
Soldier Caste-Specific Protein 1 Is Involved in Soldier Differentiation in Termite Reticulitermes aculabialis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and JH Treatment
2.2. Sample Preparing
2.3. cDNA Cloning and Homology Search
2.4. Real-Time Quantitative PCR
2.5. dsRNA Preparing
2.6. RNAi Experiment
2.7. Statistical Analysis
3. Results
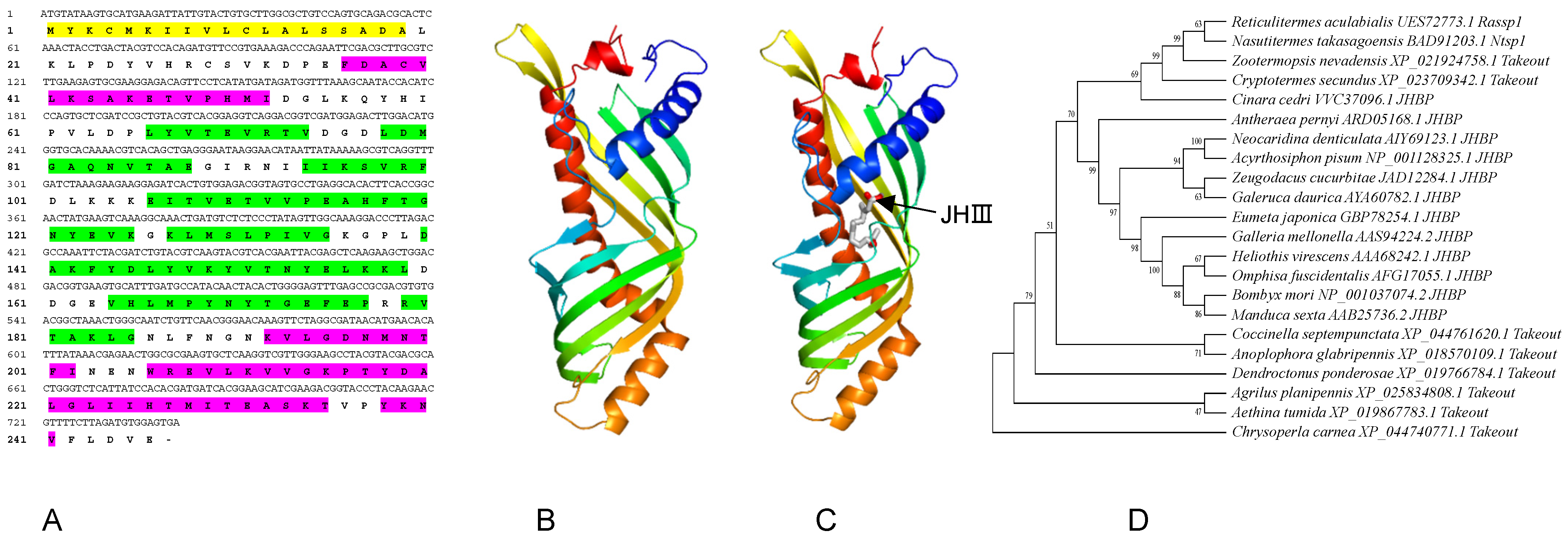
3.1. Characterization and Bioinformatics Analysis of RaSsp1
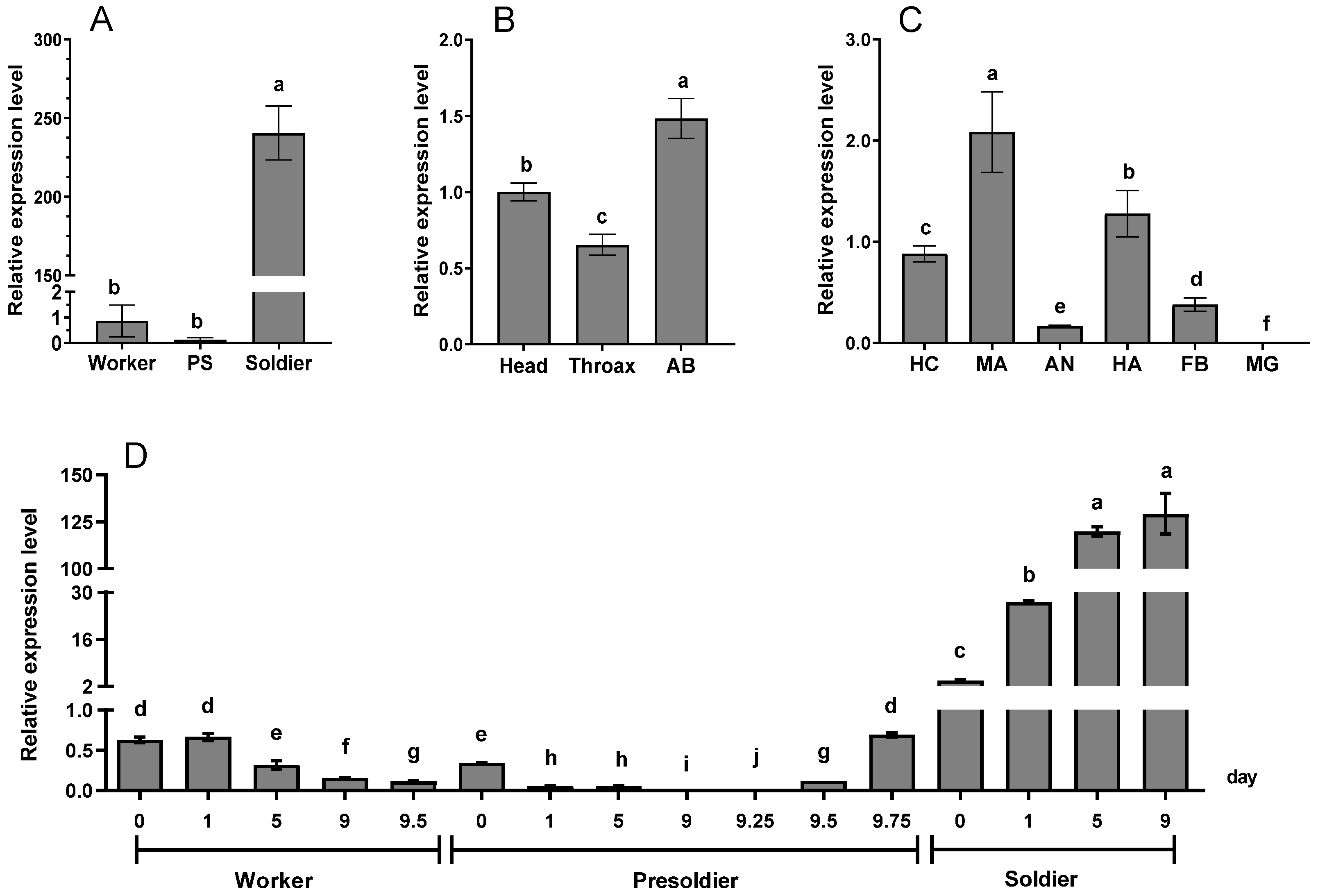
3.2. Temporospatial Expression Pattern of RaSsp1
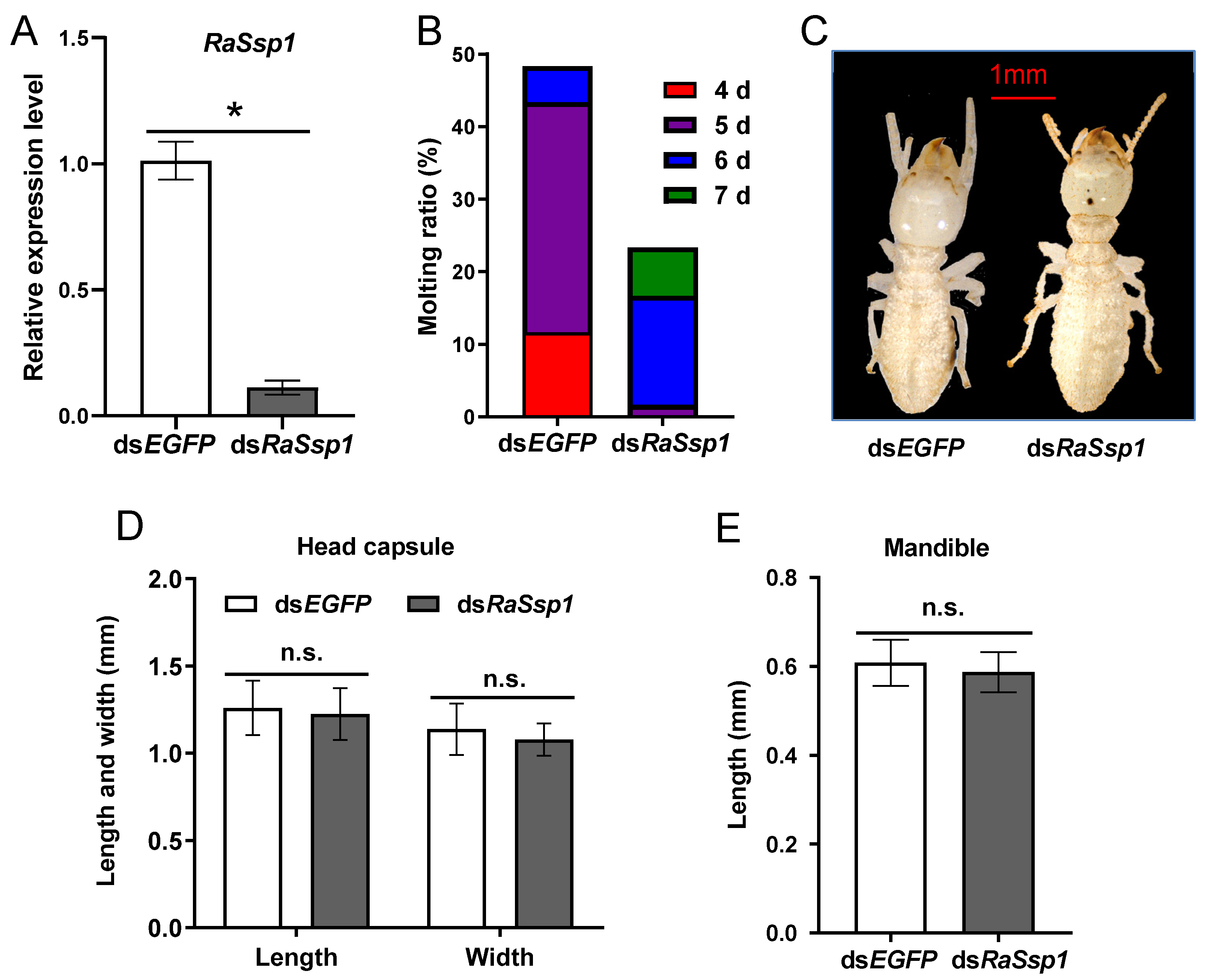
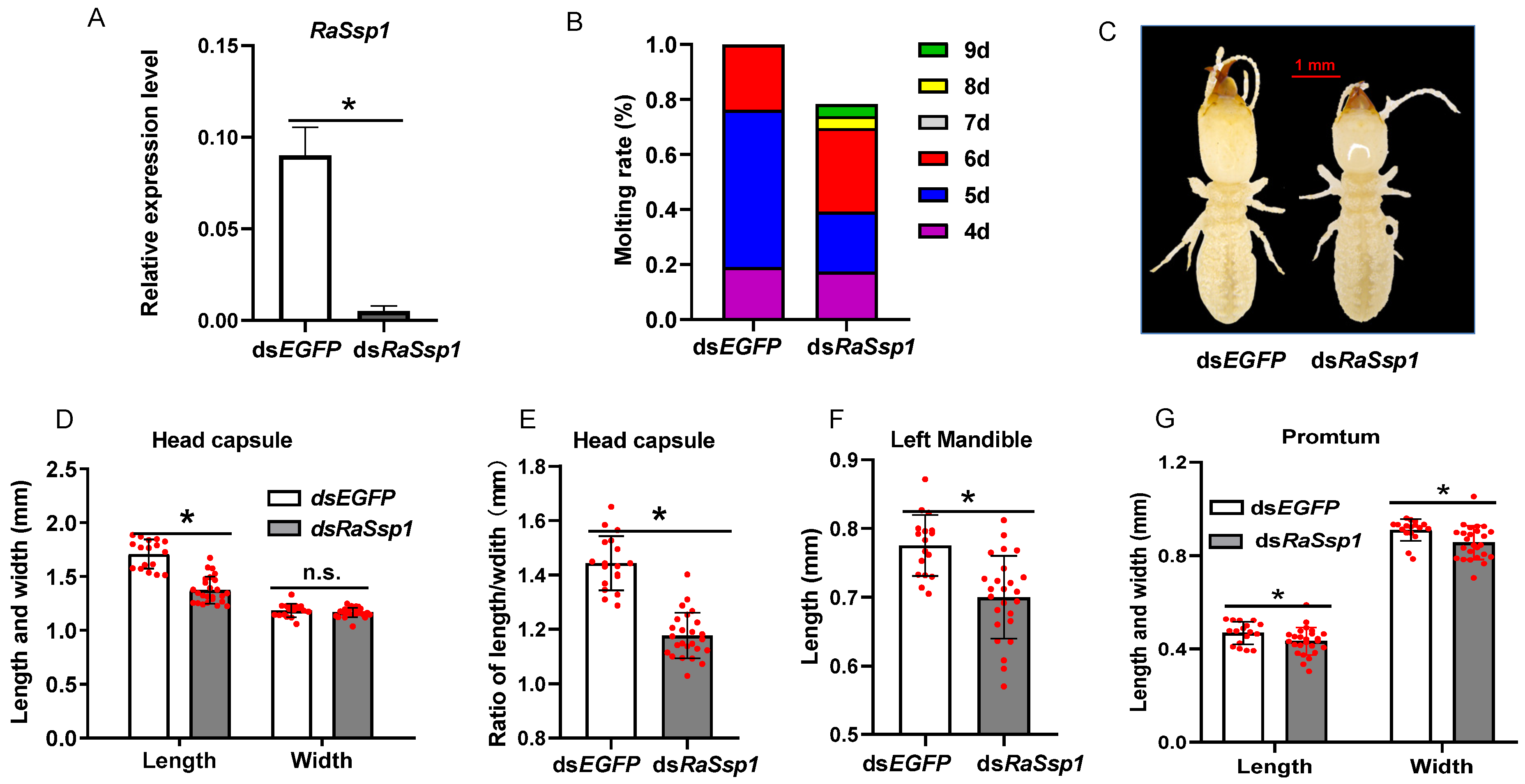
3.3. Effects of RaSsp1 RNAi on Presoldier Development before Worker Molting
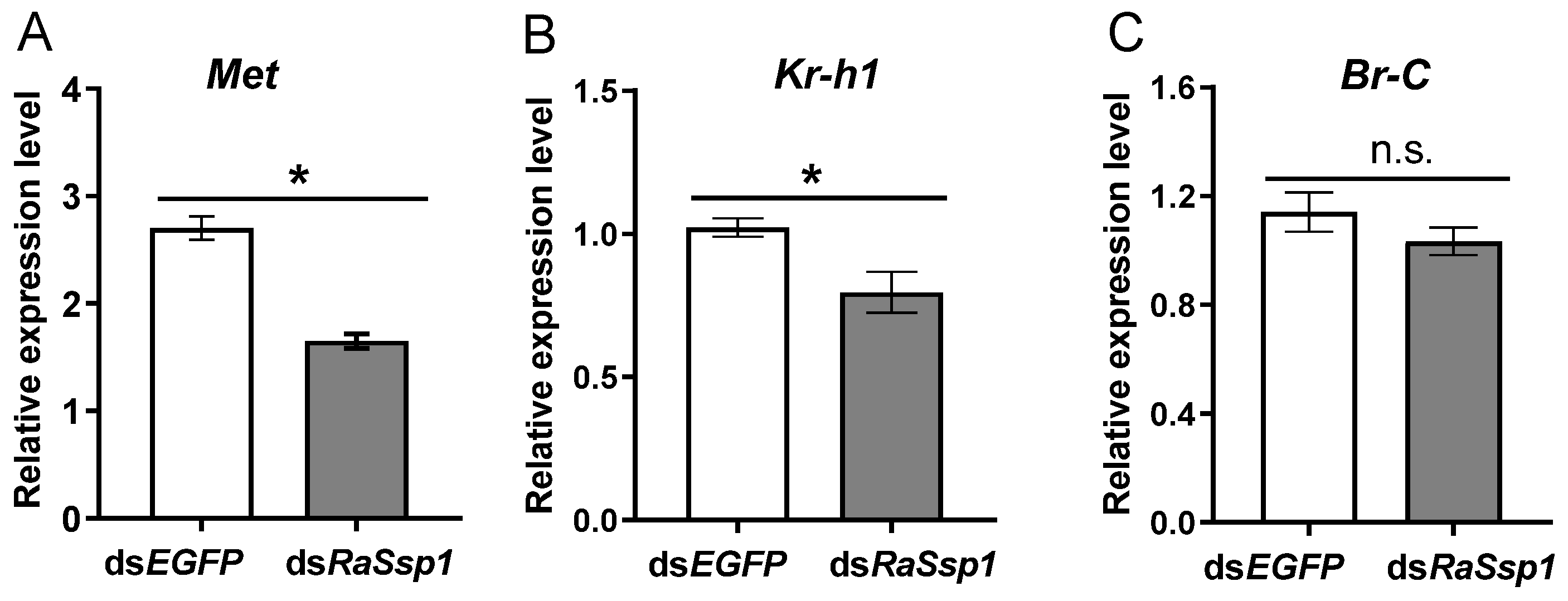
3.4. Effects of RaSsp1 RNAi on Soldier Development before Presoldier Molting
4. Discussion
4.1. Characteristic and Functional Prediction of RaSsp1
4.2. Temporospatial Expression Pattern of RaSsp1 in R. aculabialis
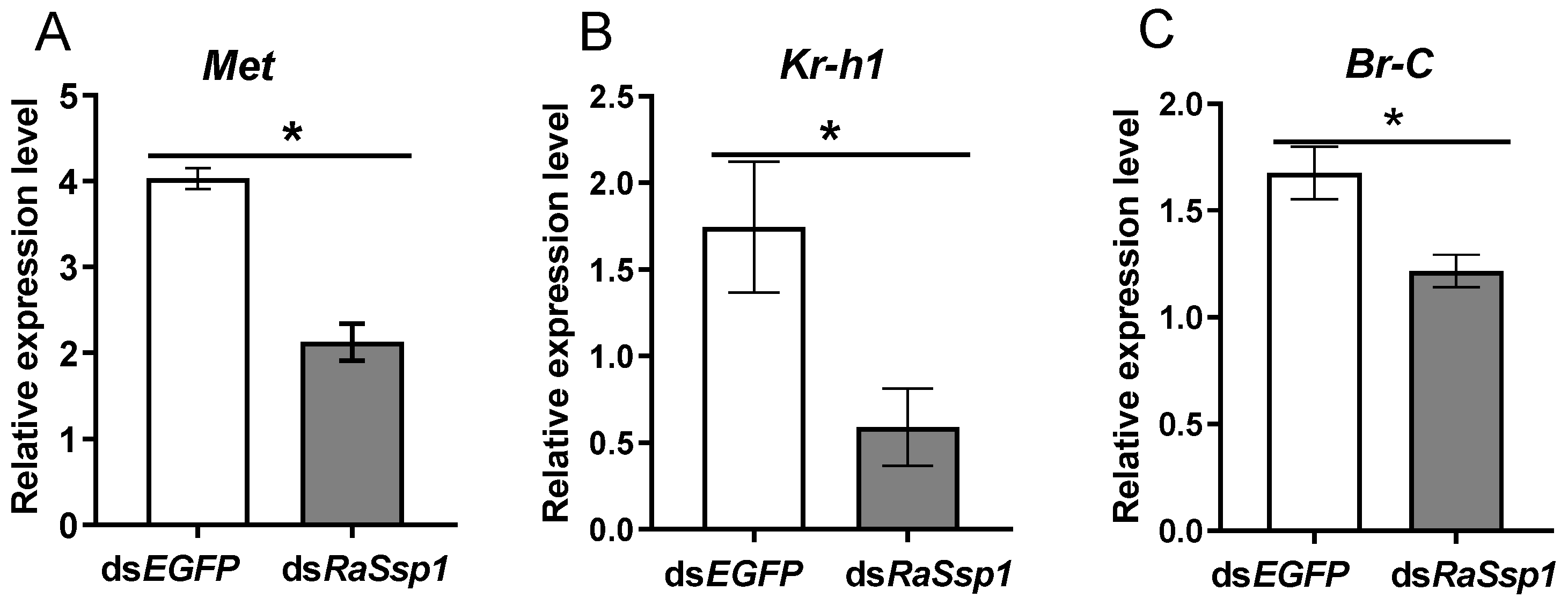
4.3. The Function of RaSsp1 in Soldier Differentiation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Koshikawa, S.; Matsumoto, T.; Miura, T. Morphometric changes during soldier differentiation of the damp-wood termite Hodotermopsis japonica (Isoptera, Termopsidae). Insectes Soc. 2022, 49, 245–250. [Google Scholar] [CrossRef]
- Miura, T.; Maekawa, K. The making of the defensive caste: Physiology, development, and evolution of the soldier differentiation in termites. Evol. Dev. 2020, 22, 425–437. [Google Scholar] [CrossRef] [PubMed]
- Nijhout, H.F. Development and evolution of adaptive polyphenisms. Evol. Dev. 2003, 5, 9–18. [Google Scholar] [CrossRef]
- Masuoka, Y.; Nuibe, K.; Hayase, N.; Oka, T.; Maekawa, K. Reproductive soldier development is controlled by direct physical interactions with reproductive and soldier termites. Insects 2021, 12, 76. [Google Scholar] [CrossRef]
- Hayashi, Y.; Lo, N.; Miyata, H.; Kitade, O. Sex-linked genetic influence on caste determination in a termite. Science 2007, 318, 985–987. [Google Scholar] [CrossRef] [PubMed]
- Miura, T. Developmental regulation of caste-specific characters in social-insect polyphenism. Evol. Dev. 2005, 7, 122–129. [Google Scholar] [CrossRef]
- Korb, J.; Hartfelder, K. Life history and development—A frame work for understanding developmental plasticity in lower termites. Biol. Rev. 2008, 83, 295–313. [Google Scholar] [CrossRef]
- Nijhout, H.F.; Wheeler, D.E. Juvenile hormone and the physiological basis of insect polyphenisms. Q. Rev. Biol. 1982, 57, 109–133. [Google Scholar] [CrossRef]
- Korb, J.; Hoffmann, K.; Hartfelder, K. Endocrine signatures underlying plasticity in postembryonic development of a lower termite, Cryptotermes secundus (Kalotermitidae). Evol. Dev. 2009, 11, 269–277. [Google Scholar] [CrossRef]
- Korb, J.; Greiner, C.; Foget, M.; Geiler, A. How can termites achieve their unparalleled postembryonic developmental plasticity? A test for the role of intermolt-specific high juvenile hormone titers. Front. Ecol. Evol. 2021, 9, 619594. [Google Scholar] [CrossRef]
- Cornette, R.; Gotoh, H.; Koshikawa, S.; Miura, T. Juvenile hormone titers and caste differentiation in the damp-wood termite Hodotermopsis sjostedti (Isoptera, Termopsidae). J. Insect Physiol. 2008, 54, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, H.; Ye, C.; Meng, Y.; Ran, Y.; Li, J.; Su, X. Comparative transcriptomic analysis andendocuticular protein gene expression of alate adults, workers and soldiers of the termite Reticulitermes aculabialis. BMC Genom. 2019, 20, 742. [Google Scholar] [CrossRef] [Green Version]
- Chu, J.; Wu, Z.; Du, Y.; Xi, Y.; An, S.; Su, L. Characteristics of soldier differentiation of termite Reticulitermes aculabialis induced by juvenile hormone and expression profiling of juvenile hormone-related genes. J. Plant Prot. 2020, 47, 1099–1107. [Google Scholar]
- Roisin, Y. Diversity and evolution of caste patterns. In Termites: Evolution, Symbioses, Ecology; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 95–119. [Google Scholar]
- Tsuchiya, M.; Watanabe, D.; Maekawa, K. Effect on mandibular length of juvenile hormones and regulation of soldier differentiation in the termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Appl. Entomol. Zool. 2008, 43, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, D.; Shirasaki, I.; Maekawa, K. Effects of juvenile hormone III on morphogenetic changes during a molt from each nymphal instar in the termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Appl. Entomol. Zool. 2010, 45, 377–386. [Google Scholar] [CrossRef]
- Masuoka, Y.; Yaguchi, H.; Suzuki, R.; Maekawa, K. Knockdown of the juvenile hormone receptor gene inhibits soldier-specific morphogenesis in the damp-wood termite Zootermopsis nevadensis (Isoptera: Archotermopsidae). Insect Biochem. Mol. Biol. 2015, 64, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Cesar, C.S.; Giacometti, D.; Costa-Leonardo, A.M.; Casarin, F.E. Drywood pest termite Cryptotermes brevis (Blattaria: Isoptera: Kalotermitidae): A detailed morphological study of pseudergates. Neotrop. Entomol. 2019, 48, 822–833. [Google Scholar] [CrossRef] [PubMed]
- Miura, T. Juvenile hormone as a physiological regulator mediating phenotypic plasticity in pancrustaceans. Dev. Growth Differ. 2018, 61, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Masuoka, Y.; Toga, K.; Nalepa, C.A.; Maekawa, K. A crucial caste regulation gene detected by comparing termites and sister group cockroaches. Genetics 2018, 209, 1225–1234. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Oi, F.M.; Scharf, M.E. Social exploitation of hexamerin: RNAi reveals a major caste-regulatory factor in termites. Proc. Natl. Acad. Sci. USA 2006, 103, 4499–4504. [Google Scholar] [CrossRef] [Green Version]
- Masuoka, Y.; Maekawa, K. Gene expression changes in the tyrosine metabolic pathway regulate caste-specific cuticular pigmentation of termites. Insect Biochem. Mol. Biol. 2016, 74, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Korb, J.; Belles, X. Juvenile hormone and hemimetabolan eusociality: A comparison of cockroaches with termites. Curr. Opin. Insect Sci. 2017, 22, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Ojani, R.; Fu, X.; Ahmed, T.; Liu, P.; Zhu, J. Krüppel homolog 1 acts as a repressor and an activator in the transcriptional response to juvenile hormone in adult mosquitoes. Insect Mol. Biol. 2018, 27, 268–278. [Google Scholar] [PubMed]
- Li, X.; Chen, T.; Jiang, H.; Huang, J.; Huang, M.; Xu, R.; Xie, Q.; Zhu, H.; Su, S. Effects of methyl farnesoate on Krüppel homolog 1 (Kr-h1) during vitellogenesis in the Chinese mitten crab (Eriocheir sinensis). Anim. Reprod. Sci. 2021, 224, 106653. [Google Scholar] [CrossRef] [PubMed]
- Kayukawa, T.; Nagamine, K.; Ito, Y.; Nishita, Y.; Ishikawa, Y.; Shinoda, T. Krüppel homolog 1 inhibits insect metamorphosis via direct transcriptional repression of Broad-Complex, a pupal specifier gene. J. Biol. Chem. 2016, 291, 1751–1762. [Google Scholar] [CrossRef] [Green Version]
- Hattori, A.; Sugime, Y.; Sasa, C.; Miyakawa, H.; Ishikawa, Y.; Miyazaki, S.; Okada, Y.; Cornette, R.; Lavine, L.C.; Emlen, D.J.; et al. Soldier morphogenesis in the damp-wood termite is regulated by the insulin signaling pathway. J. Exp. Zool. (Mol. Dev. Evol.) 2013, 320, 295–306. [Google Scholar] [CrossRef]
- Masuoka, Y.; Maekawa, K. Ecdysone signaling regulates soldier-specific cuticular pigmentation in the termite Zootermopsis nevadensis. FEBS Lett. 2016, 590, 1694–1703. [Google Scholar] [CrossRef]
- Masuoka, Y.; Yaguchi, H.; Toga, K.; Shigenobu, S.; Maekawa, K. TGFβ signaling related genes are involved in hormonal mediation during termite soldier differentiation. PLoS Genet. 2018, 14, e1007338. [Google Scholar] [CrossRef] [Green Version]
- Ye, C.; Song, Z.; Wu, T.; Zhang, W.; Saba, N.U.; Xing, L.; Su, X. Endocuticle is involved in caste differentiation of the lower termite. Curr. Zool. 2021, 67, 489–499. [Google Scholar] [CrossRef]
- Touhara, K.; Lerro, K.A.; Bonning, B.C.; Hammock, B.D.; Prestwich, G.D. Ligand binding by a recombinant insect juvenile hormone binding protein. Biochemistry 1993, 32, 2068–2075. [Google Scholar] [CrossRef]
- Dupas, S.; Neiers, F.; Granon, E.; Rougeux, E.; Dupont, S.; Beney, L.; Bousquet, F.; Shaik, H.A.; Briand, L.; Wojtasek, H.; et al. Collisional mechanism of ligand release by Bombyx mori JHBP, a member of the TULIP/Takeout family of lipid transporters. Insect Biochem. Mol. Biol. 2020, 117, 103293. [Google Scholar] [CrossRef] [PubMed]
- Touhara, K.; Prestwich, G.D. Binding site mapping of a photoaffinity-labeled juvenile hormone binding protein. Biochem. Biophys. Res. Commun. 1992, 182, 466–473. [Google Scholar] [CrossRef]
- Wojtasek, H.; Prestwich, G.D. Key disulfide bonds in an insect hormone binding protein: cDNA cloning of a juvenile hormone binding protein of Heliothis virescens and ligand binding by native and mutant forms. Biochemistry 1995, 34, 5234–5241. [Google Scholar] [CrossRef] [PubMed]
- Hojo, M.; Morioka, M.; Matsumoto, T.; Miura, T. Identification of soldier caste-specific protein in the frontal gland of nasute termite Nasutitermes takasagoensis (Isoptera: Termitidae). Insect Biochem. Mol. Bio. 2005, 35, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Zhu, S.; Ping, Z. Fauna Sinica Insecta. Vol. 17. Isoptera; Science Press: Beijing, China, 2000; Volume 17, pp. 341–346. [Google Scholar]
- Fujikawa, K.; Seno, K.; Ozaki, M. A novel Takeout-like protein expressed in the taste and olfactory organs of the blowfly, Phormia regina. FEBS J. 2006, 273, 4311–4321. [Google Scholar] [CrossRef] [PubMed]
- Saurabh, S.; Vanaphan, N.; Wen, W.; Dauwalder, B. High functional conservation of takeout family members in a courtship model system. PLoS ONE 2018, 13, e0204615. [Google Scholar] [CrossRef]
- Hamiaux, C.; Stanley, D.; Greenwood, D.R.; Baker, E.N.; Newcomb, R.D. Crystal structure of Epiphyas postvittana takeout 1 with bound ubiquinone supports a role as ligand carriers for takeout proteins in insects. J. Biol. Chem. 2009, 284, 3496–3503. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, Z.; Suzuki, R.; Shiotsuki, T.; Tsuchiya, W.; Tase, A.; Momma, M.; Yamazaki, T. Crystal structure of silkworm Bombyx mori JHBP in complex with 2-methyl-2,4-pentanediol: Plasticity of JH-binding pocket and ligand-induced conformational change of the second cavity in JHBP. PLoS ONE 2013, 8, e56261. [Google Scholar] [CrossRef]
- Suzuki, R.; Fujimoto, Z.; Shiotsuki, T.; Tsuchiya, W.; Momma, M.; Tase, A.; Miyazawa, M.; Yamazaki, T. Structural mechanism of JH delivery in hemolymph by JHBP of silkworm, Bombyx mori. Sci. Rep. 2011, 1, 133. [Google Scholar] [CrossRef] [Green Version]
- Benito, J.; Hoxha, V.; Lama, C.; Lazareva, A.A.; Ferveur, J.F.; Hardin, P.E.; Dauwalder, B. The circadian output gene takeout is regulated by Pdp1 epsilon. Proc. Natl. Acad. Sci. USA 2010, 107, 2544–2549. [Google Scholar] [CrossRef] [Green Version]
- Bauer, J.; Antosh, M.; Chang, C.; Schorl, C.; Kolli, S.; Neretti, N.; Helfand, S.L. Comparative transcriptional profiling identifies takeout as a gene that regulates life span. Aging 2010, 2, 298–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meunier, N.; Belgacem, Y.H.; Martin, J.R. Regulation of feeding behaviour and locomotor activity by takeout in Drosophila. J. Exp. Biol. 2007, 210, 1424–1434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Justice, R.W.; Dimitratos, S.; Walter, M.F.; Woods, D.F.; Biessmann, H. Sexual dimorphic expression of putative antennal carrier protein genes in the malaria vector Anopheles gambiae. Insect Mol. Biol. 2003, 12, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Alvarado, Y.; Rivera-Marchand, B. Worker queens? Behavioral flexibility of queens in the little fire ant Wasmannia auropunctata. Front. Ecol. Evol. 2020, 8, 2544–2549. [Google Scholar] [CrossRef]
- Schwinghammer, M.A.; Zhou, X.; Kambhampati, S.; Bennett, G.W.; Scharf, M.E. A novel gene from the takeout family involved in termite trail-following behavior. Gene 2011, 474, 12–21. [Google Scholar] [CrossRef]
- Park, Y.I.; Raina, A.K. Juvenile hormone III titers and regulation of soldier caste in Coptotermes formosanus (Isoptera: Rhinotermitidae). J. Insect Physiol. 2004, 50, 561–566. [Google Scholar] [CrossRef]
- Liu, Y.; Henderson, G.; Mao, L.; Laine, R.A. Seasonal variation of juvenile hormone titers of the formosan subterranean termite, Coptotermes formosanus (Rhinotermitidae). Environ. Entomol. 2005, 34, 557–562. [Google Scholar] [CrossRef]
- So, W.V.; Sarov-Blat, L.; Kotarski, C.K.; McDonald, M.J.; Allada, R.; Rosbash, M. takeout, a novel Drosophila gene under circadian clock transcriptional regulation. Mol. Cell Biol. 2000, 20, 6935–6944. [Google Scholar] [CrossRef] [Green Version]
- Sarov-Blat, L.; So, W.V.; Liu, L.; Rosbash, M. The Drosophila takeout gene is a novel molecular link between circadian rhythms and feeding behavior. Cell 2000, 101, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Dauwalder, B.; Tsujimoto, S.; Moss, J.; Mattox, W. The Drosophila takeout gene is regulated by the somatic sex-determination pathway and affects male courtship behavior. Genes Dev. 2002, 16, 2879–2892. [Google Scholar] [CrossRef] [Green Version]
- Prestwich, G.D. Interspecific variation in the defence secretions of Nasutitermes soldiers. Biochem. System. Ecol. 1979, 7, 211–221. [Google Scholar] [CrossRef]
- Goh, S.H.; Tong, S.L.; Tho, Y.P. Gas chromatography–mass spectrometry analysis of termite defense secretions in the subfamily Nasutitermitinae. Microchim. Acta 1982, 77, 219–229. [Google Scholar] [CrossRef]
- Everaerts, C.; Roisin, Y.; Le Quéré, J.L.; Bonnard, O.; Pasteels, J.M. Sesquiterpenes in the frontal gland secretions of nasute soldier termites from New Guinea. J. Chem. Ecol. 1993, 19, 2865–2879. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.A.; Costa-Leonardo, A.M. Anatomy of the frontal gland and ultramorphology of the frontal tube in the soldier caste of species of Nasutitermitinae (Isoptera, Termitidae). Microsc. Res. Tech. 2006, 69, 913–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, R.W.; Haverty, M.I. Termites and juvenile hormone analogues: A review of methodology and observed effects. Sociobiology 1979, 4, 269–278. [Google Scholar]
- Maekawa, K.; Hayashi, Y.; Lee, T.; Lo, N. Presoldier differentiation of Australian termite species induced by juvenile hormone analogs. Aust. Entomol. 2014, 53, 138–143. [Google Scholar] [CrossRef]
- Toga, K.; Hojo, M.; Miura, T.; Maekawa, K. Presoldier induction by juvenile hormone analogue in the nasute termite Nasutitermes takasagoensis (Isoptera: Termitidae). Zoolog. Sci. 2009, 26, 382–388. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, L.; Tang, Q.; Zhang, Y.; Tang, H.; Lü, P.; Yao, Q.; Chen, K. Comparative proteomic analysis reveals that juvenile hormone binding protein and adenylate kinase may be involved in the molting process of silkworm, Bombyx mori. Invertebr. Surviv. J. 2017, 14, 388–403. [Google Scholar]
- Ritdachyeng, E.; Manaboon, M.; Tobe, S.S.; Tippawan, S. Possible roles of Juvenile Hormone and Juvenile Hormone binding protein on changes in the integument during termination of larval diapause in the bamboo borer Omphisa fuscidentalis. Physiol. Entomol. 2013, 38, 183–191. [Google Scholar] [CrossRef]
- Ishikawa, Y.; Aonuma, H.; Miura, T. Soldier-Specific Modification of the Mandibular Motor Neurons in Termites. PLoS ONE 2008, 3, e2617. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Du, Y.; Li, Z.; Guo, R.; Li, Y.; Wei, J.; Yin, X.; Su, L. Soldier Caste-Specific Protein 1 Is Involved in Soldier Differentiation in Termite Reticulitermes aculabialis. Insects 2022, 13, 502. https://doi.org/10.3390/insects13060502
Wu Z, Du Y, Li Z, Guo R, Li Y, Wei J, Yin X, Su L. Soldier Caste-Specific Protein 1 Is Involved in Soldier Differentiation in Termite Reticulitermes aculabialis. Insects. 2022; 13(6):502. https://doi.org/10.3390/insects13060502
Chicago/Turabian StyleWu, Zhiwei, Yunliang Du, Zhenya Li, Ruiyao Guo, Yiying Li, Jizhen Wei, Xinming Yin, and Lijuan Su. 2022. "Soldier Caste-Specific Protein 1 Is Involved in Soldier Differentiation in Termite Reticulitermes aculabialis" Insects 13, no. 6: 502. https://doi.org/10.3390/insects13060502