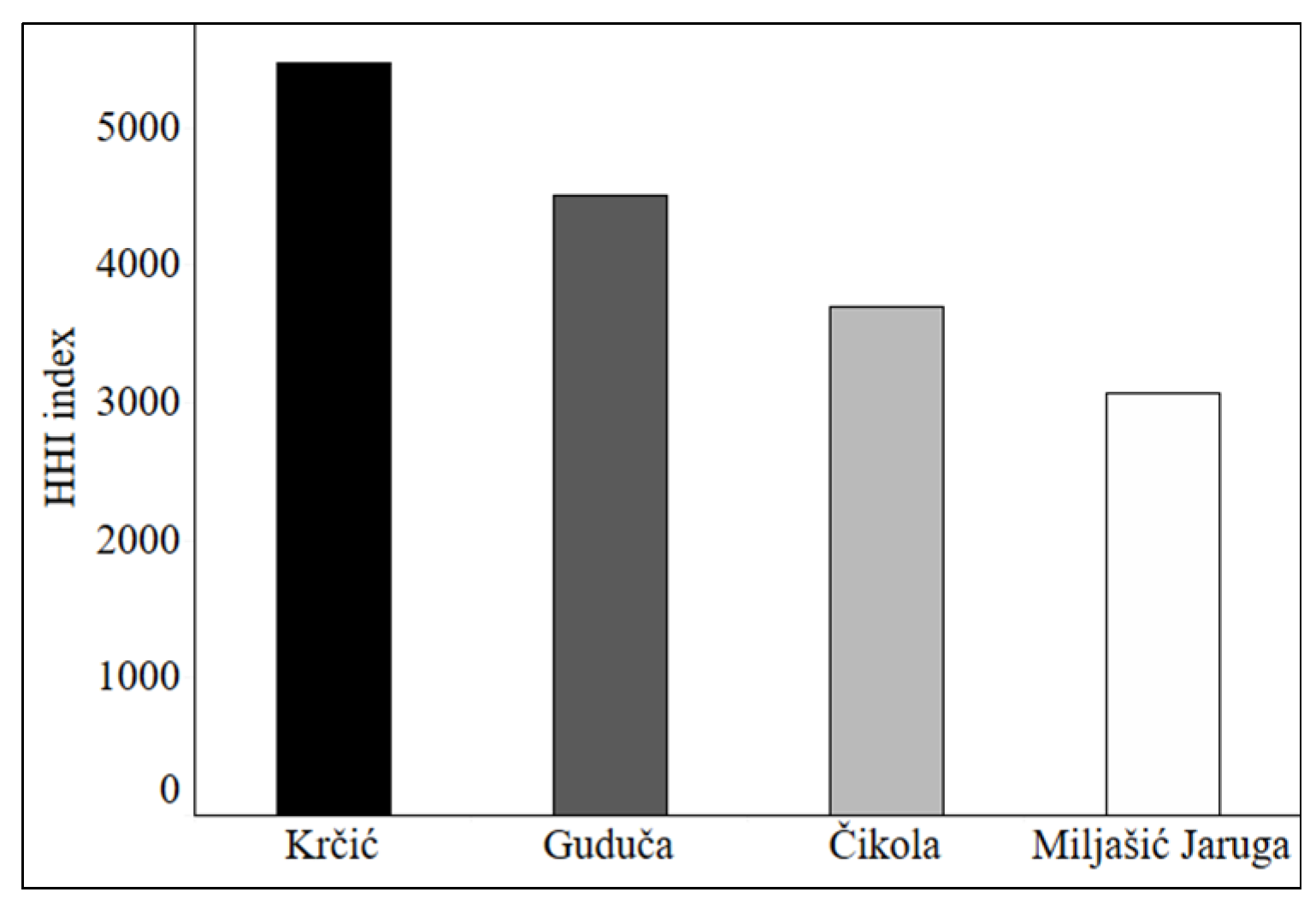
3.1. Land Cover
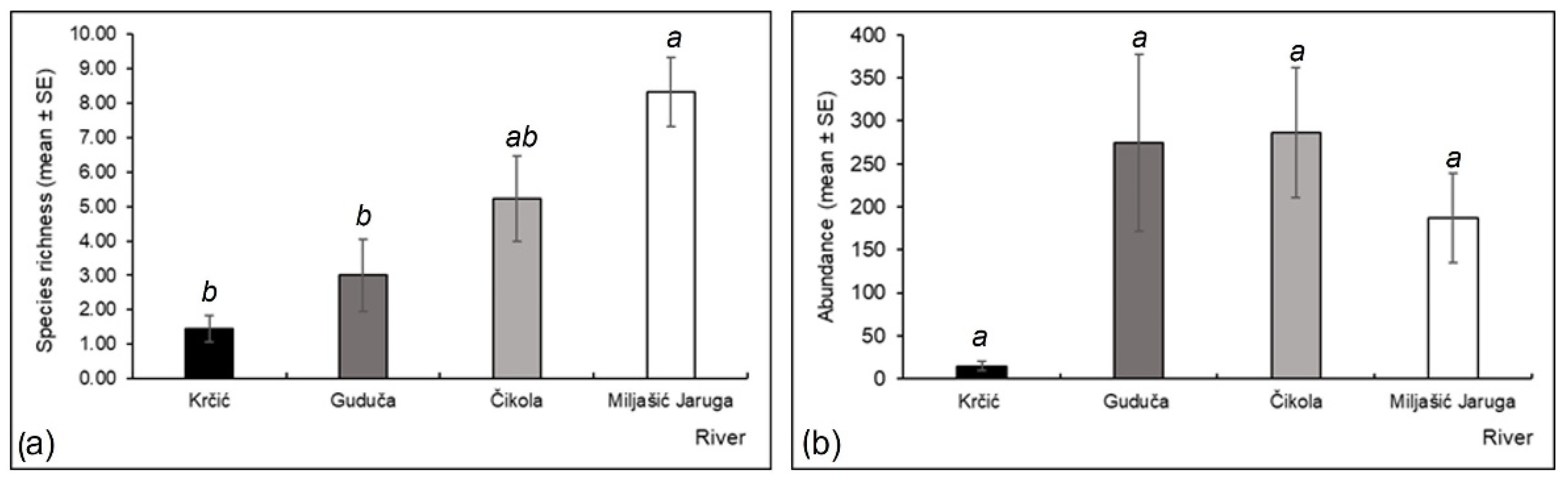
3.2. Odonata Species Occurrence
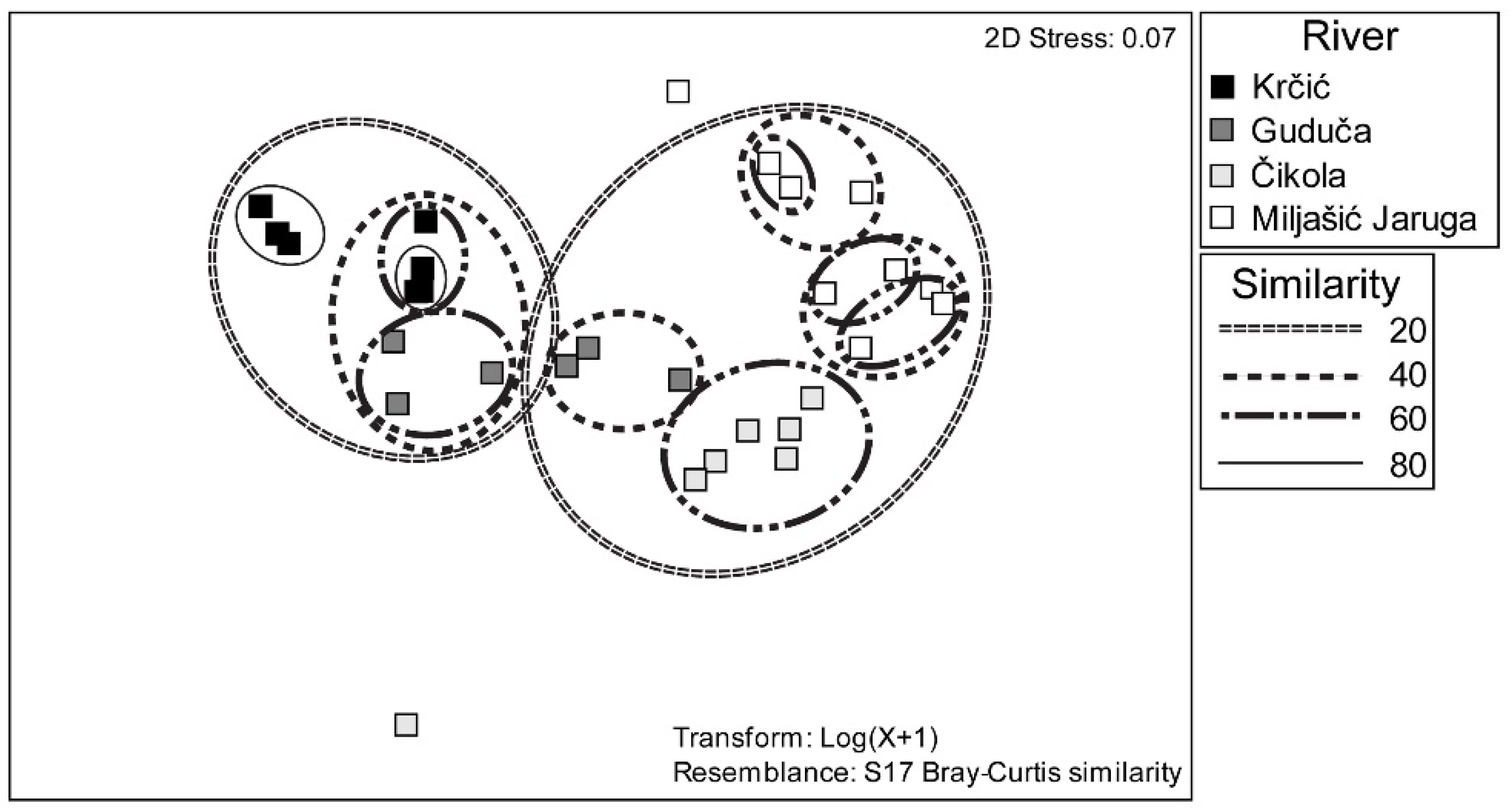
3.3. Odonata Assemblages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | KR1 | KR2 | KR3 | GU1 | GU2 | GU3 | CI1 | CI2 | CI3 | JA1 | JA2 | JA3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Coordinates, N | 44.02734 | 44.02761 | 44.0419 | 43.92848 | 43.92358 | 43.89061 | 43.84345 | 43.84572 | 43.83771 | 44.1942 | 44.20716 | 44.2198 |
| Coordinates, E | 16.31899 | 16.30671 | 16.25358 | 15.83178 | 15.80045 | 15.7996 | 16.25727 | 16.17832 | 16.04927 | 15.27819 | 15.25737 | 15.23919 |
| Altitude (m a.s.l.) | 363 | 341 | 267 | 134 | 108 | 101 | 271 | 263 | 89 | 23 | 17 | 8 |
| Substrates (%) | 60% PHY, 20% MES, 20% MIC | 60% PHY, 20% MES, 20% MIC | 40% PHY, 40% MEG, 10% XYL, 10% PSA | 60% PHY, 20% MEG, 20% MES | 20% MES, 20% XYL, 60% PHY | 10% MIC, 10% MES, 80% PHY | 80% PHY, 20% MEG | 60% PHY, 30% ARG, 10% AK | 10% PHY, 20% MIC, 20% XYL, 50% MEG | 70% PHY, 30% MES | 50% PHY, 30% MIC, 20% ARG | 80% PHY, 20% ARG |
| Average width (m) | 17.5 | 12 | 12.5 | 6.5 | 6.5 | 8 | 17.5 | 24.3 | 22.5 | 6 | 2.8 | 3.8 |
| Shading | Sunny | Sunny | Mostly shaded | Mostly shaded | Sunny | Sunny | Sunny | Partially shaded | Sunny | Sunny | Sunny | Sunny |
| Riparian vegetation | High bushes, low trees | High bushes, low trees | Forest | Trees, bushes | Grassland, sporadically trees | Grassland, bushes | Grassland, sporadically bushes and trees | Bushes, trees | Bushes, trees | Grassland, sporadically trees | Grassland, bushes, trees | Grassland |
| Aquatic vegetation | Submerged 1, 2 | Submerged 1 | Submerged 1 | Submerged 1, emergent | Submerged 2, emergent | Submerged 1 | Submerged 2, dense emergent, floating | Submerged 2, dense emergent | Submerged 1 | Submerged 2, dense emergent, floating | Submerged 2, dense emergent, floating | Submerged 2, dense emergent, floating |
| Species/Study Site | K1 | K2 | K3 | GU1 | GU2 | GU3 | CI1 | CI2 | CI3 | JA1 | JA2 | JA3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Calopteryx splendens (Harris, 1782) | 18 | 7 | 230 | 3 | ||||||||
| Calopteryx virgo (Linnaeus, 1758) | 3 | 62 | 39 | 460 | 70 | |||||||
| Chalcolestes viridis (Vander Linden, 1825) | 100 | 320 | 250 | 5 | ||||||||
| Sympecma fusca (Vander Linden, 1820) | 100 | 102 | 20 | |||||||||
| Ischnura elegans (Vander Linden, 1820) | 50 | 30 | 20 | 130 | 82 | 58 | ||||||
| Coenagrion puella (Linnaeus, 1758) | 70 | 130 | 85 | 107 | ||||||||
| Erythromma lindenii (Selys, 1840) | 70 | 140 | 8 | |||||||||
| Platycnemis pennipes (Pallas, 1771) | 20 | 1500 | 900 | 1000 | 50 | 50 | ||||||
| Aeshna affinis Vander Linden, 1820 | 4 | 1 | ||||||||||
| Aeshna isoceles (Müller, 1767) | 18 | 13 | ||||||||||
| Anax imperator (Selys, 1839) | 2 | 8 | 6 | 9 | 5 | |||||||
| Brachytron pratense (Müller, 1764) | 3 | 8 | 5 | 6 | 8 | 4 | ||||||
| Onychogomphus forcipatus (Linnaeus, 1758) | 36 | 51 | ||||||||||
| Cordulegaster heros (Theischinger, 1979) | 3 | |||||||||||
| Somatochlora meridionalis (Nielsen, 1935) | 1 | 2 | 1 | 4 | 2 | 7 | 2 | |||||
| Libellula depressa (Linnaeus, 1758) | 2 | 2 | 17 | 4 | ||||||||
| Libellula fulva (Müller, 1764) | 19 | 14 | 12 | |||||||||
| Orthetrum cancellatum (Linnaeus, 1758) | 3 | 14 | ||||||||||
| Orthetrum coerulescens (Fabricius, 1798) | 34 | 7 | 28 | 33 | ||||||||
| Orthetrum brunneum (Fonscolombe, 1837) | 20 | 28 | ||||||||||
| Sympetrum sanguineum (Müller, 1764) | 1 | 2 | 1 | |||||||||
| Sympetrum fonscolombii (Selys, 1840) | 2 | 2 | ||||||||||
| Sympetrum striolatum (Charpentier, 1840) | 101 | 3 | 1 | |||||||||
| Sympetrum meridionale (Selys, 1841) | 1 | |||||||||||
| Crocothemis erythraea (Brullé, 1832) | 2 | 2 | 4 | 6 |
References
- Steward, A.L.; Von Schiller, D.; Tockner, K.; Marshall, J.C.; Bunn, S.E. When the River Runs Dry: Human and Ecological Values of Dry Riverbeds. Front. Ecol. Environ. 2012, 10, 202–209. [Google Scholar] [CrossRef] [Green Version]
- Leigh, C.; Boulton, A.J.; Courtwright, J.L.; Fritz, K.; May, C.L.; Walker, R.H.; Datry, T. Ecological Research and Management of Intermittent Rivers: An Historical Review and Future Directions. Freshw. Biol. 2015, 61, 1181–1199. [Google Scholar] [CrossRef]
- Larned, S.T.; Datry, T.; Arscott, D.B.; Tockner, K. Emerging Concepts in Temporary-River Ecology. Freshw. Biol. 2010, 55, 717–738. [Google Scholar] [CrossRef]
- Bonada, N.; Rieradevall, M.; Prat, N.; Resh, V.H. Benthic Macroinvertebrate Assemblages and Macrohabitat Connectivity in Mediterranean-Climate Streams of Northern California. J. N. Am. Benthol. Soc. 2006, 25, 32–43. [Google Scholar] [CrossRef]
- Sánchez-Montoya, M.D.M.; Puntí, T.; Suárez, M.L.; Vidal-Abarca, M.D.R.; Rieradevall, M.; Poquet, J.M.; Zamora-Muñoz, C.; Robles, S.; Álvarez, M.; Alba-Tercedor, J.; et al. Concordance between Ecotypes and Macroinvertebrate Assemblages in Mediterranean Streams. Freshw. Biol. 2007, 52, 2240–2255. [Google Scholar] [CrossRef]
- Argyroudi, A.; Chatzinikolaou, Y.; Poirazidis, K.; Lazaridou, M. Do Intermittent and Ephemeral Mediterranean Rivers Belong to the Same River Type? Aquat. Ecol. 2009, 43, 465–476. [Google Scholar]
- Tierno de Figueroa, J.M.; López-Rodríguez, M.J.; Fenoglio, S.; Sánchez-Castillo, P.; Fochetti, R. Freshwater Biodiversity in the Rivers of the Mediterranean Basin. Hydrobiologia 2013, 719, 137–186. [Google Scholar] [CrossRef]
- Zarfl, C.; Lumsdon, A.E.; Berlekamp, J.; Tydecks, L.; Tockner, K. A Global Boom in Hydropower Dam Construction. Aquat. Sci. 2015, 77, 161–170. [Google Scholar] [CrossRef]
- Dudgeon, D. Multiple Threats Imperil Freshwater Biodiversity in the Anthropocene. Curr. Biol. 2019, 29, 960–967. [Google Scholar] [CrossRef]
- Huđek, H.; Žganec, K.; Pusch, M.T. A Review of Hydropower Dams in Southeast Europe—Distribution, Trends and Availability of Monitoring Data Using the Example of a Multinational Danube Catchment Subarea. Renew. Sustain. Energy Rev. 2020, 117, 109434. [Google Scholar] [CrossRef]
- Bouhala, Z.; Khemissa, C.; Márquez-Rodríguez, J.; Ferreras-Romero, M.; Samraoui, F.; Samraoui, B. Ecological correlates of odonate assemblages of a Mediterranean stream, Wadi Cherf, northeastern Algeria: Implications for conservation. Int. J. Odonatol. 2019, 22, 181–197. [Google Scholar] [CrossRef]
- van Vliet, J.; de Groot, H.L.F.; Rietveld, P.; Verburg, P.H. Manifestations and Underlying Drivers of Agricultural Land Use Change in Europe. Landsc. Urban Plan. 2015, 133, 24–36. [Google Scholar] [CrossRef] [Green Version]
- Underwood, E.C.; Viers, J.H.; Klausmeyer, K.R.; Cox, R.L.; Shaw, M.R. Threats and Biodiversity in the Mediterranean Biome. Divers. Distrib. 2009, 15, 188–197. [Google Scholar] [CrossRef]
- Sirami, C.; Nespoulous, A.; Cheylan, J.P.; Marty, P.; Hvenegaard, G.T.; Geniez, P.; Schatz, B.; Martin, J.L. Long-Term Anthropogenic and Ecological Dynamics of a Mediterranean Landscape: Impacts on Multiple Taxa. Landsc. Urban Plan. 2010, 96, 214–223. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeler, R.A.; Mittermeler, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity Hotspots for Conservation Priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Novacek, M.J.; Cleland, E.E. The Current Biodiversity Extinction Event: Scenarios for Mitigation and Recovery. Proc. Natl. Acad. Sci. USA 2001, 98, 5466–5470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tscharntke, T.; Klein, A.M.; Kruess, A.; Steffan-Dewenter, I.; Thies, C. Landscape Perspectives on Agricultural Intensification and Biodiversity—Ecosystem Service Management. Ecol. Lett. 2005, 8, 857–874. [Google Scholar] [CrossRef]
- Tscharntke, T.; Clough, Y.; Wanger, T.C.; Jackson, L.; Motzke, I.; Perfecto, I.; Vandermeer, J.; Whitbread, A. Global Food Security, Biodiversity Conservation and the Future of Agricultural Intensification. Biol. Conserv. 2012, 151, 53–59. [Google Scholar] [CrossRef]
- Allan, J.D. Landscapes and Riverscapes: The Influence of Land Use on Stream Ecosystems. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 257–284. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.B.; Host, G.E. Recent Developments in Landscape Approaches for the Study of Aquatic Ecosystems. J. N. Am. Benthol. Soc. 2010, 29, 41–66. [Google Scholar] [CrossRef] [Green Version]
- Walsh, C.J.; Roy, A.H.; Feminella, J.W.; Cottingham, P.D.; Groffman, P.M.; Morgan, R.P. The Urban Stream Syndrome: Current Knowledge and the Search for a Cure. J. N. Am. Benthol. Soc. 2005, 24, 706–723. [Google Scholar] [CrossRef]
- Marchi, L.; Borga, M.; Preciso, E.; Gaume, E. Characterisation of Selected Extreme Flash Floods in Europe and Implications for Flood Risk Management. J. Hydrol. 2010, 394, 118–133. [Google Scholar] [CrossRef]
- Haslett, J.R.; Berry, P.M.; Bela, G.; Jongman, R.H.G.; Pataki, G.; Samways, M.J.; Zobel, M. Changing Conservation Strategies in Europe: A Framework Integrating Ecosystem Services and Dynamics. Biodivers. Conserv. 2010, 19, 2963–2977. [Google Scholar] [CrossRef]
- Nagy, H.B.; László, Z.; Szabó, F.; Szőcs, L.; Dévai, G.; Tóthmérész, B. Landscape-Scale Terrestrial Factors Are Also Vital in Shaping Odonata Assemblages of Watercourses. Sci. Reports 2019, 9, 18196. [Google Scholar] [CrossRef] [PubMed]
- Wiberg-Larsen, P.; Wiberg-Larsen, P.; Brodersen, K.P.; Birkholm, S.; Grénx, P.N.; Skriver, J. Species Richness and Assemblage Structure of Trichoptera in Danish Streams. Freshw. Biol. 2000, 43, 633–647. [Google Scholar] [CrossRef]
- Brown, B.L. Spatial Heterogeneity Reduces Temporal Variability in Stream Insect Communities. Ecol. Lett. 2003, 6, 316–325. [Google Scholar] [CrossRef]
- Andersson, G.K.S.; Birkhofer, K.; Rundlöf, M.; Smith, H.G. Landscape Heterogeneity and Farming Practice Alter the Species Composition and Taxonomic Breadth of Pollinator Communities. Basic Appl. Ecol. 2013, 14, 540–546. [Google Scholar] [CrossRef]
- Maskell, L.C.; Botham, M.; Henrys, P.; Jarvis, S.; Maxwell, D.; Robinson, D.A.; Rowland, C.S.; Siriwardena, G.; Smart, S.; Skates, J.; et al. Exploring Relationships between Land Use Intensity, Habitat Heterogeneity and Biodiversity to Identify and Monitor Areas of High Nature Value Farming. Biol. Conserv. 2019, 231, 30–38. [Google Scholar] [CrossRef]
- Frainer, A.; Polvi, L.E.; Jansson, R.; McKie, B.G. Enhanced Ecosystem Functioning Following Stream Restoration: The Roles of Habitat Heterogeneity and Invertebrate Species Traits. J. Appl. Ecol. 2018, 55, 377–385. [Google Scholar] [CrossRef]
- Laudon, H.; Kuglerová, L.; Sponseller, R.A.; Futter, M.; Nordin, A.; Bishop, K.; Lundmark, T.; Egnell, G.; Ågren, A.M. The Role of Biogeochemical Hotspots, Landscape Heterogeneity, and Hydrological Connectivity for Minimizing Forestry Effects on Water Quality. Ambio 2016, 45, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Soininen, J.; Bartels, P.; Heino, J.; Luoto, M.; Hillebrand, H. Toward More Integrated Ecosystem Research in Aquatic and Terrestrial Environments. Bioscience 2015, 65, 174–182. [Google Scholar] [CrossRef] [Green Version]
- Sahlén, G.; Ekestubbe, K. Identification of Dragonflies (Odonata) as Indicators of General Species Richness in Boreal Forest Lakes. Biodivers. Conserv. 2001, 10, 673–690. [Google Scholar] [CrossRef]
- Simaika, J.P.; Samways, M.J. An Easy-to-Use Index of Ecological Integrity for Prioritizing Freshwater Sites and for Assessing Habitat Quality. Biodivers. Conserv. 2009, 18, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Kutcher, T.E.; Bried, J.T. Adult Odonata Conservatism as an Indicator of Freshwater Wetland Condition. Ecol. Indic. 2014, 38, 31–39. [Google Scholar] [CrossRef]
- Corbet, P.S. Dragonflies. Behaviour and Ecology of Odonata; Harley Books: Colchester, UK, 1999. [Google Scholar]
- World Odonata List. Available online: https://www2.pugetsound.edu/academics/academic-resources/slater-museum/biodiversity-resources/dragonflies/world-odonata-list2/ (accessed on 26 May 2022).
- Boudot, J.P.; Kalkman, V.J. Atlas of the European Dragonflies and Damselflies; KNNV Uitgeverij: Zeist, The Netherlands, 2015. [Google Scholar]
- Buchwald, R. Vegetation and Dragonfly Fauna—Characteristics and Examples of Biocenological Field Studies. Vegetatio 1992, 101, 99–107. [Google Scholar] [CrossRef]
- Steytler, N.S.; Samways, M.J. Biotope Selection by Adult Male Dragonflies (Odonata) at an Artificial Lake Created for Insect Conservation in South Africa. Biol. Conserv. 1995, 72, 381–386. [Google Scholar] [CrossRef]
- Corbet, P.; Brooks, S. Dragonflies (Collins New Naturalist Library, Book 106); Collins: London, UK, 2008. [Google Scholar]
- Vilenica, M. Ecological Traits of Dragonfly (Odonata) Assemblages along an Oligotrophic Dinaric Karst Hydrosystem. Ann. Limnol.-Int. J. Limnol. 2017, 53, 377–389. [Google Scholar] [CrossRef]
- Vilenica, M.; Rebrina, F.; Matoničkin Kepčija, R.; Šegota, V.; Rumišek, M.; Ružanović, L.; Brigić, A. Aquatic Macrophyte Vegetation Promotes Taxonomic and Functional Diversity of Odonata Assemblages in Intermittent Karst Rivers in the Mediterranean. Diversity 2022, 14, 31. [Google Scholar] [CrossRef]
- Bried, J.T.; Herman, B.D.; Ervin, G.N. Umbrella Potential of Plants and Dragonflies for Wetland Conservation: A Quantitative Case Study Using the Umbrella Index. J. Appl. Ecol. 2007, 44, 833–842. [Google Scholar] [CrossRef]
- Korbaa, M.; Ferreras-Romero, M.; Ruiz-Garcia, A.; Boumaiza, M. TSOI—A New Index Based on Odonata Populations to Assess the Conservation Relevance of Watercourses in Tunisia. Odonatologica 2018, 47, 43–47. [Google Scholar]
- Khelifa, R. Sensitivity of Biodiversity Indices to Life History Stage, Habitat Type and Landscape in Odonata Community. Biol. Conserv. 2019, 237, 63–69. [Google Scholar] [CrossRef]
- Salinas, M.J.; Casas, J.J. Riparian Vegetation of Two Semi-Arid Mediterranean Rivers: Basin-Scale Responses of Woody and Herbaceous Plants to Environmental Gradients. Wetlands 2007, 27, 831–845. [Google Scholar] [CrossRef]
- Larsen, S.; Sorace, A.; Mancini, L. Riparian Bird Communities as Indicators of Human Impacts along Mediterranean Streams. Environ. Manag. 2010, 45, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Eken, G.; Bennun, L.; Brooks, T.M.; Darwall, W.; Fishpool, L.D.C.; Foster, M.; Knox, D.; Langhammer, P.; Matiku, P.; Radford, E.; et al. Key Biodiversity Areas as Site Conservation Targets. Bioscience 2004, 54, 1110–1118. [Google Scholar] [CrossRef] [Green Version]
- Hochkirch, A.; Samways, M.J.; Gerlach, J.; Böhm, M.; Williams, P.; Cardoso, P.; Cumberlidge, N.; Stephenson, P.J.; Seddon, M.B.; Clausnitzer, V.; et al. A Strategy for the next Decade to Address Data Deficiency in Neglected Biodiversity. Conserv. Biol. 2021, 35, 502–509. [Google Scholar] [CrossRef]
- Illies, J. Limnofauna Europaea, 2nd ed.; Gustav Fischer Verlag: Stuttgart, Germany, 1978. [Google Scholar]
- Bonacci, O.; Željković, I.; Galić, A. Karst Rivers’ Particularity: An Example from Dinaric Karst (Croatia/Bosnia and Herzegovina). Environ. Earth Sci. 2013, 70, 963–974. [Google Scholar] [CrossRef]
- Šegota, T.; Filipčić, A. Köppen’s Classification of Climates and the Problem of Corresponding Croatian Terminology. Geoadria 2003, 8, 17–37. [Google Scholar] [CrossRef] [Green Version]
- Zaninović, K.; Gajić-Čapka, M.; Perčec Tadić, M.; Vučetić, M.; Milković, J.; Bajić, A.; Cindrić, K.; Cvitan, L.; Katušin, Z.; Kaučić, D.; et al. Climate Atlas of Croatia 1961–1990, 1971–2000; Zaninović, K., Ed.; Meteorological and Hydrological Service of Croatia: Zagreb, Croatia, 2008.
- Dijkstra, K.-D.B.; Schröter, A. Field Guide to the Dragonflies of Britain and Europe; Bloomsbury Publishing: London, UK, 2020. [Google Scholar]
- Pearce-Higgins, J.W.; Chandler, D. Do surveys of adult dragonflies and damselflies yield repeatable data? Variation in monthly counts of abundance and species richness. J. Insect Conserv. 2020, 24, 877–889. [Google Scholar] [CrossRef]
- Tews, J.; Brose, U.; Grimm, V.; Tielbörger, K.; Wichmann, M.C.; Schwager, M.; Jeltsch, F. Animal Species Diversity Driven by Habitat Heterogeneity/Diversity: The Importance of Keystone Structures. J. Biogeogr. 2004, 31, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Kosztra, B.; Büttner, G.; Hazeu, G.; Arnold, S. Updated CLC Illustrated Nomenclature Guidelines; European Topic Centre on Urban, Land and Soil Systems (ETC/ULS) Environment Agency Austria: Vienna, Austria, 2017.
- Büttner, G.; Kosztra, B.; Maucha, G.; Pataki, R.; Kleeschulte, S.; Hazeu, G.; Vittek, M.; Littkopf, A. Copernicus Land Monitoring Service, CORINE Land Cover User Manual; European Environment Agency: Copenhagen, Denmark, 2021.
- Zagorskas, J. GIS-Based Modelling and Estimation of Land Use Mix in Urban Environment. Int. J. Environ. Sci. 2016, 1, 284–293. [Google Scholar]
- Jiao, J.; Rollo, J.; Fu, B. The Hidden Characteristics of Land-Use Mix Indices: An Overview and Validity Analysis Based on the Land Use in Melbourne, Australia. Sustainability 2021, 13, 1898. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer V6: User Manual/Tutorial; Plymouth: Auckland, New Zealand, 2006. [Google Scholar]
- Ricotta, C. On Parametric Diversity Indices in Ecology: A Historical Note. Community Ecol. 2005, 6, 241–244. [Google Scholar] [CrossRef] [Green Version]
- Tóthmérész, B. On the Characterization of Scale-Dependent Diversity. Abstr. Bot. 1998, 22, 149–156. [Google Scholar]
- Kindt, R.; Coe, R. Tree Diversity Analysis: A Manual and Software for Common Statistical Methods for Ecological and Biodiversity Studies; World Agroforestry Centre: Nairobi, Kenya, 2005. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Ellingsen, K.E.; Clarke, K.R.; Somerfield, P.J.; Warwick, R.M. Taxonomic Distinctness as a Measure of Diversity Applied over a Large Scale: The Benthos of the Norwegian Continental Shelf. J. Anim. Ecol. 2005, 74, 1069–1079. [Google Scholar] [CrossRef]
- Freitag, S.; Van Jaarsveld, A.S. Relative Occupancy, Endemism, Taxonomic Distinctiveness and Vulnerability: Prioritizing Regional Conservation Actions. Biodivers. Conserv. 1997, 6, 211–232. [Google Scholar] [CrossRef]
- White, E.; Hunt, P.; Schlesinger, M.; Corser, J.; deMaynadier, P. A Conservation Status Assessment of Odonata for the Northeastern United States; New York Natural Heritage Program: Albany, NY, USA, 2014. [Google Scholar]
- Samways, M.J.; Taylor, S. Impacts of Invasive Alien Plants on Red-Listed South African Dragonflies (Odonata). S. Afr. J. Sci. 2004, 100, 78–81. [Google Scholar]
- Harabiš, F.; Dolný, A. Ecological Factors Determining the Density-Distribution of Central European Dragonflies (Odonata). Eur. J. Entomol. 2010, 107, 571–577. [Google Scholar] [CrossRef] [Green Version]
- Petrovičová, K.; Langraf, V.; David, S.; Krumpálová, Z.; Schlarmannová, J. Distinct Odonata Assemblage Variations in Lentic Reservoirs in Slovakia (Central Europe). Biologia 2021, 76, 3727–3741. [Google Scholar] [CrossRef]
- Kotarac, M.; Šalamun, A.; Vilenica, M. EU Natura 2000 Integration Project: Field Research and Laboratory Processing for Collecting New Inventory Data for Taxonomic Groups: Actinopterygii and Cephalaspidomorphi, Amphibia and Reptilia, Aves, Chiroptera, Decapoda, Lepidoptera, Odonata, Plecoptera–Final Report for the Taxonomic Group Odonata; Ministry of Environmental and Nature Protection: Zagreb, Croatia, 2016.
- Belančić, A.; Bogdanović, T.; Franković, M.; Ljuština, M.; Mihoković, N.; Vitas, B. Red Data Book of Dragonflies of Croatia; Franković, M., Ed.; Ministry of Culture, State Institute for Nature Protection, Republic of Croatia: Zagreb, Croatia, 2008.
- IBM Corp. IBM SPSS Statistics for Windows, Version 27.0; IBM: Armonk, NY, USA, 2020.
- Ollero, A. Channel Adjustments, Floodplain Changes and Riparian Ecosystems of the Middle Ebro River: Assessment and Management. Int. J. Water Resour. Dev. 2007, 23, 73–90. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Dick, D.A. Spatial Heterogeneity of Soil Nutrients and Plant Species in Herb-Dominated Communities of Contrasting Land Use. Plant Ecol. 2010, 209, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, W.H.; Reckhow, K.H.; Bernhardt, E.S. Global Change: The Nitrogen Cycle and Rivers. Water Resour. Res. 2006, 42. [Google Scholar] [CrossRef] [Green Version]
- Freshwaterecology. Available online: https://www.freshwaterecology.info/fwe_search.php?og=mzb (accessed on 17 May 2022).
- Buffagni, A.; Erba, S.; Armanini, D.G. The Lentic-Lotic Character of Mediterranean Rivers and Its Importance to Aquatic Invertebrate Communities. Aquat. Sci. 2010, 72, 45–60. [Google Scholar] [CrossRef]
- Pitlo, R.H.; Dawson, F.H. Flow Resistance by Aquatic Weeds. In Aquatic Weeds: The Ecology and Management of Nuisance Aquatic Vegetation; Pieterse, A.H., Murphy, K.J., Eds.; Oxford University Press: Oxford, UK, 1990; pp. 74–84. [Google Scholar]
- Perron, M.A.C.; Pick, F.R. Stormwater Ponds as Habitat for Odonata in Urban Areas: The Importance of Obligate Wetland Plant Species. Biodivers. Conserv. 2020, 29, 913–931. [Google Scholar] [CrossRef]
- Purse, B.V.; Thompson, D.J. Oviposition Site Selection by Coenagrion Mercuriale (Odonata: Coenagrionidae). Int. J. Odonatol. 2009, 12, 257–273. [Google Scholar] [CrossRef]
- Brito, J.S.; Michelan, T.S.; Juen, L. Aquatic Macrophytes Are Important Substrates for Libellulidae (Odonata) Larvae and Adults. Limnology 2021, 22, 139–149. [Google Scholar] [CrossRef]
- Hardersen, S. Dragonfly (Odonata) Communities at Three Lotic Sites with Different Hydrological Characteristics. Ital. J. Zool. 2008, 75, 271–283. [Google Scholar] [CrossRef]
- Belmar, O.; Velasco, J.; Gutiérrez-Cánovas, C.; Mellado-Díaz, A.; Millán, A.; Wood, P.J. The Influence of Natural Flow Regimes on Macroinvertebrate Assemblages in a Semiarid Mediterranean Basin. Ecohydrology 2012, 6, 363–379. [Google Scholar] [CrossRef] [Green Version]
- Yalles Satha, A.; Samraoui, B. Environmental Factors Influencing Odonata Communities of Three Mediterranean Rivers: Kebir-East, Seybouse, and Rhumel Wadis, Northeastern Algeria. Rev. Ecol. 2017, 72, 314–329. [Google Scholar]
- Magnusson, A.K.; Williams, D.D. The Roles of Natural Temporal and Spatial Variation versus Biotic Influences in Shaping the Physicochemical Environment of Intermittent Ponds: A Case Study. Arch. Hydrobiol. 2006, 165, 537–556. [Google Scholar] [CrossRef]
- Stoks, R.; McPeek, M.A. Predators and Life Histories Shape Lestes Damselfly Assemblages along a Freshwater Habitat Gradient. Ecology 2003, 84, 1576–1587. [Google Scholar] [CrossRef]
- Johansson, F.; Suhling, F. Behaviour and Growth of Dragonfly Larvae along a Permanent to Temporary Water Habitat Gradient. Ecol. Entomol. 2004, 29, 196–202. [Google Scholar] [CrossRef]
- Suhling, F.; Sahlén, G.; Gorb, S.; Kalkman, V.J.; Dijkstra, K.D.B.; van Tol, J. Order Odonata. In Thorp and Covich’s Freshwater Invertebrates: Ecology and General Biology, 4th ed.; Thorp, J.H., Rogers, D.C., Eds.; Academic Press: Cambridge, MA, USA, 2015; Volume 1, pp. 893–932. [Google Scholar]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; Conze, K.-J.; De Knijf, G.; Dyatlova, E.; Ferreira, S.; Jović, M.; Ott, J.; Riservato, E.; et al. European Red List of Dragonflies; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Cordulegaster Heros. The IUCN Red List of Threatened Species 2020. Available online: https://www.iucnredlist.org/species/158700/140520373 (accessed on 26 May 2022). [CrossRef]
- Pernecker, B.; Mauchart, P.; Csabai, Z. What to Do If Streams Go Dry? Behaviour of Balkan Goldenring (Cordulegaster Heros, Odonata) Larvae in a Simulated Drought Experiment in SW Hungary. Ecol. Entomol. 2020, 45, 1457–1465. [Google Scholar] [CrossRef]
- Negro, M.; Casale, A.; Migliore, L.; Palestrini, C.; Rolando, A. The Effect of Local Anthropogenic Habitat Heterogeneity on Assemblages of Carabids (Coleoptera, Caraboidea) Endemic to the Alps. Biodivers. Conserv. 2007, 16, 3919–3932. [Google Scholar] [CrossRef]
- Vilenica, M.; Alegro, A.; Koletić, N.; Mihaljević, Z. New Evidence of Lindenia tetraphylla (Vander Linden, 1825) (Odonata, Gomphidae) Reproduction at the North-Western Border of Its Distribution. Nat. Croat. Period. Musei Hist. Nat. Croat. 2016, 25, 287–294. [Google Scholar] [CrossRef] [Green Version]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilenica, M.; Rebrina, F.; Ružanović, L.; Gulin, V.; Brigić, A. Odonata Assemblages as a Tool to Assess the Conservation Value of Intermittent Rivers in the Mediterranean. Insects 2022, 13, 584. https://doi.org/10.3390/insects13070584
Vilenica M, Rebrina F, Ružanović L, Gulin V, Brigić A. Odonata Assemblages as a Tool to Assess the Conservation Value of Intermittent Rivers in the Mediterranean. Insects. 2022; 13(7):584. https://doi.org/10.3390/insects13070584
Chicago/Turabian StyleVilenica, Marina, Fran Rebrina, Lea Ružanović, Vesna Gulin, and Andreja Brigić. 2022. "Odonata Assemblages as a Tool to Assess the Conservation Value of Intermittent Rivers in the Mediterranean" Insects 13, no. 7: 584. https://doi.org/10.3390/insects13070584