
Modified World Health Organization (WHO) Tunnel Test for Higher Throughput Evaluation of Insecticide-Treated Nets (ITNs) Considering the Effect of Alternative Hosts, Exposure Time, and Mosquito Density

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Description of Investigational ITNs
2.3. Mosquitoes
2.4. The Standard WHO Tunnel Test Procedure
2.5. Bait Used and Preparation
2.6. Study Design
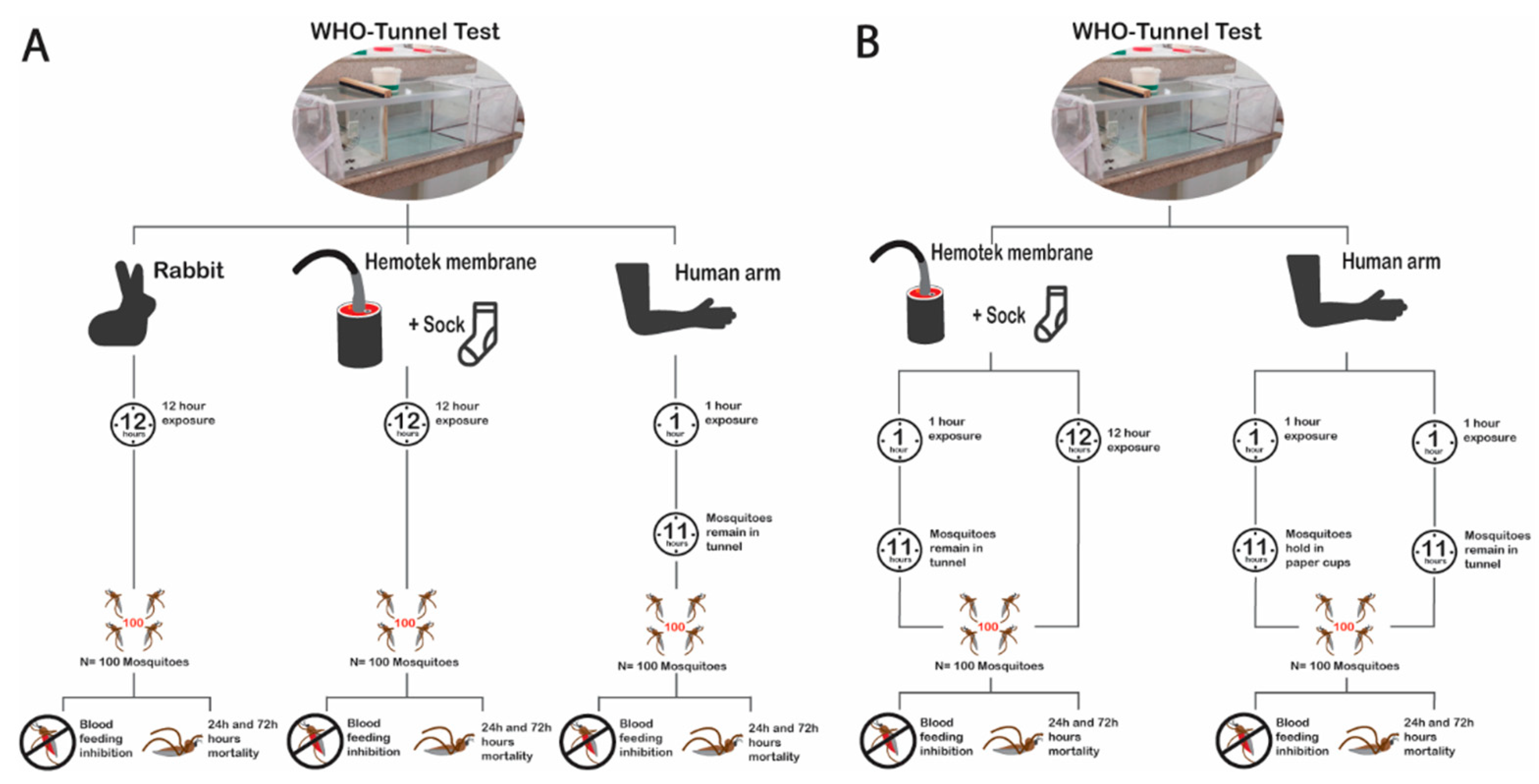
2.6.1. Experiment 1: The Impact of Bait/Host
2.6.2. Experiment 2: The Impact of Exposure Time
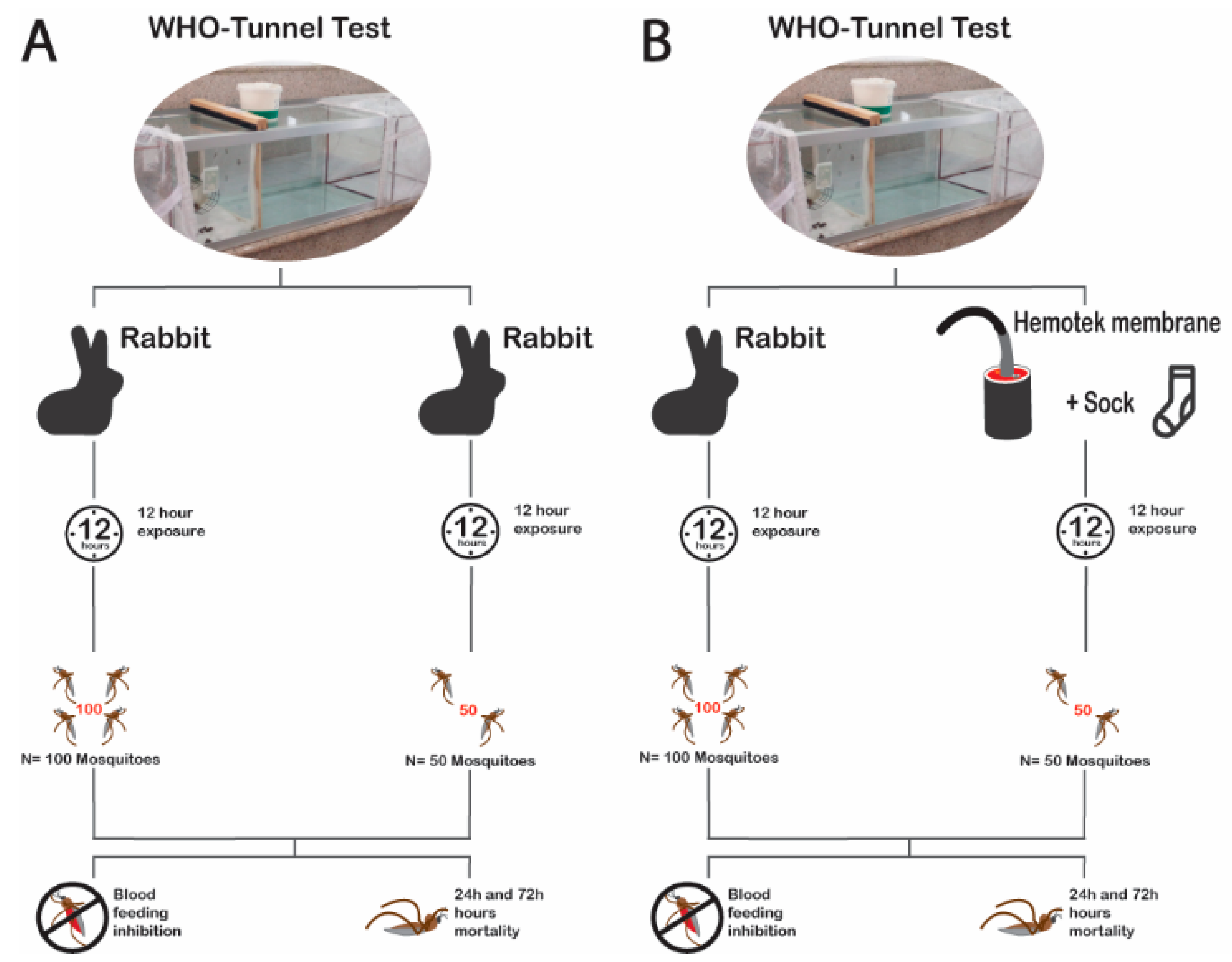
2.6.3. Experiment 3: Effects of Mosquito Density on the Bio-Efficacy Measurement of Blood-Feeding Inhibition and Mortality at 24-h or 72-h
2.6.4. Experiment 4: Possibility to Replace Standard Bait (Rabbit) with the Membrane Assay
2.7. Data Analysis
2.7.1. Sample Size and Power
2.7.2. Statistical Analysis
3. Results
3.1. Experimental Validity
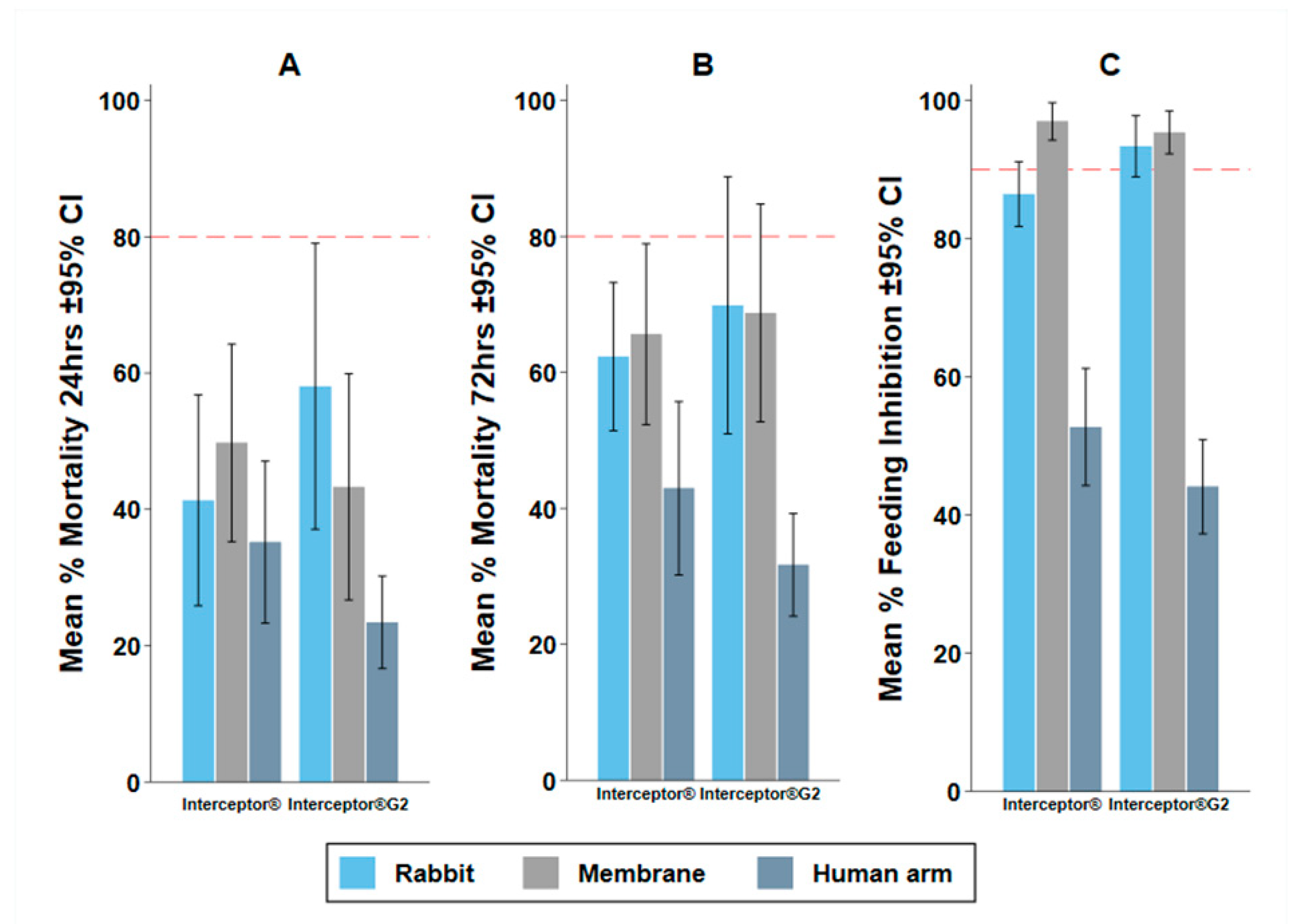
3.2. Experiment 1: The Impact of Baits
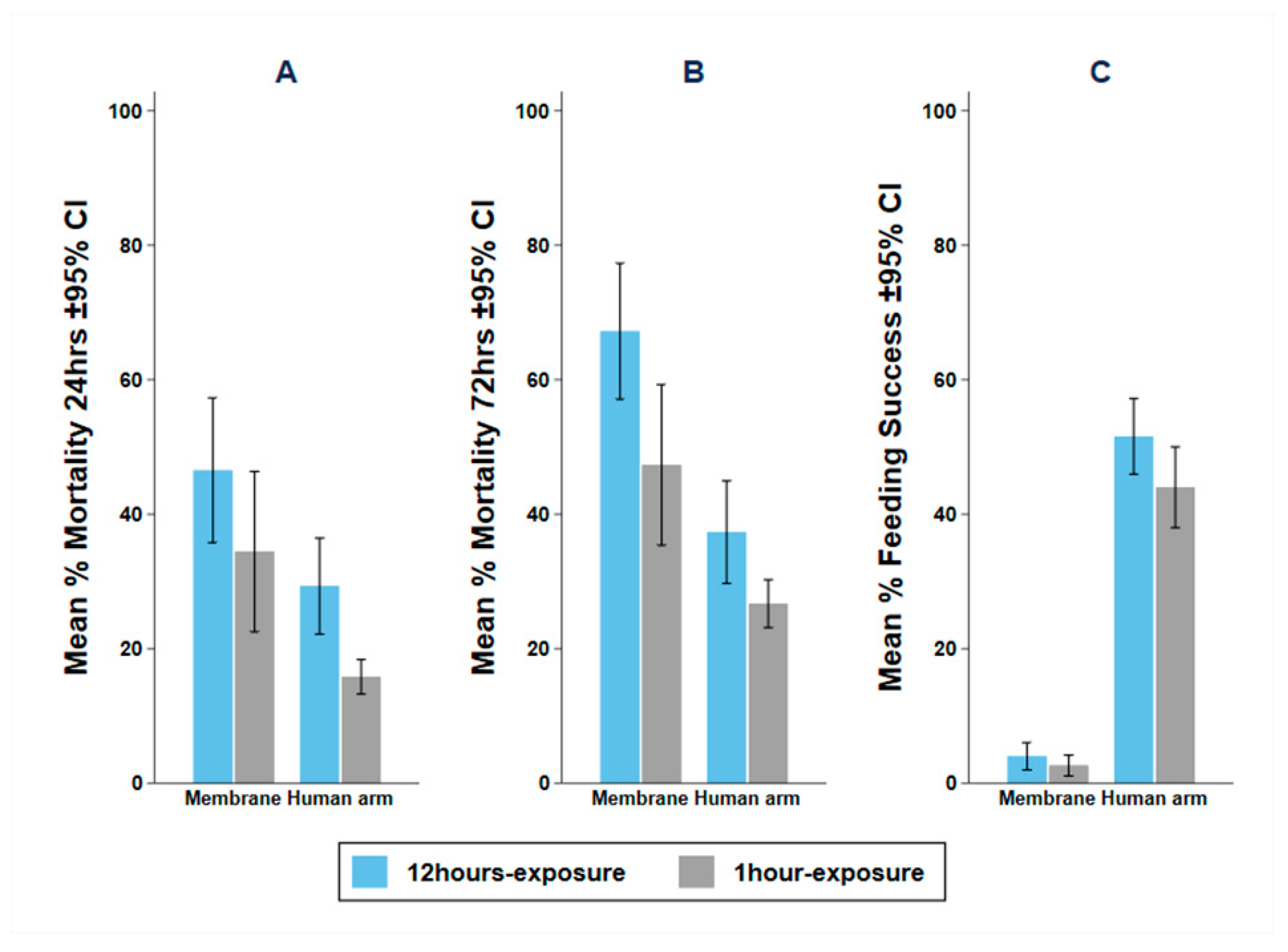
3.3. Experiment 2: Impact of Exposure Time on Mortality and Blood Feeding
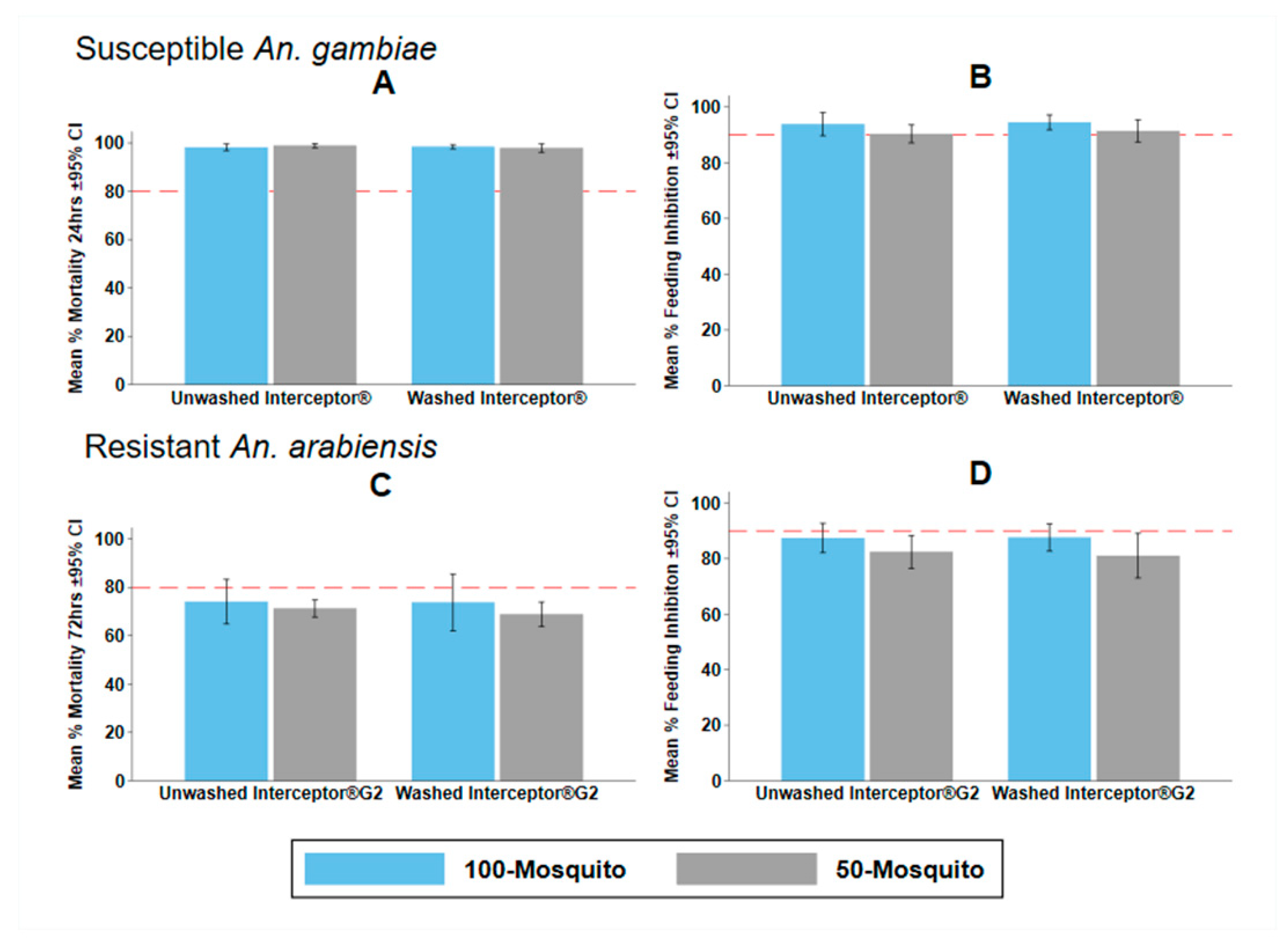
3.4. Experiment 3: Effects of Mosquito Density on Tunnel Test Endpoints
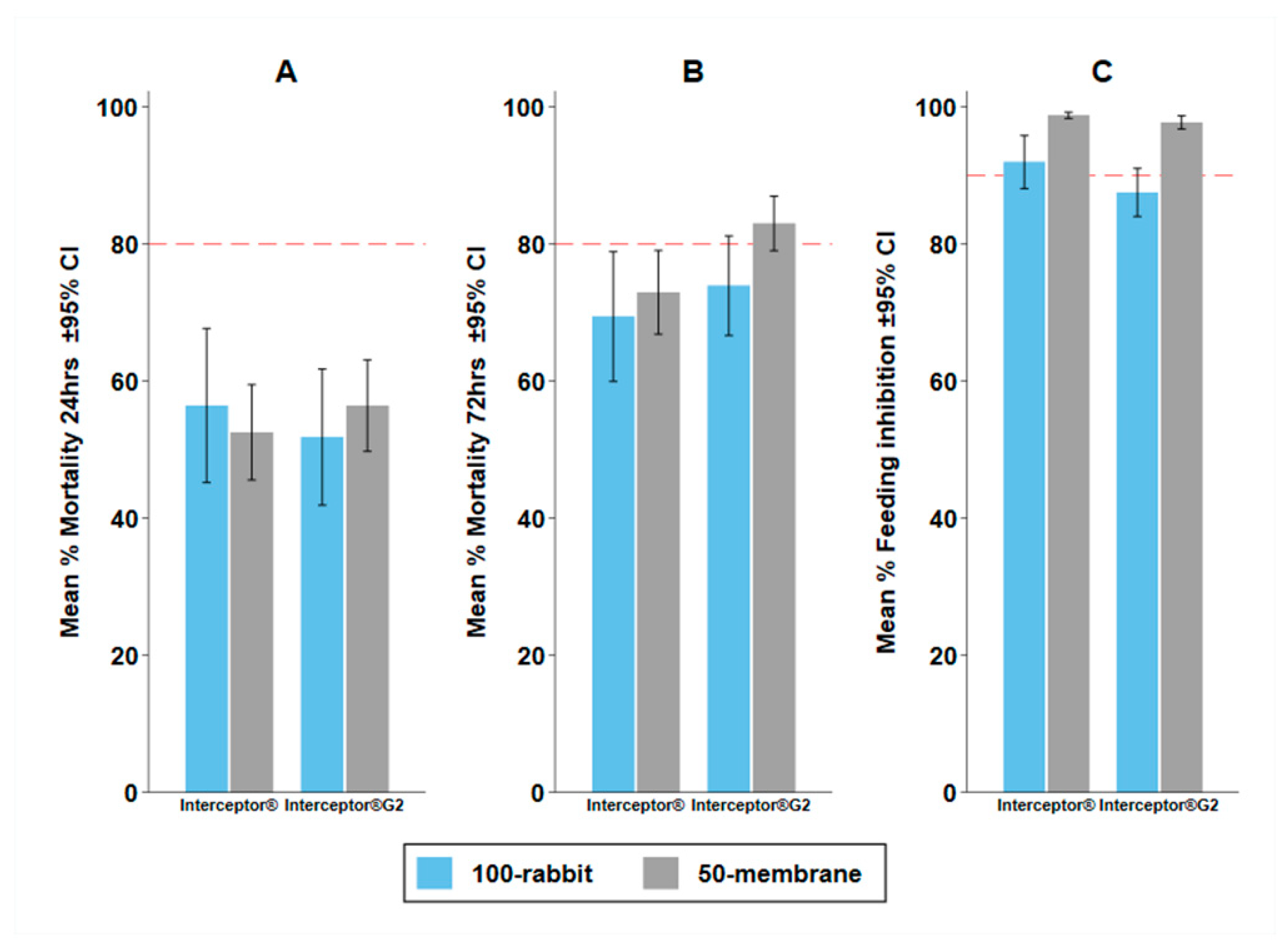
3.5. Experiment 4: Possibility to Replace Standard Bait with the Membrane Feeding
4. Discussion
4.1. Impact of the Bait
4.2. Impact of Exposure Time
4.3. Effects of Mosquito Density
4.4. Study Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021; pp. 1–322. [Google Scholar]
- Pryce, J.; Richardson, M.; Lengeler, C. Insecticide-treated nets for preventing malaria. Cochrane Database Syst. Rev. 2018, 11, CD000363. [Google Scholar] [CrossRef]
- Birget, P.L.G.; Koella, J.C. An Epidemiological Model of the Effects of Insecticide-Treated Bed Nets on Malaria Transmission. PLoS ONE 2015, 10, e0144173. [Google Scholar] [CrossRef]
- Koella, J.C.; Lynch, P.A.; Thomas, M.B.; Read, A.F. Towards evolution-proof malaria control with insecticides. Evol. Appl. 2009, 2, 469–480. [Google Scholar] [CrossRef]
- Chandre, F.; Darriet, F.; Duchon, S.; Finot, L.; Manguin, S.; Carnevale, P.; Guillet, P. Modifications of pyrethroid effects associated with kdr mutation in Anopheles gambiae. Med. Vet. Entomol. 2000, 14, 81–88. [Google Scholar] [CrossRef]
- Carnevale, P.B.; Bitsindou, P.; Diomandé, L.; Robert, V. Insecticide impregnation can restore the efficiency of torn bed nets and reduce man-vector contact in malaria endemic areas. Trans. R. Soc. Trop. Med. 1992, 86, 362–364. [Google Scholar] [CrossRef]
- Nkya, T.E.; Akhouayri, I.; Poupardin, R.; Batengana, B.; Mosha, F.; Magesa, S.; Kisinza, W.; David, J.-P. Insecticide resistance mechanisms associated with different environments in the malaria vector Anopheles Gamb. A Case Study Tanzania. Malar. J. 2014, 13, 28. [Google Scholar] [CrossRef] [Green Version]
- WHO. Global Technical Strategy for Malaria 2016–2030, 2021 Update; World Health Organization: Geneva, Switzerland, 2021; pp. 1–40. [Google Scholar]
- N’Guessan, R.; Odjo, A.; Ngufor, C.; Malone, D.; Rowland, M. A Chlorfenapyr Mixture Net Interceptor® G2 Shows High Efficacy and Wash Durability against Resistant Mosquitoes in West Africa. PLoS ONE 2016, 11, e0165925. [Google Scholar] [CrossRef] [Green Version]
- Oxborough, R.M.; Kitau, J.; Matowo, J.; Feston, E.; Mndeme, R.; Mosha, F.W.; Rowland, M.W. ITN mixtures of chlorfenapyr (Pyrrole) and alphacypermethrin (Pyrethroid) for control of pyrethroid resistant Anopheles arabiensis and Culex quinquefasciatus. PLoS ONE 2013, 8, e55781. [Google Scholar] [CrossRef]
- Ngufor, C.; Agbevo, A.; Fagbohoun, J.; Fongnikin, A.; Rowland, M. Efficacy of Royal Guard, a new alpha-cypermethrin and pyriproxyfen treated mosquito net, against pyrethroid-resistant malaria vectors. Sci. Rep. 2020, 10, 12227. [Google Scholar] [CrossRef]
- Martin, J.L.; Mosha, F.W.; Lukole, E.; Rowland, M.; Todd, J.; Charlwood, J.D.; Mosha, J.F.; Protopopoff, N. Personal protection with PBO-pyrethroid synergist-treated nets after 2 years of household use against pyrethroid-resistant Anopheles in Tanzania. Parasites Vectors 2021, 14, 150. [Google Scholar] [CrossRef]
- Bayili, K.; N’Do, S.; Yadav, R.S.; Namountougou, M.; Ouattara, A.; Dabiré, R.K.; Ouédraogo, G.A.; Diabate, A. Experimental hut evaluation of DawaPlus 3.0 LN and DawaPlus 4.0 LN treated with deltamethrin and PBO against free-flying populations of Anopheles gambiae s.l. in Vallée du Kou, Burkina Faso. PLoS ONE 2019, 14, e0226191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayili, K.; N’Do, S.; Namountougou, M.; Sanou, R.; Ouattara, A.; Dabiré, R.K.; Ouédraogo, A.G.; Malone, D.; Diabaté, A. Evaluation of efficacy of Interceptor(®) G2, a long-lasting insecticide net coated with a mixture of chlorfenapyr and alpha-cypermethrin, against pyrethroid resistant Anopheles gambiae s.l. in Burkina Faso. Malar. J. 2017, 16, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staedke, S.G.; Gonahasa, S.; Dorsey, G.; Kamya, M.R.; Maiteki-Sebuguzi, C.; Lynd, A.; Katureebe, A.; Kyohere, M.; Mutungi, P.; Kigozi, S.P.; et al. Effect of long-lasting insecticidal nets with and without piperonyl butoxide on malaria indicators in Uganda (LLINEUP): A pragmatic, cluster-randomised trial embedded in a national LLIN distribution campaign. Lancet 2020, 395, 1292–1303. [Google Scholar] [CrossRef]
- Gleave, K.; Lissenden, N.; Chaplin, M.; Choi, L.; Ranson, H. Piperonyl butoxide (PBO) combined with pyrethroids in insecticide-treated nets to prevent malaria in Africa. J. Cochrane Database Syst. Rev. 2021. [Google Scholar] [CrossRef] [PubMed]
- Mosha, J.F.; Kulkarni, M.A.; Lukole, E.; Matowo, N.S.; Pitt, C.; Messenger, L.A.; Mallya, E.; Jumanne, M.; Aziz, T.; Kaaya, R.; et al. Effectiveness and cost-effectiveness against malaria of three types of dual-active-ingredient long-lasting insecticidal nets (LLINs) compared with pyrethroid-only LLINs in Tanzania: A four-arm, cluster-randomised trial. Lancet 2022, 399, 1227–1241. [Google Scholar] [CrossRef]
- PATH. New Nets Project Interim Report 2022; PATH: Seattle, WA, USA, 2022; Volume 1–96. [Google Scholar]
- Hougard, J.M.; Duchon, S.; Darriet, F.; Zaim, M.; Rogier, C.; Guillet, P. Comparative performances, under laboratory conditions, of seven pyrethroid insecticides used for impregnation of mosquito nets. Malar. J. 2003, 5, 324–333. [Google Scholar]
- WHOPES. Guidelines for Laboratory and Field Testing of Long Lasting Insecticidal Nets; WHOPES: Geneva, Switzerland, 2013; Volume 2, p. 102. [Google Scholar]
- David, M.D. The potential of pro-insecticides for resistance management. J. Pest Manag. Sci. 2021, 77, 3631–3636. [Google Scholar] [CrossRef]
- Oxborough, R.M.; N’Guessan, R.; Jones, R.; Kitau, J.; Ngufor, C.; Malone, D.; Mosha, F.W.; Rowland, M.W. The activity of the pyrrole insecticide chlorfenapyr in mosquito bioassay: Towards a more rational testing and screening of non-neurotoxic insecticides for malaria vector control. Malar. J. 2015, 14, 124. [Google Scholar] [CrossRef] [Green Version]
- Kibondo, U.A.; Odufuwa, O.G.; Ngonyani, S.H.; Mpelepele, A.B.; Issaya, M.; Ngonyani, H.; Matanila, I.; Noel, O.M.; Antony, P.M.; Kyeba, S.; et al. Influence of testing modality on bioefficacy for the evaluation of Interceptor® G2 mosquito nets to combat malaria mosquitoes in Tanzania. Parasite Vectors 2022, 15, 124. [Google Scholar] [CrossRef]
- Takken, W.; Knols, B.G. Odor-mediated behavior of Afrotropical malaria mosquitoes. Annu. Rev. Entomol. 1999, 44, 131–157. [Google Scholar] [CrossRef]
- Takken, W.; Verhulst, N.O. Host preferences of blood-feeding mosquitoes. Annu. Rev. Entomol. 2013, 58, 433–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, J.E.A.; Angarita Jaimes, N.C.; Gleave, K.; Mashauri, F.; Abe, M.; Martine, J.; Towers, C.E.; Towers, D.; McCall, P.J. Host-seeking activity of a Tanzanian population of Anopheles arabiensis at an insecticide treated bed net. Malar. J. 2017, 16, 270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debboun, M.; Wagman, J. In vitro repellency of N,N-diethyl-3-methylbenzamide and N,N-diethylphenylacetamide analogs against Aedes aegypti and Anopheles stephensi (Diptera: Culicidae). J. Med. Entomol. 2004, 41, 430–434. [Google Scholar] [CrossRef]
- Vallejo, A.F.; Rubiano, K.; Amado, A.; Krystosik, A.R.; Herrera, S.; Arévalo-Herrera, M. Optimization of a Membrane Feeding Assay for Plasmodium vivax Infection in Anopheles albimanus. PLoS Negl. Trop. Dis. 2016, 10, e0004807. [Google Scholar] [CrossRef] [Green Version]
- Smit, M.R.; Ochomo, E.O.; Aljayyoussi, G.; Kwambai, T.K.; Abong’o, B.O.; Bousema, T.; Waterhouse, D.; Bayoh, N.M.; Gimnig, J.E.; Samuels, A.M.; et al. Human Direct Skin Feeding versus Membrane Feeding to Assess the Mosquitocidal Efficacy of High-Dose Ivermectin (IVERMAL Trial). Clin. Infect. Dis. 2018, ciy1063, 1112–1119. [Google Scholar] [CrossRef]
- Awono-Ambene, H.P.; Diawara, L.; Robert, V. Comparison of direct and membrane feeding methods to infect Anopheles arabiensis with Plasmodium falciparum. Am. J. Trop. Med. Hyg. 2001, 64, 32–34. [Google Scholar] [CrossRef] [Green Version]
- Damiens, D.; Soliban, S.M.; Balestrino, F.; Alsir, R.; Vreysen, M.; Gilles, J. Different Blood and Sugar Feeding Regimes Affect the Productivity of Anopheles arabiensis Colonies (Diptera: Culicidae). J. Med. Entomol. 2013, 50, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Gunathilaka, N.; Ranathunge, T.; Udayanga, L.; Abeyewickreme, W. Efficacy of Blood Sources and Artificial Blood Feeding Methods in Rearing of Aedes aegypti (Diptera: Culicidae) for Sterile Insect Technique and Incompatible Insect Technique Approaches in Sri Lanka. BioMed Res. Int. 2017, 2017, 3196924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phasomkusolsil, S.; Tawong, J.; Monkanna, N.; Pantuwatana, K.; Damdangdee, N.; Khongtak, W.; Kertmanee, Y.; Evans, B.P.; Schuster, A.L. Maintenance of mosquito vectors: Effects of blood source on feeding, survival, fecundity, and egg hatching rates. J. Vector Ecol. 2013, 38, 38–45. [Google Scholar] [CrossRef]
- Robert, V. Age grading Anopheles arabiensis: Their gorging and surviving responses using a membrane feeding system. Parasite 1998, 5, 87–90. [Google Scholar] [CrossRef] [Green Version]
- Timinao, L.; Vinit, R.; Katusele, M.; Schofield, L.; Burkot, T.R.; Karl, S. Optimization of the feeding rate of Anopheles farauti s.s. colony mosquitoes in direct membrane feeding assays. Parasite Vectors 2021, 14, 356. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, C. Methods in Anopheles Research Manual; BEI Resources: Washington, DC, USA, 2014; p. 419. [Google Scholar]
- Ouédraogo, A.L.; Guelbéogo, W.M.; Cohuet, A.; Morlais, I.; King, J.G.; Gonçalves, B.P.; Bastiaens, G.J.H.; Vaanhold, M.; Sattabongkot, J.; Wu, Y.; et al. A protocol for membrane feeding assays to determine the infectiousness of P. falciparum naturally infected individuals to Anopheles gambiae. Malar. World J. 2013, 1, 4. [Google Scholar]
- Pates, H.V.; Takken, W.; Stuke, K.; Curtis, C.F. Differential behaviour of Anopheles gambiae sensu stricto (Diptera: Culicidae) to human and cow odours in the laboratory. Bull. Entomol. Res. 2001, 91, 289–296. [Google Scholar] [CrossRef] [Green Version]
- Edman, J.D.; Webber, L.A.; Kale, H.W., 2nd. Effect of mosquito density on the interrelationship of host behavior and mosquito feeding success. Am. J. Trop. Med. Hyg. 1972, 21, 487–491. [Google Scholar] [CrossRef]
- Lines, J.D.; Myamba, J.; Curtis, C.F. Experimental hut trials of permethrin-impregnated mosquito nets and eave curtains against malaria vectors in Tanzania. Med. Vet. Entomol. 1987, 1, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Lissenden, N.; Armistead, J.S.; Gleave, K.; Irish, S.R.; Martin, J.L.; Messenger, L.A.; Moore, S.J.; Ngufor, C.; Protopopoff, N.; Oxborough, R.; et al. Developing Consensus Standard Operating Procedures (SOPs) to Evaluate New Types of Insecticide-Treated Nets. Insects 2022, 13, 7. [Google Scholar] [CrossRef]
- Johnson, P.C.D.; Barry, S.J.E.; Ferguson, H.F.; Muller, P. Power analysis for generalized linear mixed models in ecology and evolution. Methods Ecol. Evol. 2014, 6, 133–142. [Google Scholar] [CrossRef]
- Bland, J.M.; Altman, D.G. Measuring agreement in method comparison studies. Stat. Methods Med. Res. 1999, 8, 135–160. [Google Scholar] [CrossRef]
- Abilio, A.P.; Obi, E.; Koenker, H.; Babalola, S.; Saifodine, A.; Zulliger, R.; Swamidoss, I.; Ponce de Leon, G.; Alfai, E.; Blaufuss, S.; et al. Monitoring the durability of the long-lasting insecticidal nets MAGNet and Royal Sentry in three ecological zones of Mozambique. Malar. J. 2020, 19, 209. [Google Scholar] [CrossRef]
- Kilian, A.; Koenker, H.; Obi, E.; Selby, R.A.; Fotheringham, M.; Lynch, M. Field durability of the same type of long-lasting insecticidal net varies between regions in Nigeria due to differences in household behaviour and living conditions. Malar. J. 2015, 14, 123. [Google Scholar] [CrossRef] [Green Version]
- Mutuku, F.M.; Khambira, M.; Bisanzio, D.; Mungai, P.; Mwanzo, I.; Muchiri, E.M.; King, C.H.; Kitron, U. Physical condition and maintenance of mosquito bed nets in Kwale County, coastal Kenya. Malar. J. 2013, 12, 46. [Google Scholar] [CrossRef] [Green Version]
- Kilian, A.; Obi, E.; Mansiangi, P.; Abílio, A.P.; Haji, K.A.; Blaufuss, S.; Olapeju, B.; Babalola, S.; Koenker, H. Variation of physical durability between LLIN products and net use environments: Summary of findings from four African countries. Malar. J. 2021, 20, 26. [Google Scholar] [CrossRef] [PubMed]
- Allan, R.; O’Reilly, L.; Gilbos, V.; Kilian, A. An Observational Study of Material Durability of Three World Health Organization-Recommended Long-Lasting Insecticidal Nets in Eastern Chad. Am. J. Trop. Med. Hyg. 2012, 87, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansiangi, P.; Umesumbu, S.; Etewa, I.; Zandibeni, J.; Bafwa, N.; Blaufuss, S.; Olapeju, B.; Ntoya, F.; Sadou, A.; Irish, S.; et al. Comparing the durability of the long-lasting insecticidal nets DawaPlus(®) 2.0 and DuraNet© in northwest Democratic Republic of Congo. Malar. J. 2020, 19, 189. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, L.M.; Bradley, J.; Yukich, J.; Massue, D.J.; Mageni Mboma, Z.; Pigeon, O.; Moore, J.; Kilian, A.; Lines, J.; Kisinza, W.; et al. Comparative functional survival and equivalent annual cost of 3 long-lasting insecticidal net (LLIN) products in Tanzania: A randomised trial with 3-year follow up. PLOS Med. 2020, 17, e1003248. [Google Scholar] [CrossRef]
- Hossain, M.I.; Curtis, C.F. Permethrin-impregnated bednets: Behavioural and killing effects on mosquitoes. Med. Vet. Entomol. 1989, 3, 367–376. [Google Scholar] [CrossRef]
- N’Guessan, R.; Boko, P.; Odjo, A.; Akogbéto, M.; Yates, A.; Rowland, M. Chlorfenapyr: A pyrrole insecticide for the control of pyrethroid or DDT resistant Anopheles gambiae (Diptera: Culicidae) mosquitoes. Acta Trop. 2007, 102, 69–78. [Google Scholar] [CrossRef]
- Smallegange, R.C.; Verhulst, N.O.; Takken, W. Sweaty skin: An invitation to bite? Trends Parasitol. 2011, 27, 143–148. [Google Scholar] [CrossRef]
- Mukabana, W.R.; Takken, W.; Coe, R.; Knols, B.G. Host-specific cues cause differential attractiveness of Kenyan men to the African malaria vector Anopheles gambiae. Malar. J. 2002, 1, 17. [Google Scholar] [CrossRef]
- van Loon, J.J.A.; Smallegange, R.C.; Bukovinszkiné-Kiss, G.; Jacobs, F.; De Rijk, M.; Mukabana, W.R.; Verhulst, N.O.; Menger, D.J.; Takken, W. Mosquito Attraction: Crucial Role of Carbon Dioxide in Formulation of a Five-Component Blend of Human-Derived Volatiles. J. Chem. Ecol. 2015, 41, 567–573. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, F. Water vapour and carbon dioxide receptors in Aedes aegypti. J. Insect Physiol. 1970, 16, 99–108. [Google Scholar] [CrossRef]
- Hawkes, F.M.; Dabiré, R.K.; Sawadogo, S.P.; Torr, S.; Gibson, G. Exploiting Anopheles responses to thermal, odour and visual stimuli to improve surveillance and control of malaria. Sci. Rep. 2017, 7, 17283. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Maibach, H.I. A study of the probing response of Aedes aegypti. 4. Effect of dry and moist heat on probing. J. Econ. Entomol. 1971, 64, 442–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okumu, F.; Biswaro, L.; Mbeleyela, E.; Killeen, G.F.; Mukabana, R.; Moore, S.J. Using nylon strips to dispense mosquito attractants for sampling the malaria vector Anopheles gambiae s.s. J. Med. Entomol. 2010, 47, 274–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, D.; Stefanini, C.; Canale, A.; Benelli, G. Artificial blood feeders for mosquito and ticks-Where from, where to? Acta Trop. 2018, 183, 43–56. [Google Scholar] [CrossRef]
- Kweka, E.J.; Mwang’onde, B.J.; Lyaruu, L.; Tenu, F.; Mahande, A.M. Effect of Different Hosts on Feeding Patterns and Mortality of Mosquitoes (Diptera: Culicidae) and their Implications on Parasite. Am. J. Trop. Med. Hyg. 2010, 2, 121. [Google Scholar] [CrossRef]
- Mburu, M.M.; Mweresa, C.K.; Omusula, P.; Hiscox, A.; Takken, W.; Mukabana, W.R. 2-Butanone as a carbon dioxide mimic in attractant blends for the Afrotropical malaria mosquitoes Anopheles gambiae and Anopheles funestus. Malar. J. 2017, 16, 351. [Google Scholar] [CrossRef]
- Morimoto, Y.; Kawada, H.; Kuramoto, K.Y.; Mitsuhashi, T.; Saitoh, T.; Minakawa, N. New mosquito repellency bioassay for evaluation of repellents and pyrethroids using an attractive blood-feeding device. Parasites Vectors 2021, 14, 151. [Google Scholar] [CrossRef]
- Anderson, R.A.; Brust, R.A. Blood Feeding Success of Aedes Aegypti and Culex Nigripalpus (Diptera: Cullicidae) in Relation to Defensive Behavior of Japanese Quail (Coturnix Japonica) in the Laboratory. J. Vector Ecol. 1996, 21, 94–104. [Google Scholar]
- Walker, E.D.; Edman, J.D. Influence of defensive behavior of eastern chipmunks and gray squirrels (Rodentia: Sciuridae) on feeding success of Aedes triseriatus (Diptera: Culicidae). J. Med. Entomol. 1986, 23, 1–10. [Google Scholar] [CrossRef]
- Styer, L.M.; Carey, J.R.; Wang, J.-L.; Scott, T.W. Mosquitoes do senesce: Departure from the paradigm of constant mortality. Am. J. Trop. Med. Hyg. 2007, 76, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Hauser, G.; Thiévent, K.; Koella, J.C. The ability of Anopheles gambiae mosquitoes to bite through a permethrin-treated net and the consequences for their fitness. Sci. Rep. 2019, 9, 8141. [Google Scholar] [CrossRef] [PubMed]
- Machani, M.G.; Ochomo, E.; Sang, D.; Bonizzoni, M.; Zhou, G.; Githeko, A.K.; Yan, G.; Afrane, Y.A. Influence of blood meal and age of mosquitoes on susceptibility to pyrethroids in Anopheles gambiae from Western Kenya. Malar. J. 2019, 18, 112. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | 1 | 2 | 3 | 4 | |
|---|---|---|---|---|---|
| Factor | Host/Baits | Exposure Time | Mosquito Density | Replacement of Rabbit | |
| Comparison | Human or membrane vs. rabbit with 100 mosquitoes | 1 h vs. 12 h for human or membrane (within host) | 50 vs 100 mosquitoes using rabbit | Rabbit with 100 mosquitoes vs. membrane with 50 mosquitoes | |
| ITNs arms | Interceptor® G2 Unwashed Interceptor® G2 Washed 20× Interceptor® Unwashed Interceptor® Washed 20× Negative control | Interceptor® G2 Unwashed Interceptor® G2 Washed 20× Negative control | Interceptor® Unwashed Interceptor® Washed 20× Negative control | Interceptor® G2 Unwashed Interceptor® G2 Washed 20× Interceptor® Unwashed Interceptor® Washed 20× Negative control | |
| Replicates per arm per comparison | 5 | 15 | 15 | 15 | |
| Total replicates | 75 | 100 | 90 | 90 | 150 |
| Number of nights | 15 | 10 | 10 | 10 | 16 |
| Mosquitoes exposed | 100 | 100, 50 | 100, 50 | ||
| Host/bait | Rabbit, Human, Membrane | Human, Membrane | Rabbit | Rabbit-100, Membrane-50 | |
| Exposure time | 12 h | 12 h 1 h | 12 h | 12 h | |
| Mosquito species | Anopheles arabiensis | Anopheles arabiensis | Anopheles gambiae | Anopheles arabiensis | |
| Primary Outcomes | Blood feeding success (BFS), 24-h mortality (M24), 72-h mortality (M72) | ||||
| Additional Outcome | Blood feeding Inhibition (BFI) | ||||
| BFS | M24 | M72 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| % (95% CI) | OR (95%CI) | p-Value | % (95% CI) | OR (95%CI) | p-Value | % (95% CI) | OR (95%CI) | p-Value | |
| Control | |||||||||
| Rabbit | 64.8 (51.2–78.3) | 1 | 3.8 (0.8-6.8) | 1 | 7.7 (5.1–10.3) | 1 | |||
| Membrane | 22.8 (10.4–35.1) | 0.16 (0.14–0.20) | <0.001 | 6.8 (5.9–7.6) | 1.83 (1.22–2.75) | 0.004 | 8.9 (8.3–9.5) | 1.16 (0.84–1.59) | 0.366 |
| Human arm | 74.4 (67.9–80.8) | 1.59 (1.25–2.02) | <0.001 | 6.4 (4.9–7.8) | 1.71 (1.05–2.77) | 0.030 | 11.7 (9.0–14.4) | 1.58 (1.11–2.26) | 0.012 |
| Treatment | |||||||||
| Rabbit | 6.6 (2.2–11.0) | 1 | 49.7 (36.4–62.9) | 1 | 66.1 (55.3–76.9) | 1 | |||
| Membrane | 4.6 (1.5–7.7) | 0.34 (0.28–0.48) | <0.001 | 46.5 (35.7–57.3) | 0.90 (0.79–1.02) | 0.086 | 67.2 (57.0–77.3) | 1.07 (0.93–1.22) | 0.352 |
| Human arm | 55.9 (49.1–62.7) | 9.81 (8.25–11.67) | < 0.001 | 29.3 (22.1–36.5) | 0.42 (0.37–0.48) | <0.001 | 37.3 (29.7–45.0) | 0.31 (0.27–0.35) | <0.001 |
| Assays | BFS | M24 | M72 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| % | OR | p-Value | % | OR | p-Value | % | OR | p-Value | |
| (95% CI) | (95% CI) | (95% CI) | (95% CI) | (95% CI) | (95% CI) | ||||
| Membrane | |||||||||
| 1 h-exposure | 1.2 (0.1–2.3) | 1 | 24.7 (17.0–32.4) | 1 | 38.9 (26.5–51.2) | 1 | |||
| 12 h-exposure | 4.6 (1.5–7.7) | 1.55 (1.08–2.22) | 0.017 | 43.3 (25.9–60.6) | 1.66 (1.46–1.89) | <0.001 | 68.8 (52.0–85.5) | 2.30 (2.02–2.62) | <0.001 |
| Human arm | |||||||||
| 1 h-exposure | NA | 20.3 (17.7–22.8) | 1 | 31.1 (26.1–36.1) | 1 | ||||
| 12 h-exposure | NA | 35.2 (22.7–47.6) | 2.26 (1.93–2.64) | <0.001 | 43.0 (29.6–56.3) | 1.66 (1.45–1.90) | <0.001 | ||
| BFS | M24 | M72 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Density | % | OR | p-Value | % | OR | p-Value | % | OR | p-Value |
| (95% CI) | (95% CI) | (95% CI) | (95% CI) | (95% CI) | (95% CI) | ||||
| Susceptible An. gambiae with Interceptor® | |||||||||
| 100 Mosquitoes | 5.8 (3.4–8.2) | 1 | 98.3 (97.5–99.1) | 1 | 99.1 (98.6–99.6) | 1 | |||
| 50 Mosquitoes | 9.1 (6.6–11.6) | 2.35 (0.80–6.92) | 0.122 | 98.4 (97.5–99.3) | 1.10 (0.32–3.72) | 0.882 | 99.6 (99.3–99.9) | 1.80 (0.43–7.54) | 0.421 |
| Resistant An. arabiensis with Interceptor® G2 | |||||||||
| 100 Mosquitoes | 12.5 (8.9–16.0) | 1 | 51.8 (41.9–61.7) | 1 | 73.9 (66.7–81.2) | 1 | |||
| 50 Mosquitoes | 18.3 (13.3–23.2) | 1.54 (0.74–3.22) | 0.249 | 45.1 (40.7–49.6) | 0.69 (0.23–2.12) | 0.518 | 70.0 (67.0–73.1) | 0.65 (0.25–1.67) | 0.375 |
| Treatment | 100-Rabbit | 50-Rabbit | ||||||
|---|---|---|---|---|---|---|---|---|
| BFS | M72 | BFS | M72 | |||||
| OR (95% CI) | p-Value | OR (95% CI) | p-Value | OR (95% CI) | p-Value | OR (95% CI) | p-Value | |
| Overall | ||||||||
| Interceptor® | 1 | 1 | 1 | 1 | 1 | |||
| Interceptor® G2 | 1.76 (1.55–1.99) | <0.001 | 1.23 (1.13–1.33) | <0.001 | 12.93 (9.63–17.36) | <0.001 | 1.41 (1.26–1.57) | <0.001 |
| Unwashed | ||||||||
| Interceptor® | 1 | 1 | 1 | 1 | 1 | |||
| Interceptor® G2 | 1.64 (1.38–1.95) | <0.001 | 1.15 (1.02–1.29) | 0.018 | 8.50 (5.95–12.15) | <0.001 | 1.83 (1.56–2.14) | <0.001 |
| Washed 20× | ||||||||
| Interceptor® | 1 | 1 | 1 | 1 | 1 | |||
| Interceptor® G2 | 1.90 (1.58–2.27) | <0.001 | 1.31 (1.17–1.47) | <0.001 | 24.34 (14.16–41.85) | <0.001 | 1.07 (0.85–1.34) | 0.432 |
| Assay | BFS | p-Value | M24 | p-Value | M72 | p-Value | |||
|---|---|---|---|---|---|---|---|---|---|
| % (95% CI) | OR (95% CI) | % (95% CI) | OR (95% CI) | % (95% CI) | OR (95% CI) | ||||
| Interceptor® | |||||||||
| 100 Rabbit | 7.9 (4.1–11.8) | 1 | 56.4 (45.3–67.6) | 1 | 69.4 (60.0–78.8) | 1 | |||
| 50 Membrane | 1.2 (0.8–1.7) | 0.19 (0.08–0.45) | <0.001 | 52.5 (45.6–59.4) | 0.39 (0.10–1.61) | 0.195 | 73.0 (66.9–79.0) | 0.54 (0.14–2.06) | 0.370 |
| Interceptor® G2 | |||||||||
| 100 Rabbit | 12.5 (9.0–16.0) | 1 | 51.8 (42.0–61.7) | 1 | 73.9 (66.7–81.1) | 1 | |||
| 50 Membrane | 2.3 (1.3–3.2) | 0.17 (0.09–0.30) | <0.001 | 56.4 (49.8–63.1) | 1.10 (0.51–2.36) | 0.814 | 83.0 (79.1–86.9) | 1.50 (0.75–2.98) | 0.251 |
| Treatment | 100-Rabbit | 50-Membrane | ||||||
|---|---|---|---|---|---|---|---|---|
| BFS | M72 | BFS | M72 | |||||
| OR (95% CI) | p-Value | OR (95% CI) | p-Value | OR (95% CI) | p-Value | OR (95% CI) | p-Value | |
| Overall | ||||||||
| Interceptor® | 1 | 1 | 1 | 1 | ||||
| Interceptor® G2 | 1.76 (1.47–2.10) | <0.001 | 1.23 (1.10–1.38) | <0.001 | 1.87 (1.05–3.33) | 0.033 | 1.79 (1.50–2.14) | <0.001 |
| Unwashed | ||||||||
| Interceptor® | 1 | 1 | 1 | 1 | ||||
| Interceptor® G2 | 1.64 (1.28–2.09) | <0.001 | 1.15 (0.98–1.35) | 0.094 | 2.34 (1.11–4.93) | 0.025 | 1.81 (1.43–2.29) | <0.001 |
| Washed 20× | ||||||||
| Interceptor® | 1 | 1 | 1 | 1 | ||||
| Interceptor® G2 | 1.90 (1.47–2.45) | <0.001 | 1.31 (1.12–1.54) | 0.001 | 1.26 (0.49–3.20) | 0.634 | 1.82 (1.39–2.37) | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamande, D.S.; Odufuwa, O.G.; Mbuba, E.; Hofer, L.; Moore, S.J. Modified World Health Organization (WHO) Tunnel Test for Higher Throughput Evaluation of Insecticide-Treated Nets (ITNs) Considering the Effect of Alternative Hosts, Exposure Time, and Mosquito Density. Insects 2022, 13, 562. https://doi.org/10.3390/insects13070562
Kamande DS, Odufuwa OG, Mbuba E, Hofer L, Moore SJ. Modified World Health Organization (WHO) Tunnel Test for Higher Throughput Evaluation of Insecticide-Treated Nets (ITNs) Considering the Effect of Alternative Hosts, Exposure Time, and Mosquito Density. Insects. 2022; 13(7):562. https://doi.org/10.3390/insects13070562
Chicago/Turabian StyleKamande, Dismas S., Olukayode G. Odufuwa, Emmanuel Mbuba, Lorenz Hofer, and Sarah J. Moore. 2022. "Modified World Health Organization (WHO) Tunnel Test for Higher Throughput Evaluation of Insecticide-Treated Nets (ITNs) Considering the Effect of Alternative Hosts, Exposure Time, and Mosquito Density" Insects 13, no. 7: 562. https://doi.org/10.3390/insects13070562