Transcriptome Profiling Reveals Genes Related to Sex Determination and Differentiation in Sugarcane Borer (Chilo sacchariphagus Bojer)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
2.2. RNA Isolation, cDNA Library Preparation and Sequencing
2.3. De Novo Transcriptome Assembly and Annotation
2.4. Identification of Deferentially Expressed Genes (DEGs)
2.5. Protein–Protein Interaction (PPI) Network
2.6. Validation of the Gene Expression Level by qRT-PCR
3. Results
3.1. Transcriptome Analysis of the Male and Female Chilo sacchariphagus Bojer Adults
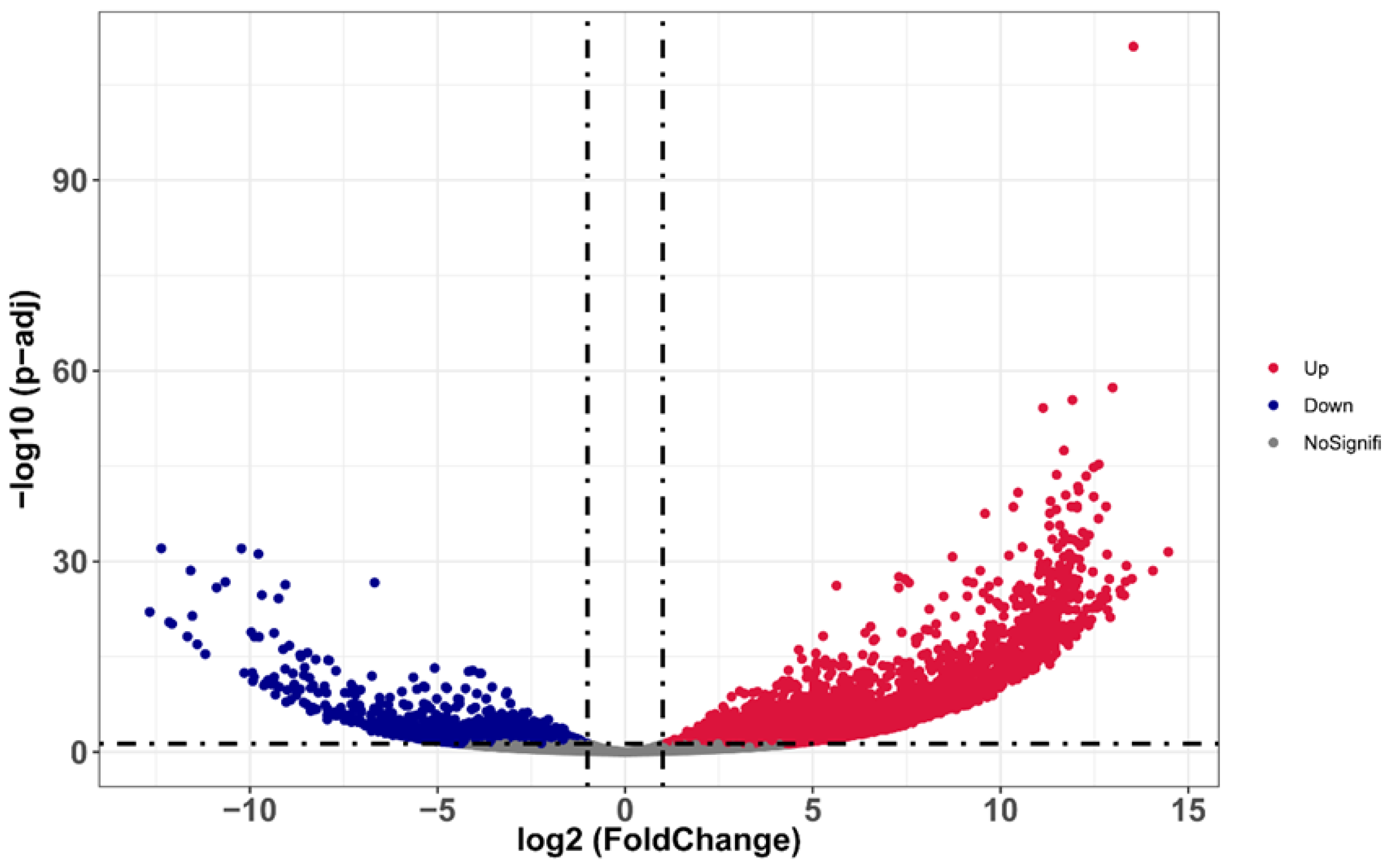
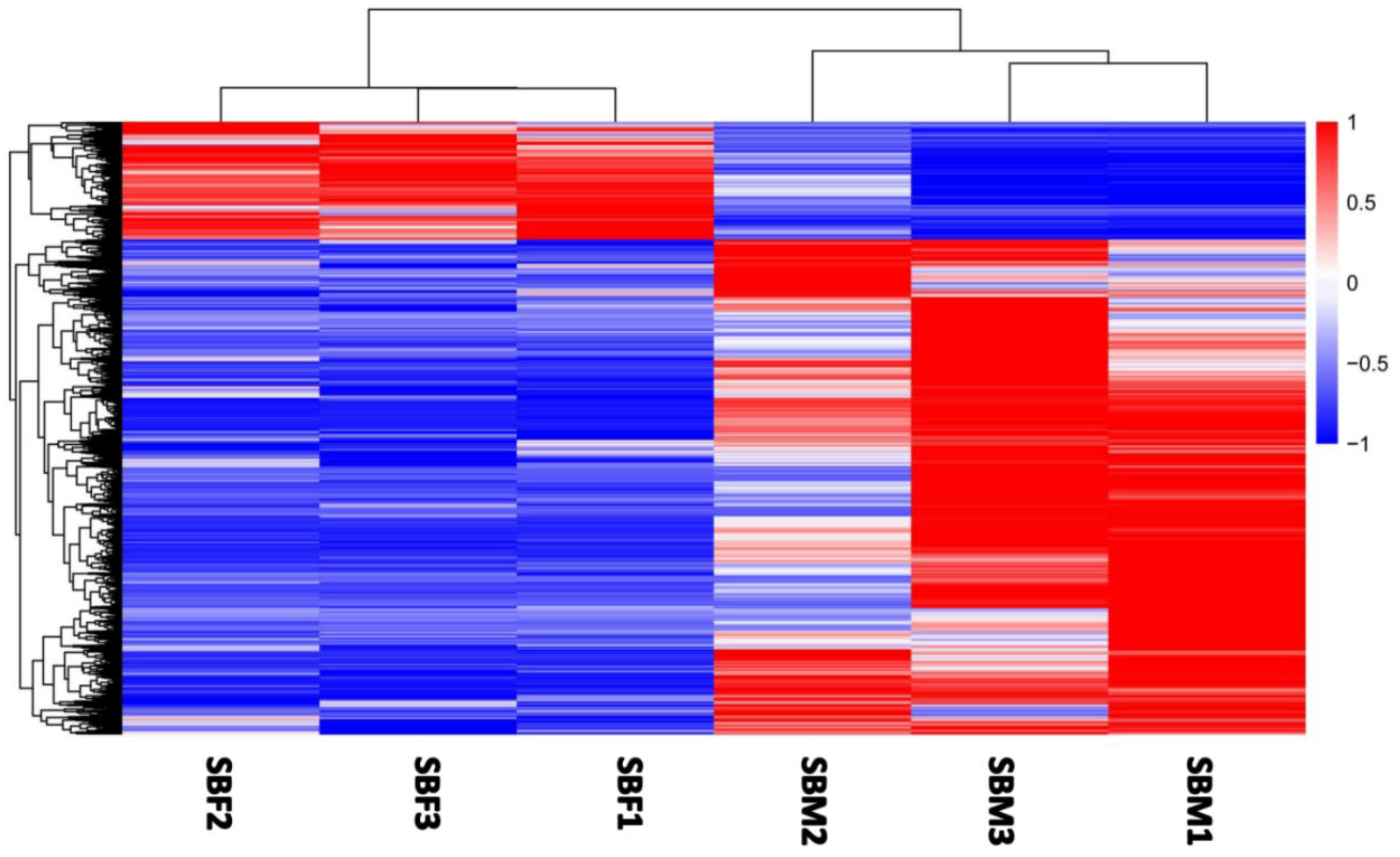
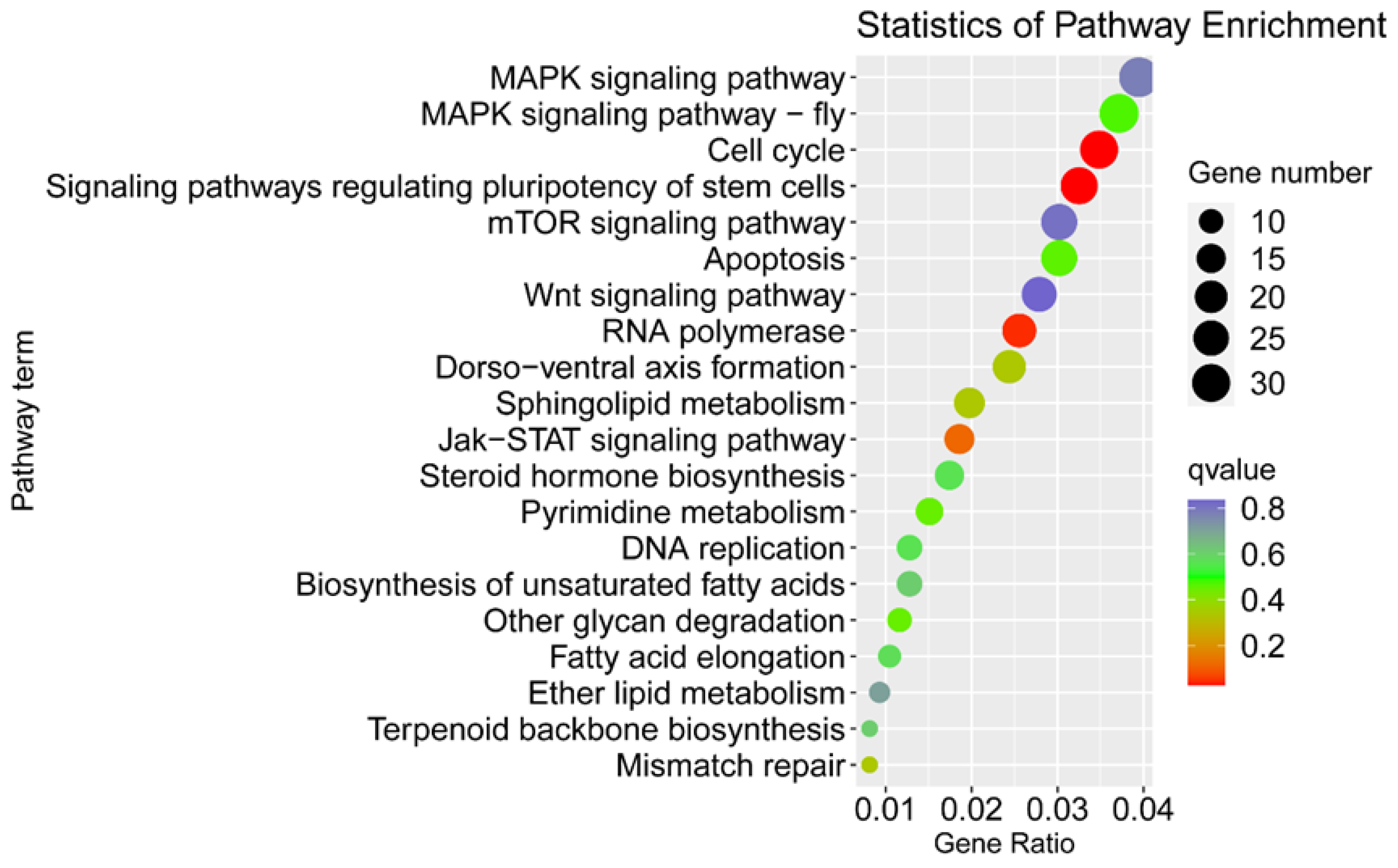
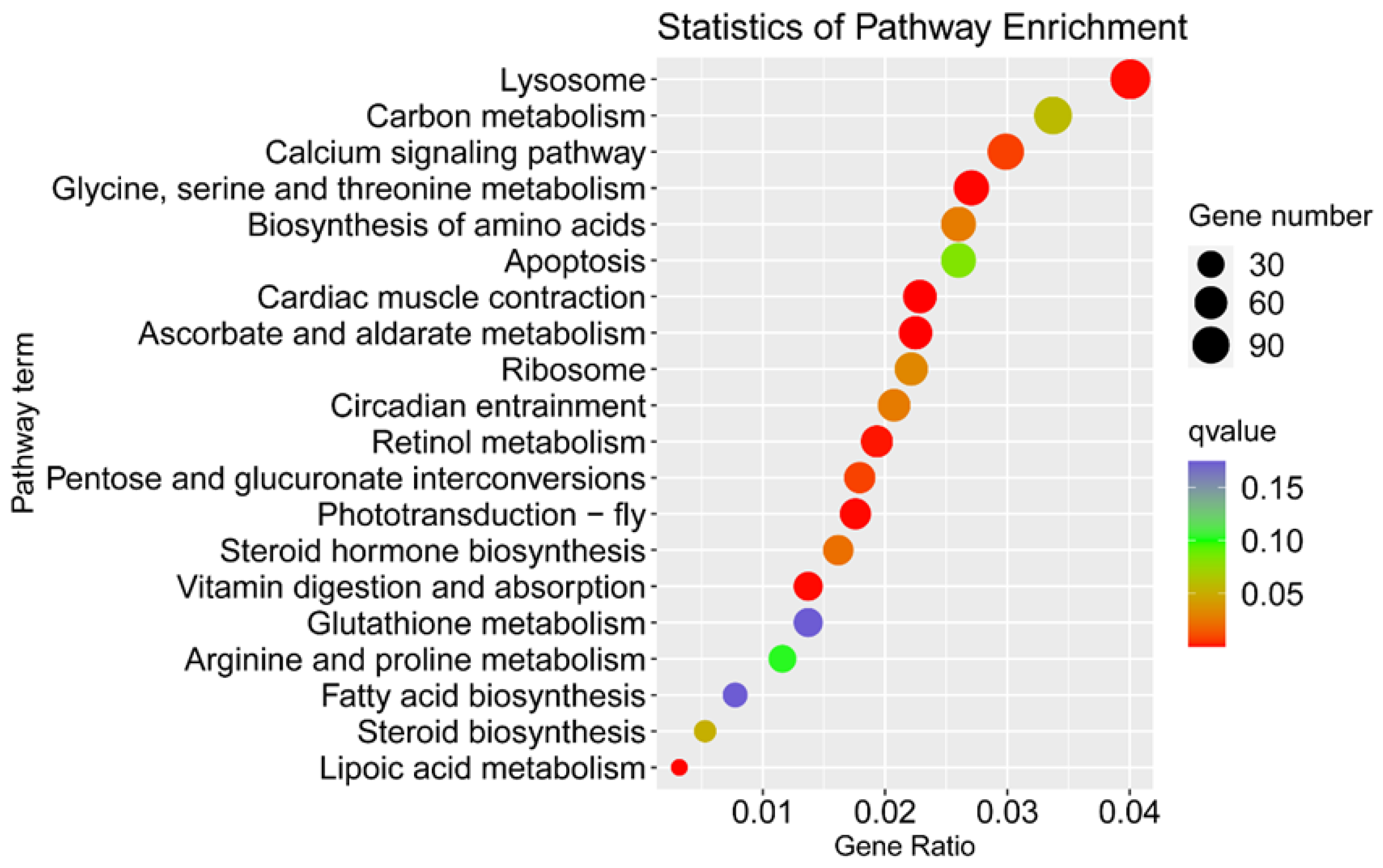
3.2. Identification of Sex-Biased Genes
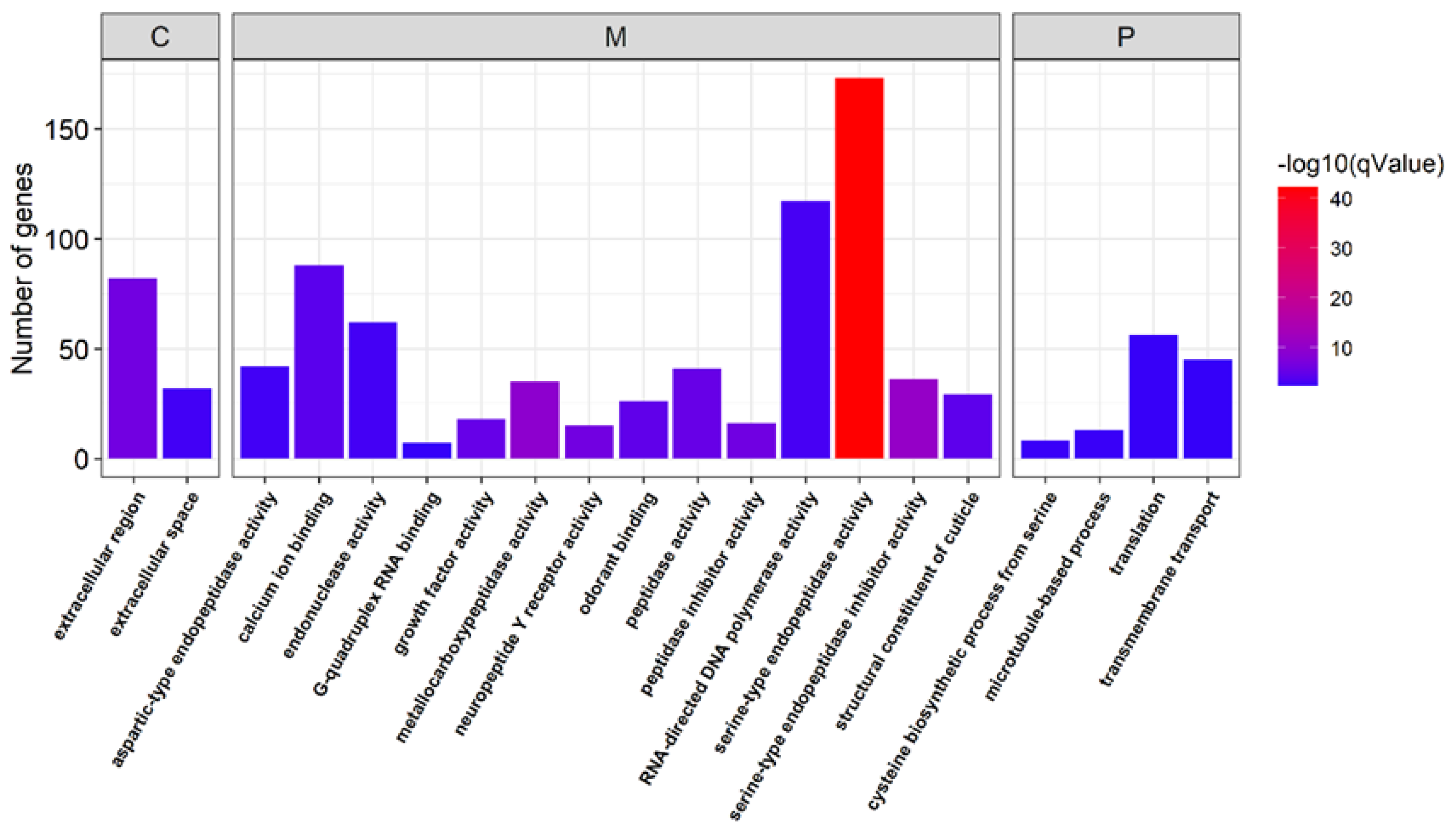
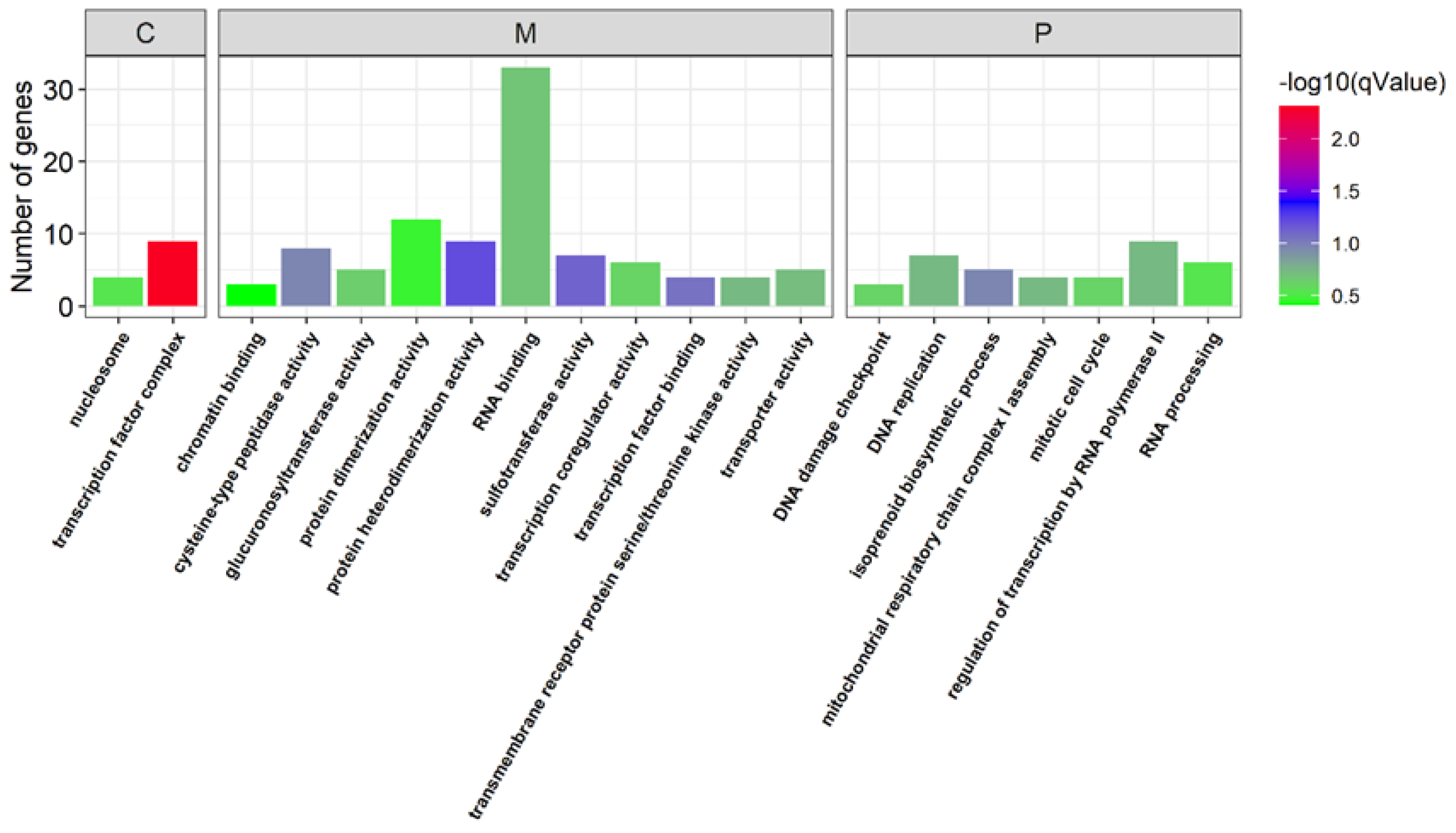
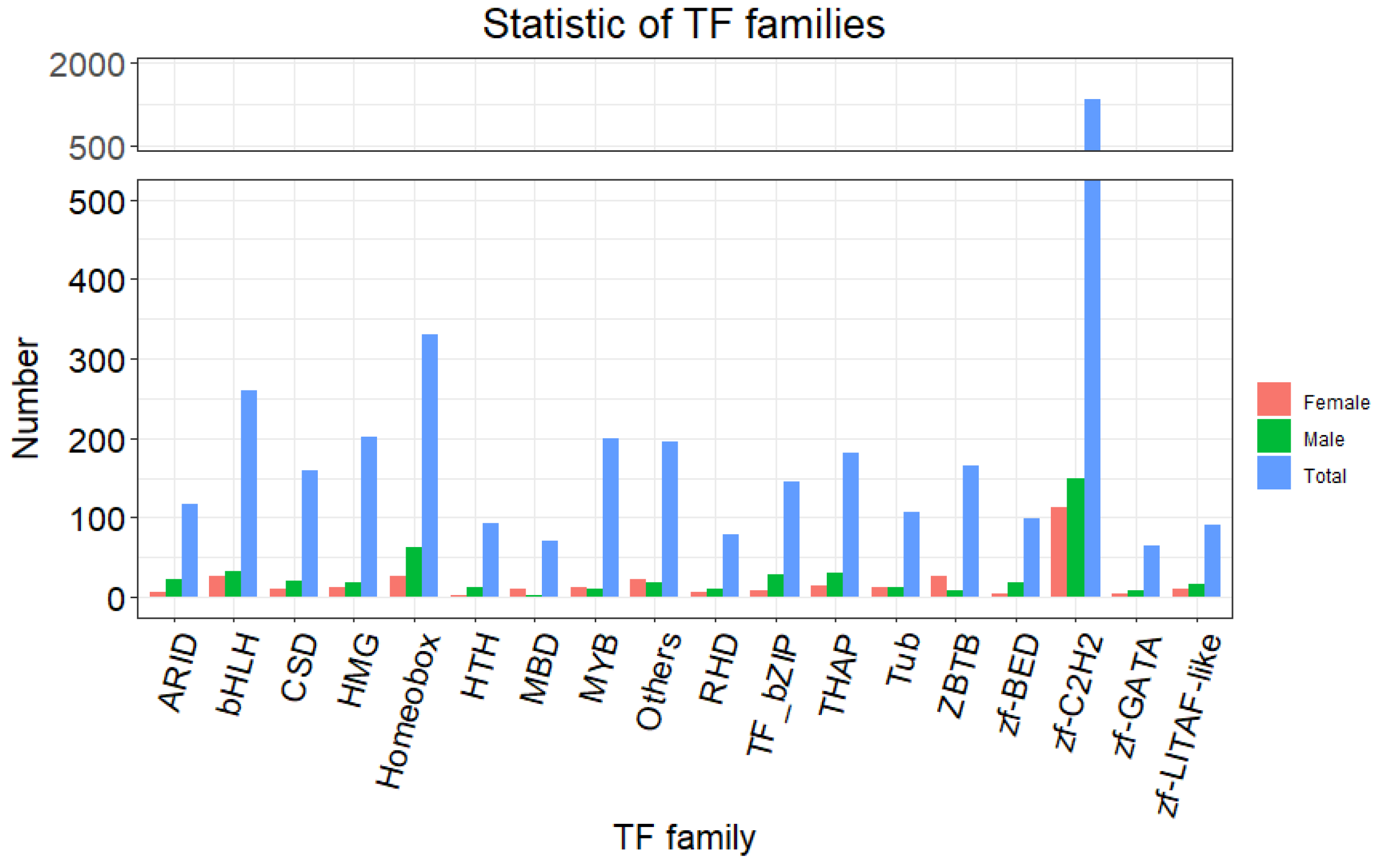
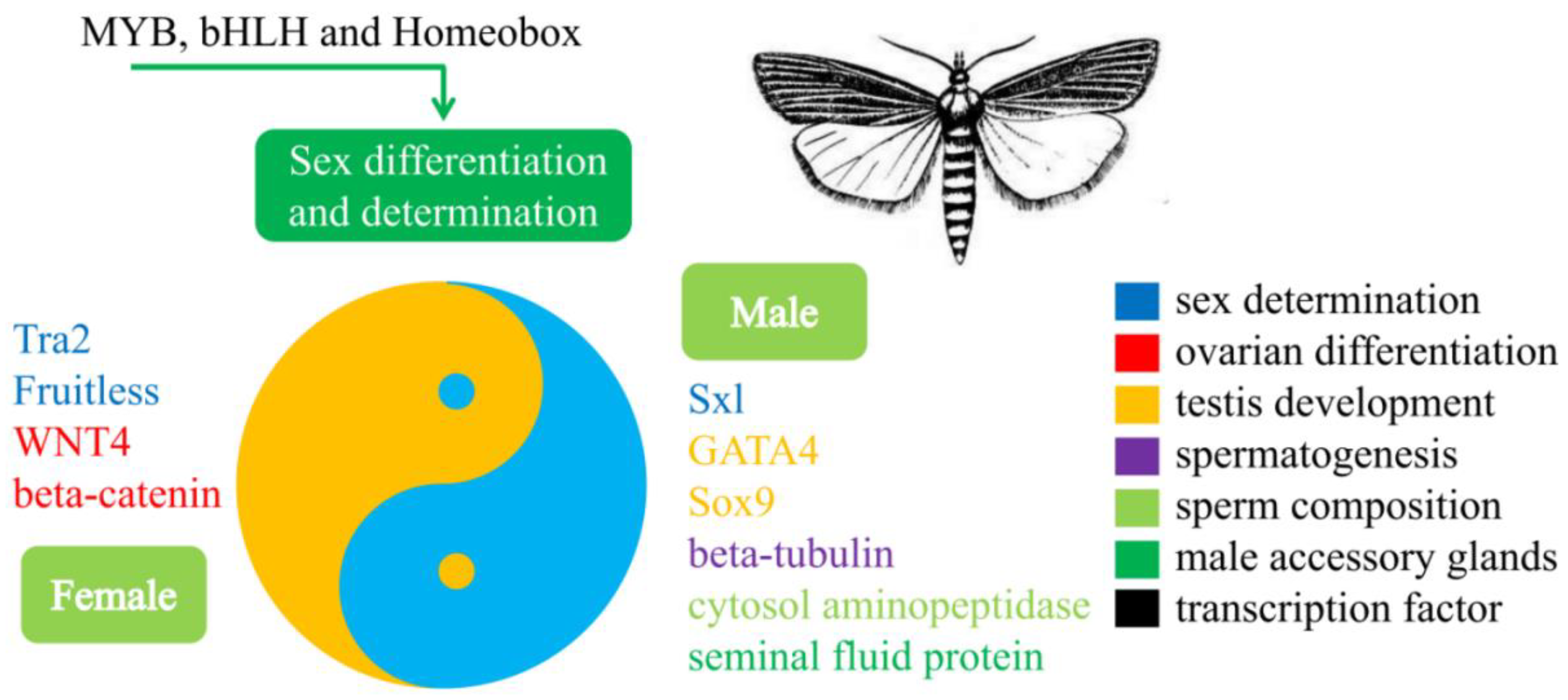
3.3. Identification of the Genes Related to Sex Determination and Differentiation
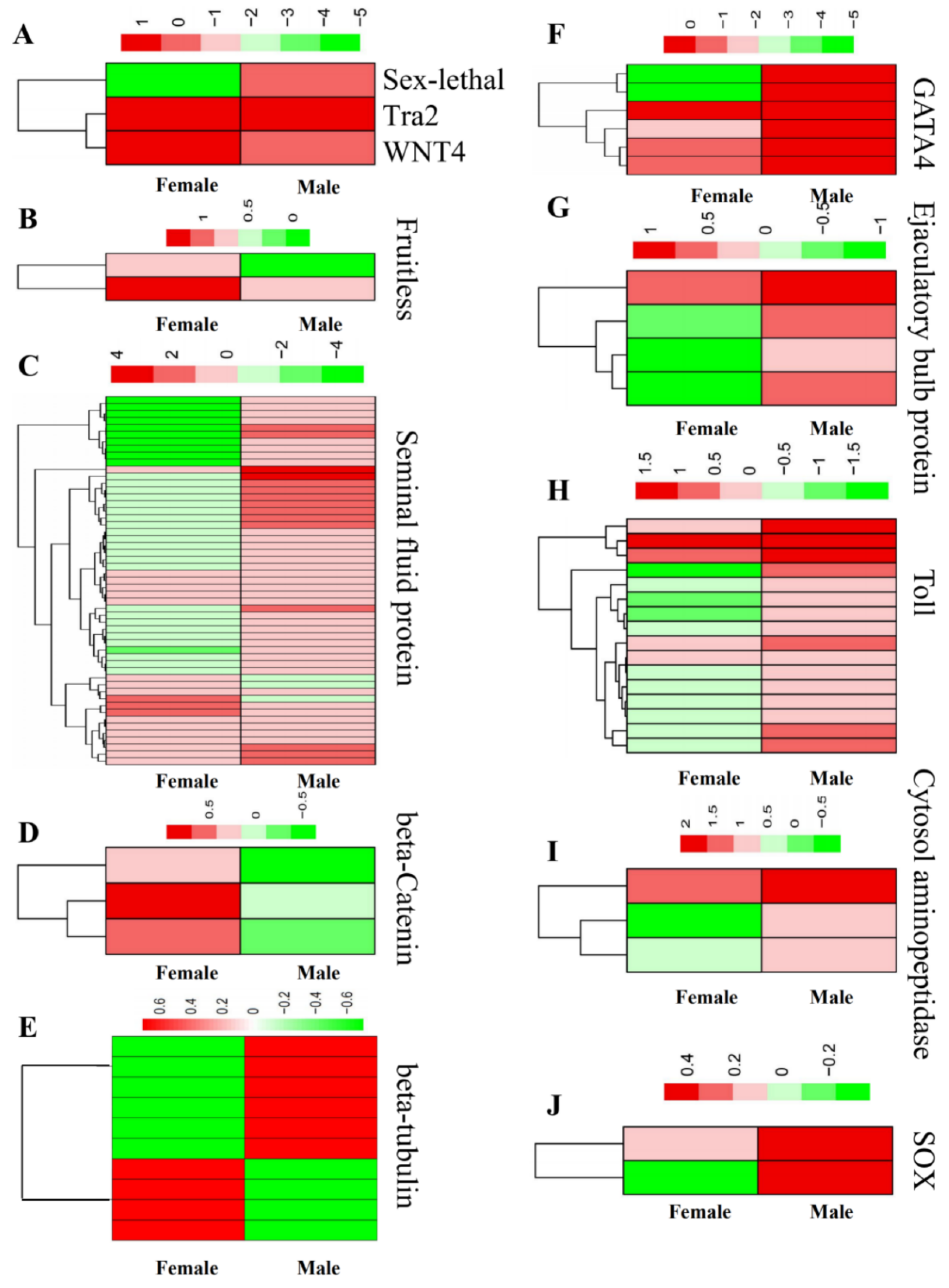
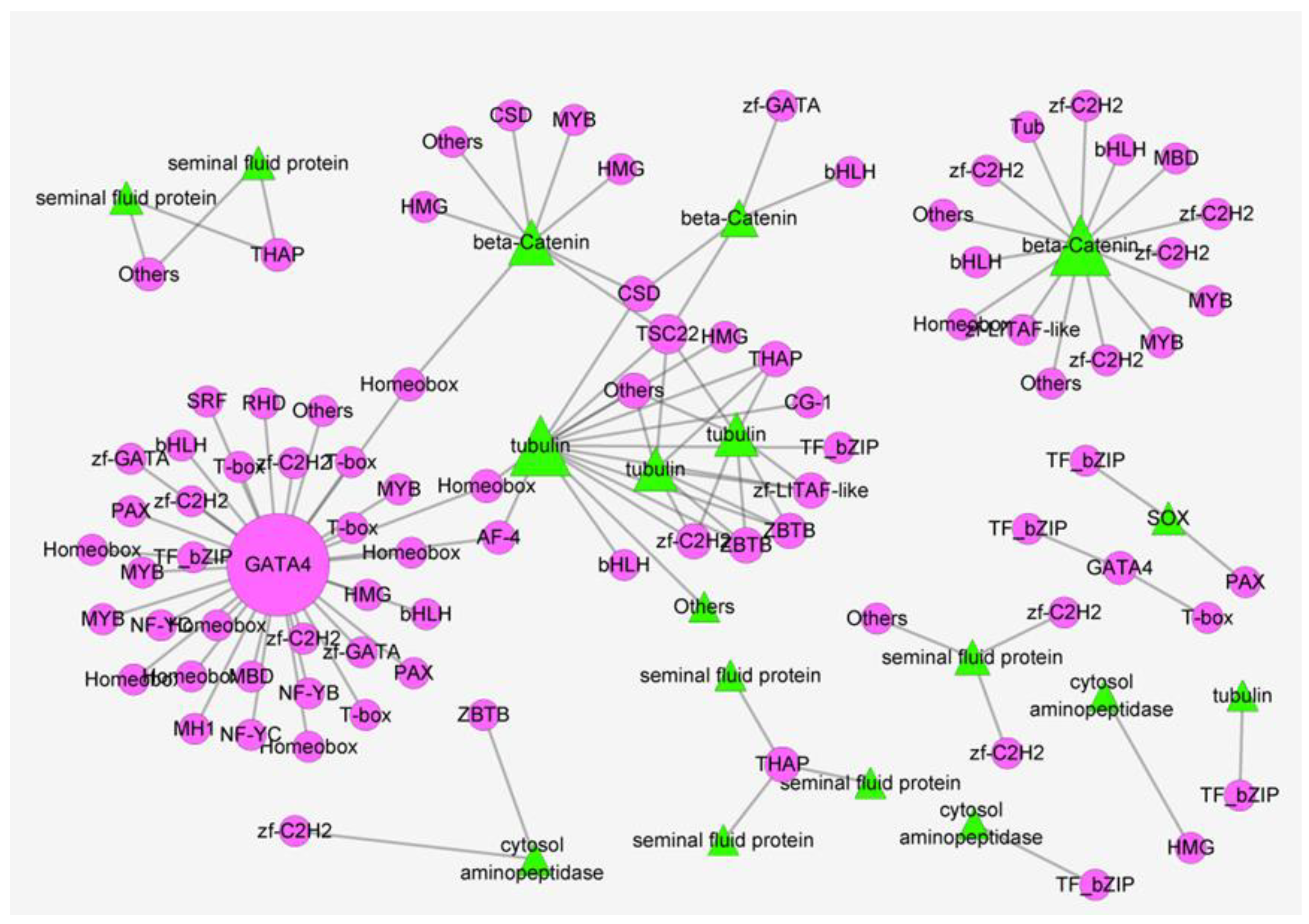
3.4. Construction of the Gene Network Involved in Sex Determination and Differentiation
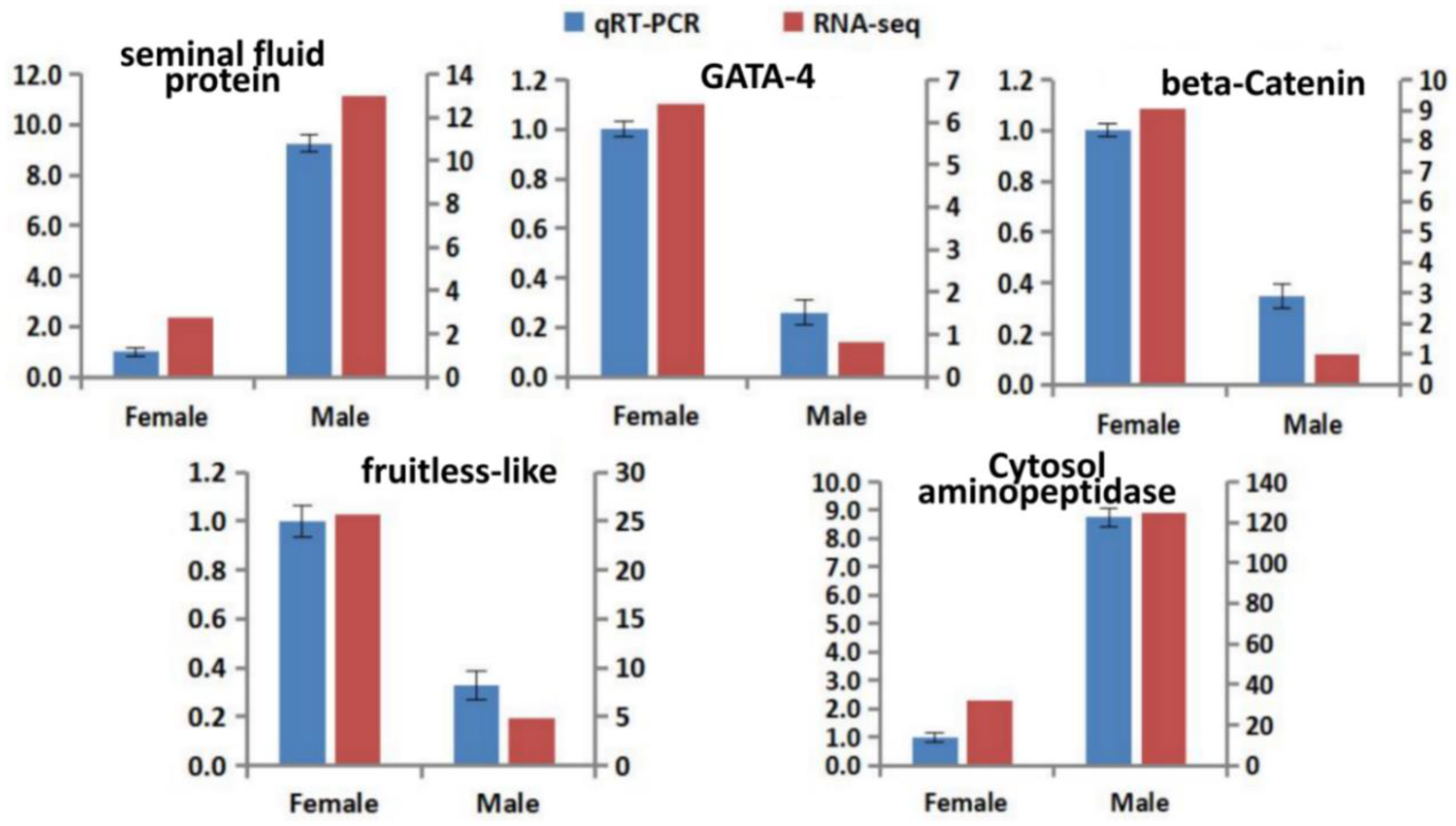
3.5. Verification of DEGs Using qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qin, Z.-Q.; Goebel, F.-R.; Li, D.-W.; Wei, J.-J.; Song, X.-P.; Luo, Y.-W.; Liu, L.; Deng, Z.-Y. Occurrence of Telenomus dignus (Gahan) on the Sugarcane Borers, Scirpophaga intacta Snellen and Chilo sacchariphagus Bojer in Guangxi Province, China. Sugar Tech 2018, 20, 725–729. [Google Scholar] [CrossRef]
- Cladera, J.L.; Vilardi, J.C.; Juri, M.; Paulin, L.E.; Giardini, M.C.; Cendra, P.V.G.; Segura, D.F.; Lanzavecchia, S.B. Genetics and biology of Anastrepha fraterculus: Research supporting the use of the sterile insect technique (SIT) to control this pest in Argentina. BMC Genom. Data 2014, 15, S12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plá, I.; de Oteyza, J.G.; Tur, C.; Martínez, M.; Dalmau, V. Sterile Insect Technique Programme against Mediterranean Fruit Fly in the Valencian Community (Spain). Insects 2021, 12, 415. [Google Scholar] [CrossRef]
- Hendrichs, J.; Franz, G.; Rendon, P. Increased effectiveness and applicability of the sterile insect technique through male-only releases for control of Mediterranean fruit flies during fruiting seasons. J. Appl. Entomol. 2010, 119, 371–377. [Google Scholar] [CrossRef]
- Thongsaiklaing, T.; Nipitwattanaphon, M.; Ngernsiri, L. The transformer2 gene of the pumpkin fruit fly, Bactrocera tau (Walker), functions in sex determination, male fertility and testis development. Insect Mol. Biol. 2018, 27, 766–779. [Google Scholar] [CrossRef] [PubMed]
- Tan, A.; Fu, G.; Jin, L.; Guo, Q.; Li, Z.; Niu, B.; Meng, Z.; Morrison, N.I.; Alphey, L.; Huang, Y. Transgene-based, female-specific lethality system for genetic sexing of the silkworm, Bombyx mori. Proc. Natl. Acad. Sci. USA 2013, 110, 6766–6770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marec, F.; Vreysen, M.J.B. Advances and Challenges of Using the Sterile Insect Technique for the Management of Pest Lepidoptera. Insects 2019, 10, 371. [Google Scholar] [CrossRef] [Green Version]
- Alphey, L. Re-engineering the sterile insect technique. Insect Biochem. Mol. Biol. 2002, 32, 1243–1247. [Google Scholar] [CrossRef]
- Degner, E.C.; Ahmed-Braimah, Y.; Borziak, K.; Wolfner, M.F.; Harrington, L.C.; Dorus, S. Proteins, Transcripts, and Genetic Architecture of Seminal Fluid and Sperm in the Mosquito Aedes aegypti. Mol. Cell. Proteom. 2018, 18, S6–S22. [Google Scholar] [CrossRef]
- Ellegren, H.; Parsch, J. The evolution of sex-biased genes and sex-biased gene expression. Nat. Rev. Genet. 2007, 8, 689–698. [Google Scholar] [CrossRef]
- Liu, P.-C.; Hao, D.-J.; Hu, H.-Y.; Wei, J.-R. Sexual dimorphism and sex-biased gene expression in an egg parasitoid species, Anastatus disparis. BMC Genom. 2020, 21, 492. [Google Scholar] [CrossRef]
- Parisi, M.; Nuttall, R.; Edwards, P.; Minor, J.; Naiman, D.; Lü, J.; Doctolero, M.; Vainer, M.; Chan, C.; Malley, J.; et al. A survey of ovary-, testis-, and soma-biased gene expression in Drosophila melanogaster adults. Genome Biol. 2004, 5, R40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, D.A.; Nolan, T.; Fischer, B.; Pinder, A.; Crisanti, A.; Russell, S. A comprehensive gene expression atlas of sex- and tissue-specificity in the malaria vector, Anopheles gambiae. BMC Genom. 2011, 12, 296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Z. Control of insect pests in sugarcane: Ipm approaches in China. Sugar Tech 2004, 6, 273–279. [Google Scholar]
- Tingjun, C. The Outbreak and Chemical Control of Sugarcane Borers. Sugarcane 1998, 5, 22–25. [Google Scholar]
- Jacob, V.; Tibère, R.; Nibouche, S. Few Sensory Cues Differentiate Host and Dead-End Trap Plant for the Sugarcane Spotted Borer Chilo sacchariphagus (Lepidoptera: Crambidae). J. Chem. Ecol. 2021, 47, 153–166. [Google Scholar] [CrossRef]
- Liu, J.; Liu, H.; Yi, J.; Mao, Y.; Li, J.; Sun, D.; An, Y.; Wu, H. Transcriptome Characterization and Expression Analysis of Chemosensory Genes in Chilo sacchariphagus (Lepidoptera Crambidae), a Key Pest of Sugarcane. Front. Physiol. 2021, 12, 636353. [Google Scholar] [CrossRef]
- Kfir, R.; Overholt, W.A.; Khan, Z.R.; Polaszek, A. Biology and Management of Economically Important Lepidopteran Cereal Stem Borers in Africa. Annu. Rev. Entomol. 2002, 47, 701–731. [Google Scholar] [CrossRef]
- Dyck, V.A.; Hendrichs, J.P.; Robinson, A.S. Sterile Insect Technique Principles and Practice in Area-Wide Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Sanchez, L. Sex-determining mechanisms in insects. Int. J. Dev. Biol. 2008, 52, 837–856. [Google Scholar] [CrossRef] [Green Version]
- Sakai, H.; Oshima, H.; Yuri, K.; Gotoh, H.; Daimon, T.; Yaginuma, T.; Sahara, K.; Niimi, T. Dimorphic sperm formation by Sex-lethal. Proc. Natl. Acad. Sci. USA 2019, 116, 10412–10417. [Google Scholar] [CrossRef] [Green Version]
- Meccariello, A.; Salvemini, M.; Primo, P.; Hall, B.; Koskinioti, P.; Dalíková, M.; Gravina, A.; Gucciardino, M.A.; Forlenza, F.; Gregoriou, M.-E.; et al. Maleness-on-the-Y (MoY) orchestrates male sex determination in major agricultural fruit fly pests. Science 2019, 365, 1457–1460. [Google Scholar] [CrossRef] [PubMed]
- Amrein, H.; Maniatis, T.; Nöthiger, R. Alternatively spliced transcripts of the sex-determining gene tra-2 of Drosophila encode functional proteins of different size. EMBO J. 1990, 9, 3619–3629. [Google Scholar] [CrossRef] [PubMed]
- Hedley, M.L.; Maniatis, T. Sex-specific splicing and polyadenylation of dsx pre-mRNA requires a sequence that binds specifically to tra-2 protein in vitro. Cell 1991, 65, 579–586. [Google Scholar] [CrossRef]
- Salvemini, M.; Robertson, M.; Aronson, B.; Atkinson, P.; Polito, L.C.; Saccone, G. Ceratitis capitata transformer-2 gene is required to establish and maintain the autoregulation of Cctra, the master gene for female sex determination. Int. J. Dev. Biol. 2009, 53, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Verhulst, E.C.; van de Zande, L.; Beukeboom, L.W. Insect sex determination: It all evolves around transformer. Curr. Opin. Genet. Dev. 2010, 20, 376–383. [Google Scholar] [CrossRef] [Green Version]
- Rideout, E.; Billeter, J.-C.; Goodwin, S.F. The Sex-Determination Genes fruitless and doublesex Specify a Neural Substrate Required for Courtship Song. Curr. Biol. 2007, 17, 1473–1478. [Google Scholar] [CrossRef] [Green Version]
- Shirangi, T.R.; Taylor, B.J.; McKeown, M. A double-switch system regulates male courtship behavior in male and female Drosophila melanogaster. Nat. Genet. 2006, 38, 1435–1439. [Google Scholar] [CrossRef]
- Hopkins, B.R.; Kopp, A. Evolution of sexual development and sexual dimorphism in insects. Curr. Opin. Genet. Dev. 2021, 69, 129–139. [Google Scholar] [CrossRef]
- Gempe, T.; Beye, M. Function and evolution of sex determination mechanisms, genes and pathways in insects. Bioessays 2011, 33, 52–60. [Google Scholar] [CrossRef] [Green Version]
- Kiuchi, T.; Koga, H.; Kawamoto, M.; Shoji, K.; Sakai, H.; Arai, Y.; Ishihara, G.; Kawaoka, S.; Sugano, S.; Shimada, T.; et al. A single female-specific piRNA is the primary determiner of sex in the silkworm. Nature 2014, 509, 633–636. [Google Scholar] [CrossRef]
- Lee, G.-S.; He, Y.; Dougherty, E.J.; Jimenez-Movilla, M.; Avella, M.; Grullon, S.; Sharlin, D.S.; Guo, C.; Blackford, J.A.; Awasthi, S.; et al. Disruption of Ttll5/Stamp Gene (Tubulin Tyrosine Ligase-like Protein 5/SRC-1 and TIF2-associated Modulatory Protein Gene) in Male Mice Causes Sperm Malformation and Infertility. J. Biol. Chem. 2013, 288, 15167–15180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khatun, A.; Kang, K.-H.; Ryu, D.-Y.; Rahman, S.; Kwon, W.-S.; Pang, M.-G. Effect of Aminopeptidase N on functions and fertility of mouse spermatozoa in vitro. Theriogenology 2018, 118, 182–189. [Google Scholar] [CrossRef] [PubMed]
- King, M.; Eubel, H.; Millar, A.H.; Baer, B. Proteins within the seminal fluid are crucial to keep sperm viable in the honeybee Apis mellifera. J. Insect Physiol. 2011, 57, 409–414. [Google Scholar] [CrossRef] [PubMed]
- Kemphues, K.J.; Kaufman, T.C.; Raff, R.A.; Raff, E.C. The testis-specific β-tubulin subunit in Drosophila melanogaster has multiple functions in spermatogenesis. Cell 1982, 31, 655–670. [Google Scholar] [CrossRef]
- Kemphues, K.J.; Raff, E.C.; Raff, R.A.; Kaufman, T.C. Mutation in a testis-specific β-tubulin in Drosophila: Analysis of its effects on meiosis and map location of the gene. Cell 1980, 21, 445–451. [Google Scholar] [CrossRef]
- Gillott, C. Male Accessory Gland Secretions: Modulators of Female Reproductive Physiology and Behavior. Annu. Rev. Entomol. 2003, 48, 163–184. [Google Scholar] [CrossRef]
- Huang, D.-L.; Gao, Y.-J.; Gui, Y.-Y.; Chen, Z.-L.; Qin, C.-X.; Wang, M.; Liao, Q.; Yang, L.-T.; Li, Y.-R. Transcriptome of High-Sucrose Sugarcane Variety GT35. Sugar Tech 2016, 18, 520–528. [Google Scholar] [CrossRef]
- Chen, Z.; Qin, C.; Wang, M.; Liao, F.; Liao, Q.; Liu, X.; Li, Y.; Lakshmanan, P.; Long, M.; Huang, D. Ethylene-mediated improvement in sucrose accumulation in ripening sugarcane involves increased sink strength. BMC Plant Biol. 2019, 19, 285. [Google Scholar] [CrossRef]
- Wang, M.; Li, A.; Liao, F.; Qin, C.; Chen, Z.; Zhou, L.; Li, Y.; Li, X.; Lakshmanan, P.; Huang, D. Control of sucrose accumulation in sugarcane (Saccharum spp. hybrids) involves miRNA-mediated regulation of genes and transcription factors associated with sugar metabolism. GCB Bioenergy 2022, 14, 173–191. [Google Scholar] [CrossRef]
- Wu, Q.; Xu, L.; Guo, J.; Su, Y.; Que, Y. Transcriptome Profile Analysis of Sugarcane Responses to Sporisorium scitaminea Infection Using Solexa Sequencing Technology. BioMed Res. Int. 2013, 2013, 298920. [Google Scholar] [CrossRef] [Green Version]
- Huang, N.; Ling, H.; Su, Y.; Liu, F.; Xu, L.; Su, W.; Wu, Q.; Guo, J.; Gao, S.; Que, Y. Transcriptional analysis identifies major pathways as response components to Sporisorium scitamineum stress in sugarcane. Gene 2018, 678, 207–218. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, S.; Su, Y.; Lin, Z.; Guo, J.; Li, M.; Wang, Z.; Que, Y.; Xu, L. Transcripts and low nitrogen tolerance: Regulatory and metabolic pathways in sugarcane under low nitrogen stress. Environ. Exp. Bot. 2019, 163, 97–111. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhong, W.; He, W.; Li, Y.; Li, Y.; Li, T.; Vasseur, L.; You, M. Genome-wide profiling of the alternative splicing provides insights into development in Plutella xylostella. BMC Genom. 2019, 20, 463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrella, V.; Aceto, S.; Colonna, V.; Saccone, G.; Sanges, R.; Polanska, N.; Volf, P.; Gradoni, L.; Bongiorno, G.; Salvemini, M. Identification of sex determination genes and their evolution in Phlebotominae sand flies (Diptera, Nematocera). BMC Genom. 2019, 20, 522. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Xie, W.; Guo, L.; Yang, X.; Yang, J.; Wang, S.; Wu, Q.; Zhou, X.; Zhang, Y. Genome-wide dissection of sex determination genes in the highly invasive whitefly species Bemisia tabaci Q/MED. Insect Mol. Biol. 2019, 28, 509–519. [Google Scholar] [CrossRef]
- Lin, J.; He, J.; Liang, A.; Wang, F. Transcriptome profiling and dimorphic expression of sex-related genes in fifth-instar nymphs of Sogatella furcifera, an important rice pest. Genomics 2020, 112, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zheng, W.; Qiao, J.; Li, Z.; Deng, Z.; Yuan, Y.; Zhang, H. Early embryonic transcriptomes of Zeugodacus tau provide insight into sex determination and differentiation genes. Insect Sci. 2021. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [Green Version]
- Davidson, N.M.; Oshlack, A. Corset: Enabling differential gene expression analysis for de novo assembled transcriptomes. Genome Biol. 2014, 15, 410. [Google Scholar] [CrossRef]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. Omi. J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franceschini, A.; Simonovic, M.; Roth, A.; Mering, C.V.; Szklarczyk, D.; Pletscherfrankild, S.; Jensen, L.J.; Kuhn, M.M.; Lin, J.; Minguez, P. STRING v9.1: Protein-protein interaction networks, with increased coverage and integration. Nucleic Acids Res. 2013, 41, 808–815. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Hrdlickova, R.; Toloue, M.; Tian, B. RNA-Seq methods for transcriptome analysis. Wiley Interdiscip. Rev. Rna. 2017, 8, e1364. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.C.; Brandman, O.; Petrov, D.A. Protein Evolution in the Context of Drosophila Development. J. Mol. Evol. 2005, 60, 774–785. [Google Scholar] [CrossRef]
- Connallon, T.; Knowles, L.L. Intergenomic conflict revealed by patterns of sex-biased gene expression. Trends Genet. 2005, 21, 495–499. [Google Scholar] [CrossRef]
- Degner, E.C.; Harrington, L.C. A mosquito sperm’s journey from male ejaculate to egg: Mechanisms, molecules, and methods for exploration. Mol. Reprod. Dev. 2016, 83, 897–911. [Google Scholar] [CrossRef]
- Ranz, J.M.; Castillo-Davis, C.I.; Meiklejohn, C.D.; Hartl, D.L. Sex-Dependent Gene Expression and Evolution of the Drosophila Transcriptome. Science 2003, 300, 1742–1745. [Google Scholar] [CrossRef]
- Hahn, M.W.; Lanzaro, G.C. Female-biased gene expression in the malaria mosquito Anopheles gambiae. Curr. Biol. 2005, 15, R192–R193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eads, B.D.; Colbourne, J.K.; Bohuski, E.; Andrews, J. Profiling sex-biased gene expression during parthenogenetic reproduction in Daphnia pulex. BMC Genom. 2007, 8, 464. [Google Scholar] [CrossRef] [Green Version]
- Meinertzhagen, I.A.; Hanson, T.E. The Development of Drosophila melanogaster. 1993. Available online: https://www.researchgate.net/publication/283413477_The_Development_of_Drosophila_melanogaster (accessed on 23 May 2022).
- Sæbøe-Larssen, S.; Lyamouri, M.; Merriam, J.; Oksvold, M.P.; Lambertsson, A. Ribosomal Protein Insufficiency and the Minute Syndrome in Drosophila: A Dose-Response Relationship. Genetics 1998, 148, 1215–1224. [Google Scholar] [CrossRef] [PubMed]
- Penalva, L.O.F.; Sánchez, L. RNA Binding Protein Sex-Lethal (Sxl) and Control of Drosophila Sex Determination and Dosage Compensation. Microbiol. Mol. Biol. Rev. 2003, 67, 343–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viger, R.; Mertineit, C.; Trasler, J.; Nemer, M. Transcription factor GATA-4 is expressed in a sexually dimorphic pattern during mouse gonadal development and is a potent activator of the Mullerian inhibiting substance promoter. Development 1998, 125, 2665–2675. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-F.; Bingham, N.; Parker, K.; Yao, H.H.-C. Sex-specific roles of beta-catenin in mouse gonadal development. Hum. Mol. Genet. 2009, 18, 405–417. [Google Scholar] [CrossRef] [Green Version]
- Schutt, C.; Nothiger, R. Structure, function and evolution of sex-determining systems in Dipteran insects. Development 2000, 127, 667–677. [Google Scholar] [CrossRef]
- Erickson, J.W.; Quintero, J.J. Indirect Effects of Ploidy Suggest X Chromosome Dose, not the X:A Ratio, Signals Sex in Drosophila. PLoS Biol. 2007, 5, e332. [Google Scholar] [CrossRef] [Green Version]
- Saccone, G.; Pane, A.; Polito, L. Sex determination in flies, fruitflies and butterflies. Genetica 2002, 116, 15–23. [Google Scholar] [CrossRef]
- Xu, J.; Zhan, S.; Chen, S.; Zeng, B.; Li, Z.; James, A.A.; Tan, A.; Huang, Y. Sexually dimorphic traits in the silkworm, Bombyx mori, are regulated by doublesex. Insect Biochem. Mol. Biol. 2017, 80, 42–51. [Google Scholar] [CrossRef] [Green Version]
- Bopp, D.; Saccone, G.; Beye, M. Sex Determination in Insects: Variations on a Common Theme. Sex. Dev. 2014, 8, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, Y.; Taniguchi, H.; Hamel, F.; Silversides, D.W.; Viger, R.S. A GATA4/WT1 cooperation regulates transcription of genes required for mammalian sex determination and differentiation. BMC Mol. Biol. 2008, 9, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osei-Amo, S.; Hussein, M.; Asad, S.; Hugo, L.; Asgari, S. Wolbachia -induced transcription factor GATA4 suppresses ovary-specific genes blastoderm-specific protein 25D and imaginal disc growth factor. Insect Mol. Biol. 2018, 27, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Vainio, S.; Heikkila, M.; Kispert, A.; Chin, N.; Andrew, P.M. Female development in mammals is regulated by Wnt-4 signalling. (cover story). Nature 1999, 397, 405. [Google Scholar] [CrossRef]
- Xu, J.; Yu, Y.; Chen, K.; Huang, Y. Intersex regulates female external genital and imaginal disc development in the silkworm. Insect Biochem. Mol. Biol. 2019, 108, 1–8. [Google Scholar] [CrossRef]
- Deshpande, G.; Nouri, A.; Schedl, P. Wnt Signaling in Sexual Dimorphism. Genetics 2015, 202, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Xu, W.-H. Wnt/β-catenin signaling regulates Helicoverpa armigera pupal development by up-regulating c-Myc and AP-4. Insect Biochem. Mol. Biol. 2014, 53, 44–53. [Google Scholar] [CrossRef]
- Defalco, T.J.; Geraldine, V.; Jenkins, A.B.; Michael, M.J.; Steven, R.; Mark, V.D. Sex-specific apoptosis regulates sexual dimorphism in the Drosophila embryonic gonad. Dev. Cell 2003, 5, 205–216. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.B.; Wu, H.; Yi, J.Q.; Li, J.H.; Cheng, Y.J.; Zhang, Y.F.; Sun, D.L.; An, Y.X. Transcriptome characterization and expression analysis related to sex-determining genes in Chilo sacchariphagus (Lepidoptera Crambidae), a key pest of sugarcane. Entomol. Res. 2022, 52, 77–79. [Google Scholar] [CrossRef]
- Jin, L.; Walker, A.S.; Fu, G.; Harvey-Samuel, T.; Dafa’Alla, T.; Miles, A.; Marubbi, T.; Granville, D.; Humphrey-Jones, N.; O’Connell, S.; et al. Engineered Female-Specific Lethality for Control of Pest Lepidoptera. ACS Synth. Biol. 2013, 2, 160–166. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Read Number | Raw Read Base Number | Clean Reads Base Number | Clean Read Q20 (%) | Clean Read Q30 (%) | Clean Read Ratio (%) |
|---|---|---|---|---|---|---|
| SBF1 | 43.82 Mb | 6.57 Gb | 6.48 Gb | 96.1 | 90.65 | 98.58 |
| SBF2 | 43.82 Mb | 6.57 Gb | 6.51 Gb | 95.98 | 90.41 | 99.01 |
| SBF3 | 41.43 Mb | 6.21 Gb | 6.1 Gb | 96.05 | 90.58 | 98.21 |
| SBM1 | 43.82 Mb | 6.58 Gb | 6.51 Gb | 96.16 | 90.78 | 98.98 |
| SBM2 | 43.82 Mb | 6.57 Gb | 6.49 Gb | 95.94 | 90.27 | 98.78 |
| SBM3 | 43.82 Mb | 6.57 Gb | 6.41 Gb | 96.29 | 91.08 | 97.57 |
| Category | Number | Ratio |
|---|---|---|
| Complete BUSCOs (C) | 1471 | 88.70% |
| Complete and single-copy BUSCOs (S) | 812 | 49% |
| Complete and duplicated BUSCOs (D) | 659 | 39.70% |
| Fragmented BUSCOs (F) | 22 | 1.30% |
| Missing BUSCOs (M) | 165 | 10.00% |
| Total BUSCO groups searched | 1658 | 100% |
| Database | Number | Percentage |
|---|---|---|
| NR | 32,735 | 54.17% |
| NT | 15,948 | 26.39% |
| Swiss-prot | 22,226 | 36.78% |
| KEGG | 24,875 | 41.16% |
| GOG | 21,590 | 35.73% |
| Pfam | 23,379 | 38.69% |
| GO | 15,013 | 24.84% |
| Total unigenes | 60,429 | 100% |
| Annotated unigenes | 34,847 | 57.67% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.-M.; He, W.-Z.; Wei, J.-L.; Chen, Z.-L.; Liao, F.; Qin, C.-X.; Pan, Y.-Q.; Shang, X.-K.; Lakshmanan, P.; Wang, M.; et al. Transcriptome Profiling Reveals Genes Related to Sex Determination and Differentiation in Sugarcane Borer (Chilo sacchariphagus Bojer). Insects 2022, 13, 500. https://doi.org/10.3390/insects13060500
Li A-M, He W-Z, Wei J-L, Chen Z-L, Liao F, Qin C-X, Pan Y-Q, Shang X-K, Lakshmanan P, Wang M, et al. Transcriptome Profiling Reveals Genes Related to Sex Determination and Differentiation in Sugarcane Borer (Chilo sacchariphagus Bojer). Insects. 2022; 13(6):500. https://doi.org/10.3390/insects13060500
Chicago/Turabian StyleLi, Ao-Mei, Wei-Zhong He, Ji-Li Wei, Zhong-Liang Chen, Fen Liao, Cui-Xian Qin, You-Qiang Pan, Xian-Kun Shang, Prakash Lakshmanan, Miao Wang, and et al. 2022. "Transcriptome Profiling Reveals Genes Related to Sex Determination and Differentiation in Sugarcane Borer (Chilo sacchariphagus Bojer)" Insects 13, no. 6: 500. https://doi.org/10.3390/insects13060500