
Biodiversity and Distribution of Reticulitermes in the Southeastern USA

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
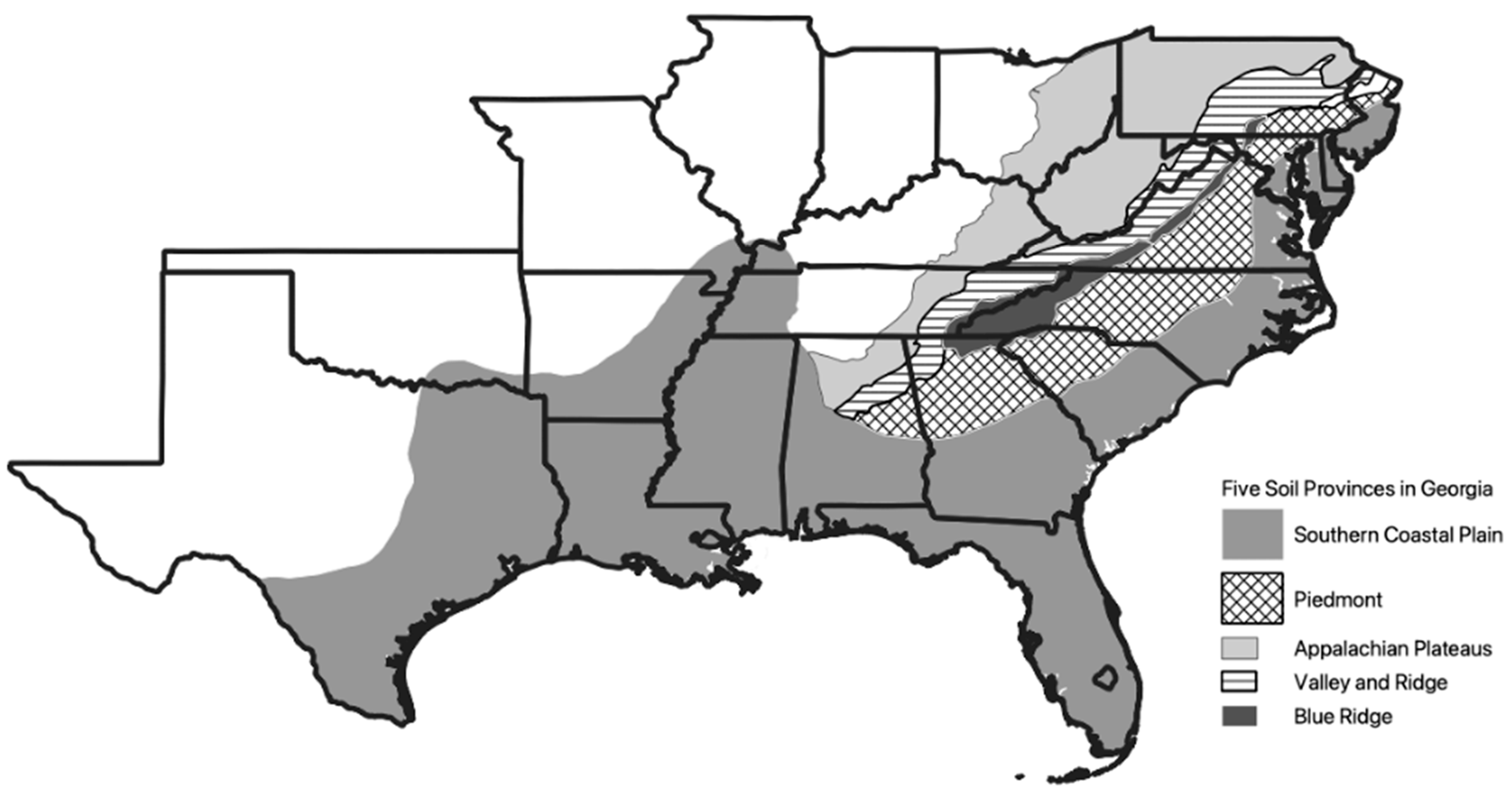
2.1. Sample Area
2.2. Sample Collection
2.3. Sample Processing
2.4. Sample Identification
2.5. GIS Visualization
2.6. Spatial Data Analysis
3. Results
3.1. Survey
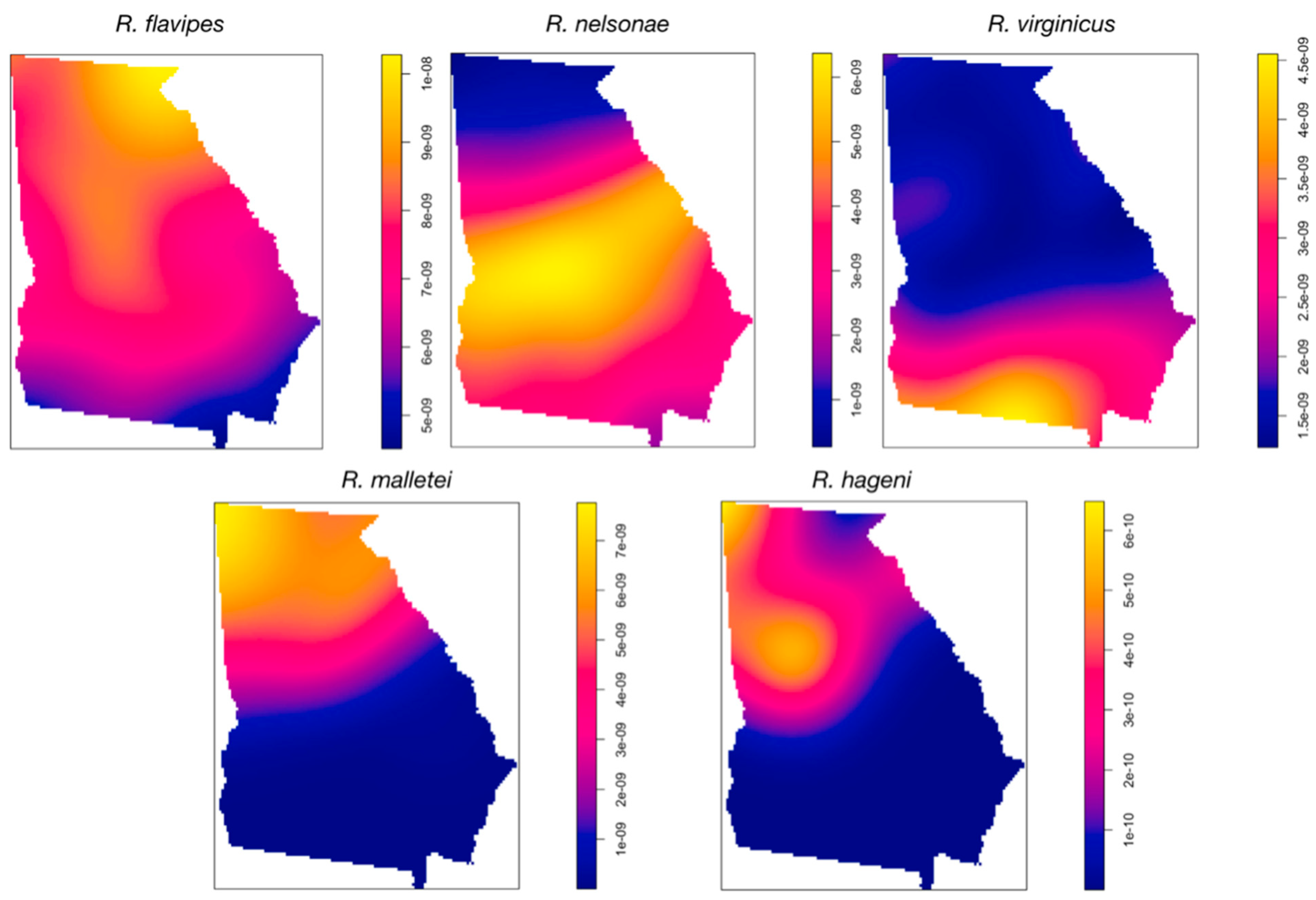
3.2. Spatial Data Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engel, M.S.; Ceríaco, L.M.; Daniel, G.M.; Dellapé, P.M.; Löbl, I.; Marinov, M.; Reis, R.E.; Young, M.T.; Dubois, A.; Agarwal, I.; et al. The taxonomic impediment: A shortage of taxonomists, not the lack of technical approaches. Zool. J. Linn. Soc. 2021, 193, 381–387. [Google Scholar] [CrossRef]
- Lobl, I. Assessing biotic diversity: The glorious past, present, and the uncertain future. Bull. Entomol. Soc. Malta 2018, 10, 5–15. [Google Scholar]
- Sharkey, M.J.; Janzen, D.H.; Hallwachs, W.; Chapman, E.G.; Smith, M.A.; Dapkey, T.; Brown, A.; Ratnasingham, S.; Naik, S.; Manjunath, R.; et al. Minimalist revision and description of 403 new species in 11 subfamilies of costa Rican braconid parasitoid wasps, including host records for 219 species. ZooKeys 2021, 1013, 1–665. [Google Scholar] [CrossRef] [PubMed]
- Cardenas, C.R.; Luo, A.R.; Jones, T.H.; Schultz, T.R.; Adams, R.M.M. Using an integrative taxonomic approach to delimit a sibling species, Mycetomoellerius Mikromelanos Sp. Nov. (Formicidae: Attini: Attina). PeerJ 2021, 9, e11622. [Google Scholar] [CrossRef] [PubMed]
- Kjærandsen, J. Current state of DNA barcoding of sciaroidea (Diptera)—Highlighting the need to build the reference library. Insects 2022, 13, 147. [Google Scholar] [CrossRef]
- Petrović, A. Sizing the knowledge gap in taxonomy: The last dozen years of aphidiinae research. Insects 2022, 13, 170. [Google Scholar] [CrossRef]
- Dubois, A.; Ohler, A.; Pyron, R.A. New concepts and methods for phylogenetic taxonomy and nomenclature in zoology, exemplified by a new ranked cladonomy of recent amphibians (Lissamphibia). Megataxa 2021, 5, 1–738. [Google Scholar] [CrossRef]
- Ride, W.D.L. International Commission on Zoological Nomenclature, 4th ed.; International Union of Biological Sciences; Natural History Museum: London, UK, 1999. [Google Scholar]
- Wagner, D.L.; Grames, E.M.; Forister, M.L.; Berenbaum, M.R.; Stopak, D. Insect decline in the Anthropocene: Death by a thousand cuts. Proc. Natl. Acad. Sci. USA 2021, 118, e2023989118. [Google Scholar] [CrossRef]
- Saunders, M.E.; Janes, J.K.; O’Hanlon, J.C. Moving on from the insect apocalypse narrative: Engaging with evidence-based insect conservation. BioScience 2020, 70, 80–89. [Google Scholar] [CrossRef] [Green Version]
- Mallet, J.; Willmott, K. Taxonomy: Renaissance or Tower of Babel? Trends Ecol. Evol. 2003, 18, 57–59. [Google Scholar] [CrossRef]
- Rust, M.K.; Su, N.-Y. Managing social insects of urban importance. Annu. Rev. Entomol. 2012, 57, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.; Wagner, T.L.; Mulrooney, J.E.; Shelton, T.G. Subterranean termites—Their prevention and control in buildings. U. S. Dep. Agric. For. Serv. Home Gard. Bull. 2006, 64, 38. [Google Scholar]
- Myer, A.; Forschler, B.T. Evidence for the role of subterranean termites (Reticulitermes spp.) in temperate forest soil nutrient cycling. Ecosystems 2018, 22, 602–618. [Google Scholar] [CrossRef]
- Jouquet, P.; Bottinelli, N.; Shanbhag, R.R.; Bourguignon, T.; Traoré, S.; Abbasi, S.A. Termites: The neglected soil engineers of tropical soils. Soil Sci. 2016, 181, 157–165. [Google Scholar] [CrossRef]
- International Code Council. International Building Code; International Code Council: Falls Church, VA, USA, 2000.
- ORKIN. Termite Infestation Probability Zones Map (TIP). Available online: https://www.orkin.com/pests/termites/termite-infestation-probability-zones-map (accessed on 30 May 2022).
- What Are the Worst U.S. States for Termites? Available online: https://www.groundworks.com/resources/worst-us-states-for-termites/ (accessed on 30 May 2022).
- Clément, J.-L.; Howard, R.W.; Blum, M.S.; Lloyd, H. L’isolement spécifique des termites du genre Reticulitermes (Isoptera) du sudest des Etats-Unis. Mise en èvidence grâce à la chimie et au comportement d’une espèce jumelle de R. virginicus = R. malletei sp. nov. et d’une semi-species de R. flavipes. Comptes Rendus Académie Sci. 1986, 302, 67–70. [Google Scholar]
- Lim, S.Y.; Forschler, B.T. A new species of Reticulitermes Holmgren, 1913 (Isoptera: Rhinotermitidae) from the Southeastern United States. Insecta Mundi 2012, 0252, 1–4. [Google Scholar]
- Nelson, L.J.; Cool, L.G.; Solek, C.W.; Haverty, M.I. Cuticular hydrocarbons and soldier defense secretions of Reticulitermes in Southern California: A critical analysis of the taxonomy of the genus in North America. J. Chem. Ecol. 2008, 34, 1452–1475. [Google Scholar] [CrossRef]
- Weesner, F.M. Termites of the Nearctic Region. In Biology of Termites, 1st ed.; Krishna, K., Weesner, F.M., Eds.; Academic Press: New York, NY, USA, 1970; Volume 2, pp. 477–525. [Google Scholar]
- Banks, F.A. Species Distinction in Reticulitermes (Isoptera: Rhinotermitidae). Master’s Thesis, University of Chicago, Chicago, IL, USA, 1946. [Google Scholar]
- Miller, E.M. Florida Termites: A Handbook; University of Miami Press: Coral Gables, FL, USA, 1949; pp. 1–30. [Google Scholar]
- Nutting, W.L. Insecta: Isotopera. In Soil Biology Guide, 1st ed.; Dindal, D.L., Ed.; John Wiley & Sons: New York, NY, USA, 1990; pp. 997–1032. [Google Scholar]
- Hostettler, N.C.; Hall, D.W.; Scheffrahn, R.H. Intracolony morphometric variation and labral shape in Florida Reticulitermes (Isoptera: Rhinotermitidae) soldiers: Significance for identification. Fla. Entomol. 1995, 78, 119–129. [Google Scholar] [CrossRef]
- Lim, S.Y.; Forschler, B.T. Reticulitermes nelsonae, a new species of subterranean termite (Rhinotermitidae) from the Southeastern United States. Insects 2012, 3, 62–90. [Google Scholar] [CrossRef] [Green Version]
- Copren, K.A.; Nelson, L.J.; Vargo, E.L.; Haverty, M.I. Phylogenetic analyses of MTDNA sequences corroborate taxonomic designations based on cuticular hydrocarbons in subterranean termites. Mol. Phylogenet. Evol. 2005, 35, 689–700. [Google Scholar] [CrossRef]
- Cameron, S.L.; Whiting, M.F. Mitochondrial genomic comparisons of the subterranean termites from the genus Reticulitermes (Insecta: Isoptera: Rhinotermitidae). Genome 2007, 50, 188–202. [Google Scholar] [CrossRef] [PubMed]
- Austin, J.W.; Szalanski, A.L.; Uva, P.; Kence, A. A comparative genetic analysis of the subterranean termite genus Reticulitermes (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am. 2002, 95, 753–760. [Google Scholar] [CrossRef]
- Will, K.W.; Mishler, B.D.; Wheeler, Q.D. The perils of DNA barcoding and the need for integrative taxonomy. Syst. Biol. 2005, 54, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Lumely, L.; Sperling, F.A.X. Integrating morphology and mitochondrial DNA for species delimitation within the spruce budworm (Choristoneura fumiferana) cryptic species complex (Lepidoptera: Tortricidae). Syst. Entomol. 2010, 35, 416–428. [Google Scholar] [CrossRef]
- United States Department of Agriculture, Natural Resources Conservation Service. Land Resource Regions and Major Land Resource Areas of the United States, the Caribbean, and the Pacific Basin. In U.S. Department of Agriculture Handbook 296; USDA: Washington, DC, USA, 2006. [Google Scholar]
- Jonathan, W.M. Digital Atlas of State of Georgia, Elevation; Digital Data Series; US Geological Survey: Atlanta, GA, USA, 1996.
- McNab, H.W.; Avers, P.E. Ecological Subregions of the United States. Southern Region and Southeastern Forest Experiment Station; USDA: Washington, DC, USA, 1996; Volume 21.
- Gore, P.J.W.; Witherspoon, W. Introduction. In Roadside Geology of Georgia, 1st ed.; Mountain Press Publishing Company: Missoula, MT, USA, 2013; pp. 2–21. [Google Scholar]
- Liu, H.; Beckenbach, A.T. Evolution of the mitochondrial cytochrome oxidase II gene among 10 orders of insects. Mol. Phylogenet. Evol. 1992, 41, 41–52. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-Tree: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef]
- Lewin-Koh, N.; Bivand, R.; Pebesma, E.; Archer, E.; Baddeley, A.; Bibiko, H.-J.; Dray, S.; Forrest, D.; Friendly, M.; Giraudoux, P. maptools: Tools for Reading and Handling Spatial Objects, R Package Version 0.8-27. 2011. Available online: https://scholar.google.com/citations?view_op=view_citation&hl=en&user=f151hGIAAAAJ&citation_for_view=f151hGIAAAAJ:4vMrXwiscB8C (accessed on 30 May 2022).
- Keitt, T.H.; Bivand, R.; Pebesma, E.; Rowlingson, B. rgdal: Bindings for the Geospatial Data Abstraction Library; R Package Version 0.6-27. 2022. Available online: https://cran.r-project.org/web/packages/rgdal/rgdal.pdf (accessed on 30 May 2022).
- Pebesma, E. Simple features for R: Standardized support for spatial vector data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Pebesma, E.J.; Bivand, R.S. Classes and methods for spatial data in R. R News 2005, 5, 9–13. [Google Scholar]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Patterns: Methodology and Applications with R, 1st ed.; Chapman and Hall, CRC Press: London, UK, 2015. [Google Scholar]
- Hill, M.O. Diversity and eveness: A unifying notation and its consequences. Ecology 1972, 54, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Chiu, C.-H.; Jost, L. Unifying species diversity, phylogenetic diversity, functional diversity, and related similarity and differentiation measures through hill numbers. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 297–324. [Google Scholar] [CrossRef] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; The University of Illinois Press: Urbana, IL, USA, 1949; p. 117. [Google Scholar]
- Simpson, E. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Vetter, R.S.; Hinkle, N.C.; Ames, L.M. Distribution of the brown recluse spider (Araneae: Sicariidae) in Georgia with comparison to poison center reports of envenomations. J. Med. Entomol. 2009, 46, 15–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargo, E.L.; Juba, T.R.; Deheer, C.J. Relative abundance and comparative breeding structure of subterranean termite colonies (Reticulitermes flavipes, Reticulitermes hageni, Reticulitermes virginicus, and Coptotermes formosanus) in a South Carolina Lowcountry Site as revealed by molecular markers. Ann. Entomol. Soc. Am. 2006, 99, 1101–1109. [Google Scholar] [CrossRef]
- Janowiecki, M.A.; Austin, J.W.; Szalanski, A.W.; Vargo, E. Identification of Reticulitermes subterranean termites (Blattodea: Rhinotermitidae) in the Eastern United States using inter-simple sequence repeats. J. Econ. Entomol. 2021, 114, 1242–1248. [Google Scholar] [CrossRef]
- Su, N.-Y.; Ye, W.; Ripa, R.; Scheffrahn, R.H.; Giblin-Davis, R.M. Identification of Chilean Reticulitermes (Isoptera: Rhinotermitidae) inferred from three mitochondrial gene DNA sequences and soldier morphology. Ann. Entomol. Soc. Am. 2006, 99, 352–363. [Google Scholar] [CrossRef]
- Chouvenc, T.; Scheffrahn, R.H.; Buss, L. Termite species distribution in Florida and UF termite identification services. EDIS 2022, 2022, 3. [Google Scholar] [CrossRef]
- Su, N.-Y.; Scheffrahn, R.H.; Cabrera, B. Native subterranean termites: Reticulitermes flavipes (Kollar), Reticulitermes virginicus (Banks), Reticulitermes hageni banks (Insecta: Isoptera: Rhinotermitidae). EDIS 2004, 2004, 5. [Google Scholar] [CrossRef]
- University of Florida, Institute of Food and Agricultural Sciences—UF/IFAS. Termite Distribution—Fort Lauderdale Research and Education Center. Available online: https://flrec.ifas.ufl.edu/termites-in-florida/termite-distribution/ (accessed on 30 May 2022).
- Banks, N.; Snyder, T.E. A revision of the Nearctic termites. Bull. United States Natl. Mus. 1920, 108, 288. [Google Scholar] [CrossRef]
- Smithsonian National Museum of Natural History. Entomology Collections Search. Available online: http://n2t.net/ark:/65665/3e999984f-7dbc-4a21-bc9b-348732688df1 (accessed on 31 May 2022).
- Dedeine, F.; Dupont, S.; Guyot, S.; Matsuura, K.; Wang, C.; Habibpour, B.; Bagnères, A.G.; Mantovani, B.; Luchetti, A. Historical biogeography of Reticulitermes termites (Isoptera: Rhinotermitidae) inferred from analyses of mitochondrial and nuclear loci. Mol. Phylogenet. Evol. 2016, 94, 778–790. [Google Scholar] [CrossRef]
- Garrick, R.C.; Collins, B.D.; Yi, R.N.; Dyer, R.J.; Hyseni, C. Identification of Eastern United States Reticulitermes termite species via PCR-RFLP, assessed using training and test data. Insects 2015, 6, 524–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Austin, J.; Bagnères, A.G.; Szalanski, A.; Scheffrahn, R.; Heintschel, B.P.; Messenger, M.; Clément, J.-L.; Gold, R. Reticulitermes malletei (Isoptera: Rhinotermitidae): A valid Nearctic subterranean termite from Eastern North America. Zootaxa 2007, 1554. [Google Scholar] [CrossRef]
- Whitney-King, S.; Austin, J.W.; Szalanski, A.L. Use of soldier pronotal width and mitochondrial DNA sequencing to distinguish the subterranean termites, Reticulitermes flavipes (Kollar) and R. virginicus (Banks) (Isoptera: Rhinotermitidae) on the Delmarva peninsula Delaware, Maryland, and Virginia, USA. Entomol. News 2007, 118, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Stephens, C. Alabama Rhinotermitidae: Nomenclature, Identification, Survey and Phenology. Master’s Thesis, Auburn University, Auburn, AL, USA, 2012. [Google Scholar]
- Scheffrahn, R.; Su, N.Y. Keys to soldier and winged adult termites (Isoptera) of Florida. Fla. Entomol. 1994, 77, 460–474. [Google Scholar] [CrossRef]
- Messenger, M.T. New Orleans Mosquito & Termite Control Board Bulletin, 2nd ed.; No. 01; New Orleans Mosquito and Termite Control Board: New Orleans, LA, USA, 2002; Volume 2, pp. 1–12.
- Szalanski, A.L.; Austin, J.W.; Owens, C.B. Identification of Reticulitermes spp. (Isoptera: Reticulitermatidae) from South Central United States by PCR-RFLP. J. Econ. Entomol. 2003, 96, 1514–1519. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X.; Jia, G.; Deng, J.; Wang, C.; Wang, Q.; et al. Assessment of four DNA fragments (COI, 16S rdna, ITS2, 12s rdna) for species identification of the IXODIDA (Acari: Ixodida). Parasites Vectors 2014, 7, 93. [Google Scholar] [CrossRef] [Green Version]
- Ruihua, Z.; Ping, J.; Chuanbo, S.; Deyong, S.; Feng, Z.; Chaochao, H. The analysis of genetic variation in the mitochondrial genome and its application for the identification of Papilio species. Mitochondr. DNA Part B 2018, 3, 687–690. [Google Scholar] [CrossRef] [Green Version]
- Jörger, K.M.; Schrödl, M. How to describe a cryptic species? Practical challenges of molecular taxonomy. Front. Zool. 2013, 10, 59. [Google Scholar] [CrossRef] [Green Version]
- Goulding, T.C.; Dayrat, B. Integrative taxonomy: Ten years of practice and looking into the future. Arch. Zool. Mus. Lomonosov Mosc. State Univ. 2016, 54, 116–133. [Google Scholar]
- Yeates, D.; Seago, A.; Nelson, L.; Cameron, S.; Joseph, L.; Trueman, J. Integrative taxonomy, or iterative taxonomy? Syst. Entomol. 2010, 36, 209–217. [Google Scholar] [CrossRef]
- Bourguignon, T.; Lo, N.; Šobotník, J.; Sillam-Dussès, D.; Roisin, Y.; Evans, T.A. Oceanic dispersal, vicariance and human introduction shaped the modern distribution of the termites Reticulitermes, Heterotermes and Coptotermes. Proc. R. Soc. B Boil. Sci. 2016, 283, 20160179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachos, F.E. (New) species concepts, species delimitation and the inherent limitations of taxonomy. J. Genet. 2018, 97, 811–815. [Google Scholar] [CrossRef]
- Gross, M. Barcoding biodiversity. Curr. Biol. 2012, 22, R73–R76. [Google Scholar] [CrossRef] [Green Version]
- Ghiselli, F.; Milani, L. Linking the mitochondrial genotype to phenotype: A complex endeavour. Philos. Trans. R. Soc. B 2019, 375, 20190169. [Google Scholar] [CrossRef] [Green Version]
- Chan, A.H.; Chaisiri, K.; Saralamba, S.; Morand, S.; Thaenkham, U. Assessing the suitability of mitochondrial and nuclear DNA genetic markers for molecular systematics and species identification of helminths. Parasites Vectors 2021, 14, 233. [Google Scholar] [CrossRef]
- Austin, J.W.; Szalanski, A.L.; Messenger, M.T. Mitochondrial DNA variation and distribution of the subterranean termite genus Reticulitermes (Isoptera: Rhinotermitidae) in Arkansas and Louisiana. Fla. Entomol. 2004, 87, 473–480. [Google Scholar] [CrossRef]
- Estoup, A.; Jarne, P.; Cornuet, J.M. Homoplasy and mutation model at microsatellite loci and their consequences for population genetics analysis. Mol. Ecol. 2002, 11, 1591–1604. [Google Scholar] [CrossRef]
- Hausdorf, B.; Hennig, C. Species delimitation and geography. Mol. Ecol. Resour. 2020, 20, 950–960. [Google Scholar] [CrossRef]
- Nagaraju, J.; Reddy, K.D.; Nagaraja, G.M.; Sethuraman, B.N. Comparison of multilocus RFLPs and PCR-based marker systems for genetic analysis of the silkworm, Bombyx mori. Heredity 2001, 86, 588–597. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Counties | ||||||
| Statewide | North of Fall Line | South of Fall Line | ||||
| Species | # of counties (P = 159) | % of counties | # of counties (P = 73) | % of counties | # of counties (P = 86) | % of counties |
| Rf | 159 | 100% | 73 | 100% | 86 | 100% |
| Rn | 128 | 80.5% | 43 | 58.9% | 85 | 98.8% |
| Rm | 73 | 45.9% | 69 | 94.5% | 4 | 4.7% |
| Rv | 122 | 76.7% | 50 | 68.5% | 72 | 83.7% |
| Rh | 18 | 14.1% | 18 | 24.7% | 0 | 0% |
| Sites | ||||||
| Statewide | North of Fall Line | South of Fall Line | ||||
| Species | # of sites (P = 1570) | % of sites | # of sites (P = 688) | % of sites | # of sites (P = 882) | % of sites |
| Rf | 1131 | 72.0% | 504 | 73.3% | 627 | 71.1% |
| Rn | 559 | 35.6% | 130 | 18.9% | 429 | 48.6% |
| Rm | 323 | 20.6% | 313 | 45.5% | 10 | 1.1% |
| Rv | 318 | 20.3% | 97 | 14.1% | 221 | 25.1% |
| Rh | 21 | 1.3% | 21 | 3.0% | 0 | 0% |
| Samples | ||||||
| Statewide | North of Fall Line | South of Fall Line | ||||
| Species | # of samples (n = 4371) | % of samples | # of samples (n = 1975) | % of samples | # of samples (n = 2396) | % of samples |
| Rf | 2260 | 51.7% | 1036 | 52.5% | 1224 | 51.1% |
| Rn | 1031 | 23.6% | 219 | 11.1% | 812 | 33.9% |
| Rm | 557 | 12.7% | 546 | 27.7% | 11 | 0.5% |
| Rv | 494 | 11.3% | 145 | 7.3% | 349 | 14.6% |
| Rh | 29 | 0.7% | 29 | 1.5% | 0 | 0% |
| Hill Numbers (r = 0, 1, 2, ∞) | |||
|---|---|---|---|
| Hill Number (N0) Number of Species | Hill Number (N1) Exponential Shannon (eH) | Hill Number (N2) Inverse Simpson (1/D) | |
| Entire State | 5 | 3.60 | 3.08 |
| North of Fall Line | 5 | 3.55 | 3.00 |
| South of Fall Line | 4 | 2.88 | 2.65 |
| Species | Clark and Evans R Score | p-Value |
|---|---|---|
| Rf | R = 1.19 | <0.001 |
| Rn | R = 1.05 | <0.05 |
| Rm | R = 0.71 | <0.001 |
| Rv | R = 0.99 | 0.79 |
| Rh | R = 0.58 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Johnson, A.; Forschler, B.T. Biodiversity and Distribution of Reticulitermes in the Southeastern USA. Insects 2022, 13, 565. https://doi.org/10.3390/insects13070565
Johnson A, Forschler BT. Biodiversity and Distribution of Reticulitermes in the Southeastern USA. Insects. 2022; 13(7):565. https://doi.org/10.3390/insects13070565
Chicago/Turabian StyleJohnson, Allison, and Brian T. Forschler. 2022. "Biodiversity and Distribution of Reticulitermes in the Southeastern USA" Insects 13, no. 7: 565. https://doi.org/10.3390/insects13070565