
Identification and Characterization of Resistance of Three Aphid Species on Contrasting Alfalfa Cultivars

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants and Insects
2.2. Preliminary Experiment: Screening of Aphid-Resistance of Alfalfa Cultivars
2.3. Aphid-Resistance of Two Contrasting Alfalfa Cultivars
- Experiment 1: Colonization ability of aphids on caged plants
- Experiment 2: Evaluation of antixenosis
- Experiment 3: Electrical penetration graphs
2.4. Statistical Analysis
3. Results
3.1. Preliminary Experiment: Weight of the Three Species of Aphids Cultured on 16 Cultivars of Alfalfa
3.2. Fecundity of Three Aphid Species Aphids on Caged Plants of Two Alfalfa Cultivars: Gannong NO.9 and Pegasis
3.3. The Preference of the Three Aphis Species for Settling on Plants of Two Alfalfa Cultivars: Gannong NO.9 and Pegasis
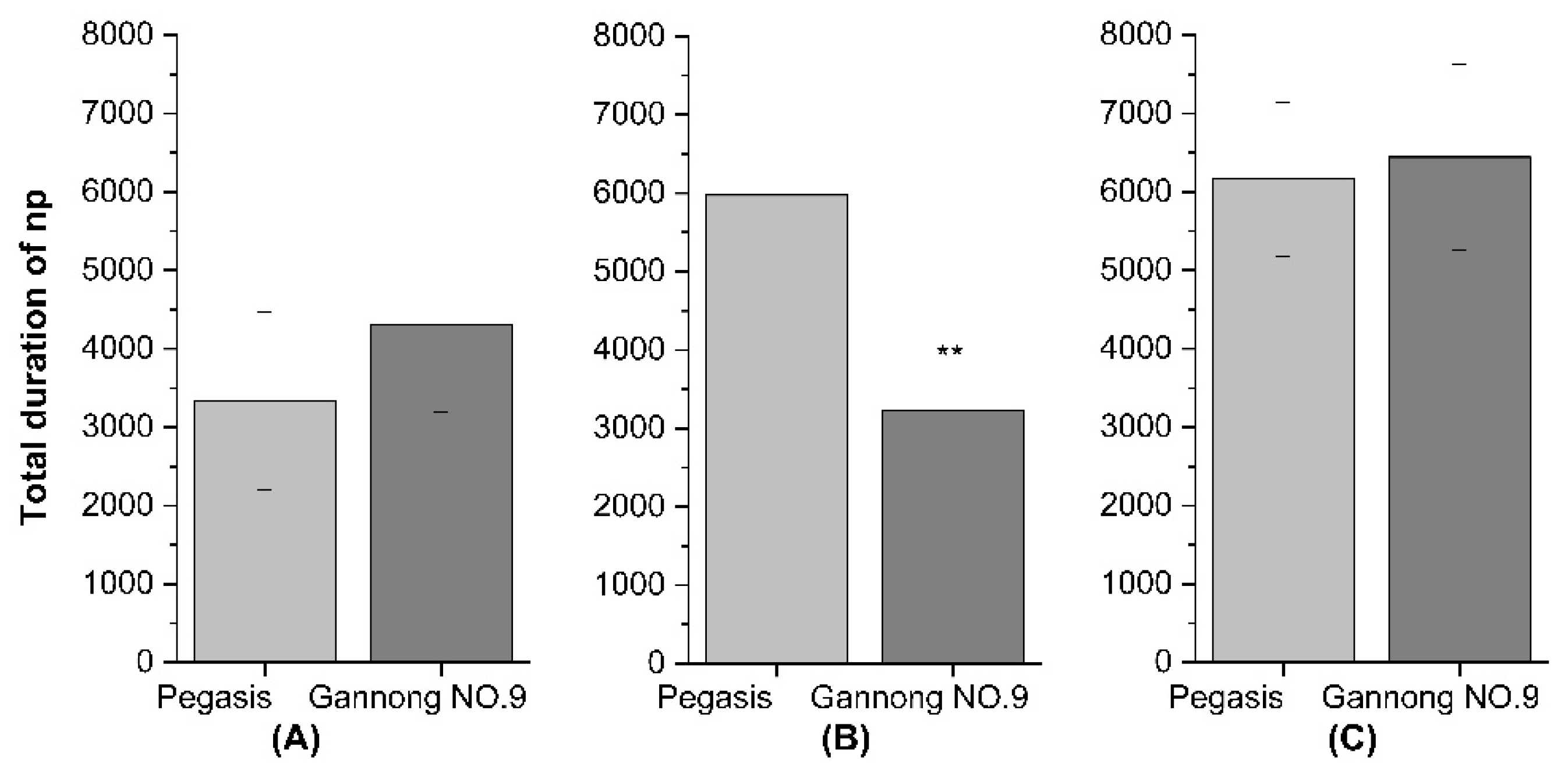
3.4. The Feeding Behavior of the Three Aphid Species on Plants of Two Alfalfa Cultivars: Gannong NO.9 and Pegasis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carvalho, M.G.; Bortolotto, O.C.; Ventura, M.U. Aromatic plants affect the selection of host tomato plants by Bemisia tabaci biotype B. Entomol. Exp. Appl. 2017, 162, 86–92. [Google Scholar] [CrossRef] [Green Version]
- Karban, R.; Myers, J.H. Induced Plant Responses to Herbivory. J. Annu. Rev. Ecol. Syst. 1989, 20, 331–348. [Google Scholar] [CrossRef]
- Garima, S.; Ashokrao, M.P.; Vartika, M. Insect–Plant Interactions: A Multilayered Relationship. Ann. Entomol. Soc. Am. 2020, 114, 1–16. [Google Scholar]
- Painter, R.H. Resistance of Plants to Insects. Annu. Rev. Entomol. 1958, 3, 267–290. [Google Scholar] [CrossRef]
- Panda, N.; Khush, G.A.J.W. Host Plant Resistance to Insects; CABI: Oxon, UK, 1995. [Google Scholar]
- Bouhssini, M.E.; Amri, A.; Lhaloui, S. Plant Resistance to Cereal and Food Legume Insect pests in North Africa, West and Central Asia: Challenges and Achievements. Curr. Opin. Insect Sci. 2021, 45, 35–41. [Google Scholar] [CrossRef]
- Togola, A.; Boukar, O.; Belko, N.; Chamarthi, S.K.; Fatokun, C.; Tamo, M.; Oigiangbe, N. Host plant resistance to insect pests of cowpea (Vigna unguiculata L. Walp.): Achievements and future prospects. Euphytica 2017, 213, 239. [Google Scholar] [CrossRef]
- Jacquot, M.; Massol, F.; Muru, D.; Derepas, B.; Deguine, J.P. Arthropod diversity is governed by bottom-up and top-down forces in a tropical agroecosystem. Agric. Ecosyst. Environ. 2019, 285, 106623. [Google Scholar] [CrossRef]
- Luo, K.; Zhao, H.; Wang, X.; Kang, Z. Prevalent Pest Management Strategies for Grain Aphids: Opportunities and Challenges. Front. Plant Sci. 2021, 12, 790919. [Google Scholar] [CrossRef]
- Yates, A.D.; Michel, A. Mechanisms of aphid adaptation to host plant resistance. Curr. Opin. Insect Sci. 2018, 26, 41–49. [Google Scholar] [CrossRef]
- Wellings, P.W.; Ward, S.A.; Dixon, A.F.G.; Rabbinge, R. Crop loss assessment. In Aphids, Their Biology, Natural Enemies and Control; Elsevier: Amsterdam, The Netherlands, 1989; Volume 2. [Google Scholar]
- Ryalls, J.M.W.; Riegler, M.; Moore, B.D.; Johnson, S.N. Biology and trophic interactions of lucerne aphids. Agric. For. Entomol. 2013, 15, 335–350. [Google Scholar] [CrossRef]
- Kamphuis, L.G.; Zulak, K.; Gao, L.-L.; Anderson, J.S. Plant-aphid interactions with a focus on legumes. J. Funct. Plant Biol. 2013, 40, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Nan, L.; Kevin, S. The Current Status, Problems, and Prospects of Alfalfa (Medicago sativa L.) Breeding in China. Agronomy 2017, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- Kataria, R.; Kumar, D. Population dynamics of Aphis craccivora (Koch) and its natural enemies on bean crop in relation to weather parameters in Vadodara, Gujarat, India. Legume Res. 2016, 40, 571–579. [Google Scholar] [CrossRef] [Green Version]
- John, G. Integrated Management of Sap Feeding Insects of Pulse Crops. Ann. Entomol. Soc. Am. 2018, 111, 172–183. [Google Scholar]
- Wilson, C.G.; Swincer, D.E.; Walden, K.J. The origins, distribution and host range of the spotted alfalfa aphid, Therioaphis trifolii (Monell) f. maculata, with a description of its spread in South Australia. J. Entomol. Soc. South. Afr. 1981, 44, 331–341. [Google Scholar]
- Klingler, J.P.; Nair, R.M.; Owain, R.E.; Singh, K.B. A single gene, AIN, in Medicago truncatula mediates a hypersensitive response to both bluegreen aphid and pea aphid, but confers resistance only to bluegreen aphid. J. Exp. Bot. 2009, 60, 4115–4127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klingler, J.; Creasy, R.; Gao, L.; Nair, R.M.; Calix, A.S.; Jacob, H.S.; Edwards, O.R.; Singh, K.B. Aphid resistance in Medicago truncatula involves antixenosis and phloem-specific, inducible antibiosis, and maps to a single locus flanked by NBS-LRR resistance gene analogs. Plant Physiol. 2005, 137, 1445–1455. [Google Scholar] [CrossRef] [Green Version]
- Guo, S.-M.; Kamphuis, L.G.; Gao, L.-L.; Klingler, J.P.; Lichtenzveig, J.; Edwards, O.; Singh, K.B. Identification of distinct quantitative trait loci associated with defence against the closely related aphids Acyrthosiphon pisum and A. kondoi in Medicago truncatula. J. Exp. Bot. 2012, 63, 3913–3922. [Google Scholar] [CrossRef] [Green Version]
- Klingler, J.P.; Edwards, O.R.; Singh, K.B. Independent action and contrasting phenotypes of resistance genes against spotted alfalfa aphid and bluegreen aphid in Medicago truncatula. New Phytol. 2010, 173, 630–640. [Google Scholar] [CrossRef]
- Xu, Y.; Guo, H.; Geng, G.; Zhang, Q.; Zhang, S. Changes in defense-related enzymes and phenolics in resistant and susceptible common wheat cultivars under aphid stress. Acta Physiol. Plant. 2021, 43, 36. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids on the world’s crops: An identification guide. Orient. Insects 1984, 35, 104. [Google Scholar]
- Ma, J.H.W.; Zhang, H.Y.; Zhang, R.; Li, K.C.; Luo, X.L.; Zhang, Y. Evaluation of resistance of main alfalfa varieties (lines) to Pea aphid in Ningxia. Acta Pratacult. Sin. 2016, 25, 190–197. [Google Scholar] [CrossRef]
- Dancewicz, K.; Gabry, B.; Morkunas, I.; Samardakiewicz, S. Probing behavior of Adelges laricis Vallot (Hemiptera: Adelgidae) on Larix decidua Mill: Description and analysis of EPG waveforms. PLoS ONE 2021, 16, e0251663. [Google Scholar] [CrossRef] [PubMed]
- FereresSaeed, A.I.; White, S.J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarjan, M.; Sturn, A.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar]
- Turcotte, M.M.; Lochab, A.K.; Turley, N.E.; Johnson, M.T.J. Plant domestication slows pest evolution. Ecol. Lett. 2015, 18, 907–915. [Google Scholar] [CrossRef]
- Birnbaum, S.; Abbot, P. Insect adaptations toward plant toxins in milkweed–herbivores systems—A review. J. Entomol. Exp. Appl. 2018, 166, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Wattier, C.; Turbant, A.; Sargos-Vallade, L.; Pelloux, J.; Rustérucci, C.; Cherqui, A. New insights into diet breadth of polyphagous and oligophagous aphids on two Arabidopsis ecotypes. Insect Sci. 2018, 26, 753–769. [Google Scholar] [CrossRef]
- Zhi, J.; Li, L.; Hou, X.; Xie, W.; Yue, W.; Zeng, G. Role of digestive enzymes in the adaptation of Frankliniella occidentalis to preferred and less-preferred host plants. Entomol. Exp. Appl. 2021, 169, 688–700. [Google Scholar] [CrossRef]
- Rigsby, C.M.; Showalter, D.N.; Herms, D.A.; Koch, J.L.; Bonello, P.; Cipollini, D. Physiological responses of emerald ash borer larvae to feeding on different ash species reveal putative resistance mechanisms and insect counter-adaptations. J. Insect Physiol. 2015, 78, 47–54. [Google Scholar] [CrossRef]
- Lucini, T.; Panizzi, A.R.; Bueno, A.d.F. Evaluating resistance of the soybean block technology cultivars to the Neotropical brown stink bug, Euschistus heros (F.). J. Insect Physiol. 2021, 131, 104–228. [Google Scholar] [CrossRef]
- Bansal, R.; Mian, M.; Michel, A. Characterizing Resistance to Soybean Aphid (Hemiptera: Aphididae): Antibiosis and Antixenosis Assessment. J. Econ. Entomol. 2021, 114, 1329–1335. [Google Scholar] [CrossRef] [PubMed]
- Natukunda, M.I.; Parmley, K.A.; Hohenstein, J.D.; Assefa, T.; Zhang, J.; MacIntosh, G.C.; Singh, A.K. Identification and Genetic Characterization of Soybean Accessions Exhibiting Antibiosis and Antixenosis Resistance to Aphis Glycines (Hemiptera: Aphididae). J. Econ. Entomol. 2019, 112, 1428–1438. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.; Tickes, B. Cowpea Aphid in Alfalfa; Yuma County Farm Notes; The University of Arizona Cooperative Extension: Tucson, AZ, USA, 2001. [Google Scholar]
- Smith, C.M. Plant Resistance to Insects: A Fundamental Approach; John Wiley & Sons: New York, NY, USA, 1989. [Google Scholar]
- Linn, P. Studies on the dispersal by upper winds of Aphis craccivora Koch. in New South Wales. Insect Sci. 1999, 19, 1–16. [Google Scholar]
- Rice, M. Cowpea Aphid: A New Pest in Iowa Alfalfa; Iowa State University Integrated Crop Management, Iowa State University Cooperative Extension: Ames, IA, USA, 2002; p. 103. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Cultivars | Source | Number | Cultivars | Source |
|---|---|---|---|---|---|
| 1 | Pegasis | Australia | 9 | Gonggong NO.1 | China |
| 2 | Eureka+ | Australia | 10 | Zhongmu NO.1 | China |
| 3 | Saiwo | Australia | 11 | Sardi 10 | Australia |
| 4 | Surprise | Canada | 12 | WL440 | America |
| 5 | Kehan | Canada | 13 | WL903 | America |
| 6 | Mufeng | China | 14 | WL656 | America |
| 7 | Algonquin | Canada | 15 | WL68 | America |
| 8 | Gannong NO.9 | China | 16 | WL525 | America |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Q.; Zhang, X.; Weng, X.; Gao, L.; Chang, X.; Wang, X.; Lu, Z. Identification and Characterization of Resistance of Three Aphid Species on Contrasting Alfalfa Cultivars. Insects 2022, 13, 530. https://doi.org/10.3390/insects13060530
Wu Q, Zhang X, Weng X, Gao L, Chang X, Wang X, Lu Z. Identification and Characterization of Resistance of Three Aphid Species on Contrasting Alfalfa Cultivars. Insects. 2022; 13(6):530. https://doi.org/10.3390/insects13060530
Chicago/Turabian StyleWu, Qiong, Xiang Zhang, Xianghao Weng, Lingling Gao, Xuefei Chang, Xingxing Wang, and Zhaozhi Lu. 2022. "Identification and Characterization of Resistance of Three Aphid Species on Contrasting Alfalfa Cultivars" Insects 13, no. 6: 530. https://doi.org/10.3390/insects13060530