
Population Fitness of Eupeodes corollae Fabricius (Diptera: Syrphidae) Feeding on Different Species of Aphids


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Aphid Collection and Culture
2.2. Hoverfly Collection and Culture
2.3. Life Table Study for E. corollae
2.4. Flight Tests for E. corollae
2.5. Data Analysis
3. Results
3.1. Development of E. corollae Fed on Different Aphids
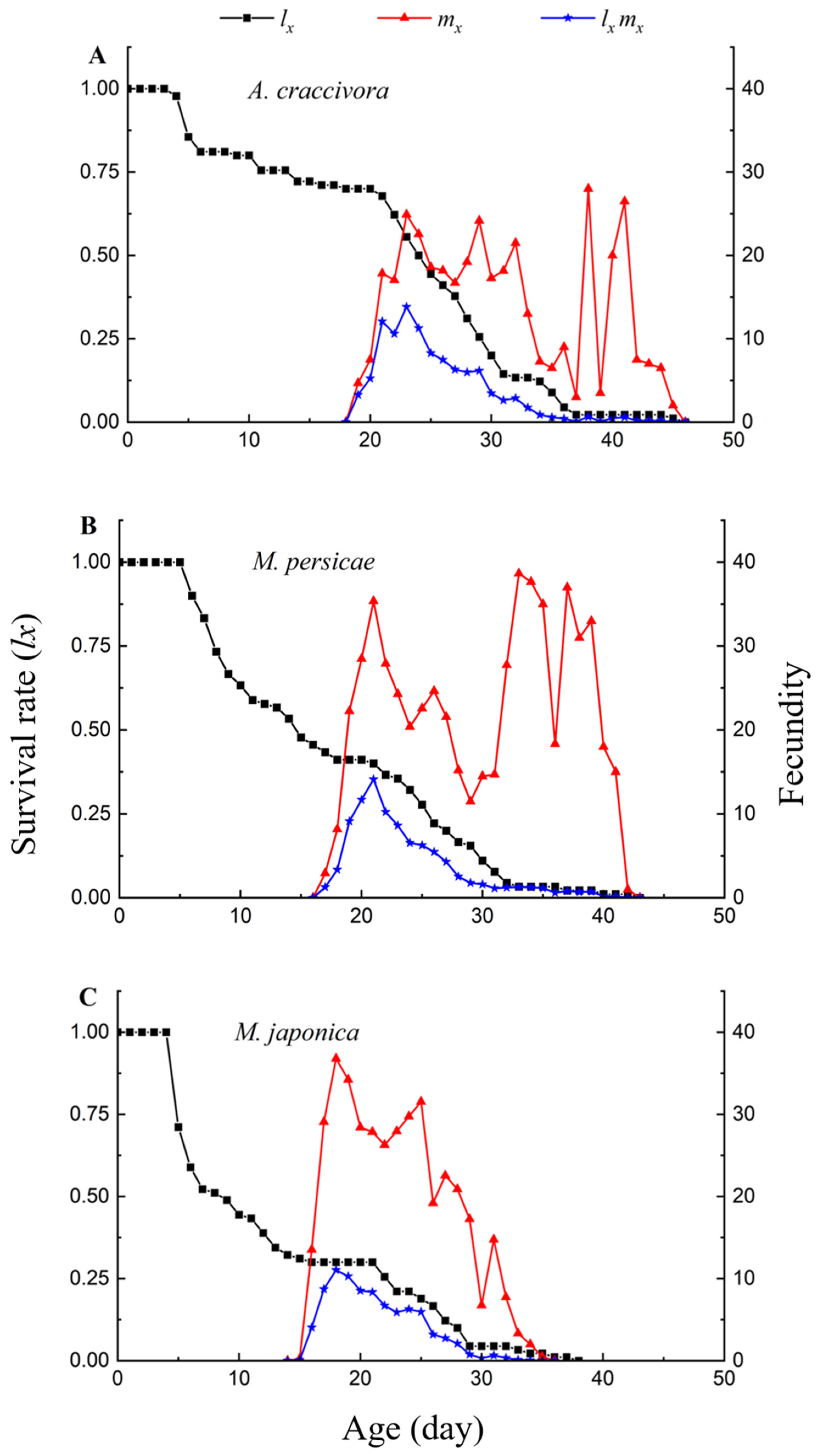
3.2. Reproduction of E. corollae Fed on Different Aphids
3.3. Flight Performance of E. corollae Fed on Different Aphids
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mengual, X.; Ståhls, G.; Rojo, S. Molecular phylogeny of Allograpta (Diptera, Syrphidae) reveals diversity of lineages and non-monophyly of phytophagous taxa. Mol. Phylogenet. Evol. 2008, 49, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Mengual, X.; Ståhls, G.; Rojo, S. Phylogenetic relationships and taxonomic ranking of pipizine flower flies (Diptera: Syrphidae) with implications for the evolution of aphidophagy. Cladistics 2015, 31, 491–508. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.Y. Hoverfly. Biol. Bull. 2004, 2, 9–12. [Google Scholar]
- Tian, J.; Ren, B.Z. Overview of hoverfly research. J. Jilin Agric. Univ. 2019, 41, 1–10. [Google Scholar]
- Ho, K.K.; Zheng, Z.M.; Zhang, H.J. Research progress of Syrphidae in China. J. Hanzhong Univ. 2002, 3, 70–75. [Google Scholar]
- Fischer, O.A.; Mátlová, L.; Dvorská, L.; Švástová, P.; Bartoš, M.; Weston, R.T.; Pavlík, I. Various stages in the life cycle of syrphid flies (Eristalis tenax; Diptera: Syrphidae) as potential mechanical vectors of pathogens causing mycobacterial infections in pig herds. Folia Microbiol. 2006, 51, 147–153. [Google Scholar] [CrossRef]
- Ymilie, B.; Marc, F.; Eric, L. Biological control of the foxglove aphid using a banker plant with Eupeodes americanus (Diptera: Syrphidae) in experimental and commercial greenhouses. Biol. Control 2021, 155, 104541. [Google Scholar]
- Reemer, M. Review and phylogenetic evaluation of associations between Microdontinae (Diptera: Syrphidae) and Ants (Hymenoptera: Formicidae). J. Entomol. 2013, 1, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Wotton, K.R.; Gao, B.; Myles, H.M.; Morris, R.K.A.; Ball, S.G.B.; Lim, K.S.; Reynolds, D.R.; Hu, G.; Chapman, J.W. Mass seasonal migrations of hoverflies provide extensive pollination and crop protection services. Curr. Biol. 2019, 29, 2167–2173. [Google Scholar] [CrossRef]
- Li, X.Y.; Luo, Y.Z. Study on the predation effect of Syrphidae on three species of aphids. J. Agric. Yunnan Univ. 2001, 2, 102–104. [Google Scholar]
- Katherine, C.R.B.; Mark, A.G.; Damien, M.H.; Kunin, W.E.; Mitschuna, N.; Morse, H.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; et al. A systems approach reveals urban pollinator hotspots and conservation opportunities. Nat. Ecol. Evol. 2019, 3, 363–373. [Google Scholar]
- Rader, R.; Cunningham, S.; Howlett, B.; Inouye, D. Non-Bee insects as visitors and pollinators of crops: Biology, ecology, and management. Annu. Rev. Entomol. 2020, 65, 391–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Cheng, J.; Luo, D.; Luo, Y.B. Using pollination syndrome prediction: Simulated breeding sites of Paphiopedilum dianthum to deceive pollination by female hoverflies. J. Syst. Evol. 2007, 4, 551–560. [Google Scholar] [CrossRef]
- He, J.L.; Chu, X.P.; Sun, X.Q.; Ye, W.J. Feeding and utilization evaluation of hoverfly. J. Agric. Shanghai Univ. 1994, 2, 79–83. [Google Scholar]
- Sagar, P. Role of insects in cross pollination of fennel crop at Ludhiana [India]. J. Res. Punjab Agric. Univ. 1981, 18, 388–392. [Google Scholar]
- Burgett, M. Pollination of parsley (Petroselinum crispum) grown for seed. J. Apic. Res. 1980, 19, 79–82. [Google Scholar] [CrossRef]
- Pu, D.-Q.; Liu, H.-L.; Gong, Y.Y.; Ji, P.-C.; Li, Y.-J.; Mou, F.-S.; Wei, S.-J. Mitochondrial genomes of the hoverflies Episyrphus balteatus and Eupeodes corollae (Diptera: Syrphidae), with a phylogenetic analysis of muscomorpha. Sci. Rep. 2017, 7, 44300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, J.L. The biology and ecology of the predatory hoverfly. J. Agric. Shanghai Univ. 1989, 4, 325–331. [Google Scholar]
- Li, H.; Jiang, S.S.; Zhang, H.W.; Geng, T.; Wyckhuys, K.A.G.; Wu, K.M. Two-way predation between immature stages of the hoverfly Eupeodes corollae and the invasive fall armyworm (Spodoptera frugiperda J. E. Smith). J. Integr. Agric. 2021, 20, 829–839. [Google Scholar] [CrossRef]
- Yang, F.C.; Li, Z.W.; Gao, H.C.; Hui, F.H.; Lin, G.L. Study on the biology of Syrphidae and its control on wheat aphid. J. Environ. Entomol. 1989, 3, 116–121. [Google Scholar]
- Xiong, H.Z.; Dong, H.F. Experiment on feeding and releasing larvae of Syrphids larvae in greenhouse. J. Biol. Control Chin. 1992, 1, 6–9. [Google Scholar]
- Rob, M.; Sten, B.; Wäckers, F.L.; Pekas, A. Aphid ophagous hoverflies reduce foxglove aphid infestations and improve seed set and fruit yield in sweet pepper. Pest Manag. Sci. 2021, 77, 2690–2696. [Google Scholar]
- Iwai, H.; Niijima, K.; Matsuka, M. An artificial diet for aphidophagous syrphids, Episyrphus balteatus (de Geer) and Eupeodes bucculatus (Rondani) (Diptera: Syrphidae) using drone honeybee brood powder. Appl. Entomol. Zool. 2007, 42, 167–172. [Google Scholar] [CrossRef] [Green Version]
- Hideki, I.; Keiko, N.; Mitsuo, M. Improvement of artificial diet for aphidophagous syrphids, Episyrphus balteatus (de Geer) and Eupeodes bucculatus (Rondani) (Diptera: Syrphidae)—additional effects of fatty acids and preservatives. Appl. Entomol. Zool. 2009, 44, 439–446. [Google Scholar]
- Pu, Z.G.; Chi, L. Research progress on biological characteristics and artificial indoor rearing technology of Syrphidae. Heilongjiang Agric. Sci. 2006, 2006, 120–123. [Google Scholar]
- Kan, E. Assessment of aphid colonies by hoverflies. I maple aphids and Episyrphus balteatus (de Geer) (Diptera: Syrphidae). J. Ethol. 1988, 6, 39–48. [Google Scholar] [CrossRef]
- Sadeghi, H.; Gilbert, F. Aphid suitability and its relationship to oviposition preference in predatory hoverflies. J. Anim. Ecol. 2000, 69, 771–784. [Google Scholar] [CrossRef] [Green Version]
- Barlow, C.A. On the biology and reproductive capacity of Syrphus corollae Fab. (Syrphidae) in the laboratory. Entomol. Exp. Appl. 1961, 4, 91–100. [Google Scholar] [CrossRef]
- Obopile, M.; Ositile, B. Life table and population parameters of cowpea aphid, Aphis craccivora Koch (Homoptera: Aphididae) on five cowpea Vigna unguiculata (L. Walp.) varieties. J. Pest Sci. 2010, 83, 9–14. [Google Scholar] [CrossRef]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Ge, S.S.; He, L.M.; He, W.; Xu, R.B.; Sun, X.T.; Wu, K.M. Determination of flying ability of Spodoptera frugiperda. Plant Prot. 2019, 45, 28–33. [Google Scholar]
- He, L.M.; Jiang, S.; Chen, Y.C.; Wyckhuy, K.A.G.; Ge, S.S.; He, W.; Gao, X.-W.; Wu, K.-M. Adult nutrition affects reproduction and flight performance of the invasive fall armyworm, Spodoptera frugiperda in China. J. Integr. Agric. 2021, 20, 715–726. [Google Scholar] [CrossRef]
- Luo, Q.H. A Preliminary observation on the habits of hoverfly. Agric. Sci. Guizhou 1982, 5, 21–23. [Google Scholar]
- Lan, X.; Luo, J.; Cheng, X.Y. Analysis of predatory hoverfly species and their control effects on soybean aphid in the soybean ecoregion of northeast China. J. Appl. Entomol. 2011, 48, 1625–1630. [Google Scholar]
- He, J.L.; Sun, X.Q.; Gui, L.M.; Ye, W.J. A preliminary study on the biology of hoverfly in Shanghai Area. J. Agric. Shanghai Univ. 1990, 03, 221–228. [Google Scholar]
- Lee, J.H.; Kang, T.J. Functional response of Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae) to Aphis gossypii Glover (Homoptera: Aphididae) in the Laboratory. Biol. Control 2004, 31, 306–310. [Google Scholar] [CrossRef]
- Moreau, J.; Benrey, B.; Thiéry, D. Grape variety affects larval performance and also female reproductive performance of the European grapevine moth Lobesia botrana (Lepidoptera: Tortricidae). Bull. Entomol. Res. 2006, 96, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, H. Effect of prey density on behaviour and development of the predatory mosquito, Toxorhynchites towadensis. Entomol. Exp. Appl. 1995, 76, 97–103. [Google Scholar] [CrossRef]
- Liu, J.B.; Cui, Y.H.; Gao, J.F.; Wang, W.H.; Cheng, X.H.; Liu, Z.Y. Observation of biological characteristics of Eupeodes corollae. Jilin Agric. Sci. 2005, 1, 38–39. [Google Scholar]
- Eduardo, M.S.; Padró, J.; Milla, C.P.; Tureo, D.T.; Carreira, V.P.; Soto, I.M. Pupal emergence pattern in cactophilic Drosophila and the effect of host plants. Insect Sci. 2017, 25, 1108–1118. [Google Scholar]
- Li, Y.; Leng, C.M.; Wu, J.X.; Li, Y.P. Effects of different host plants on growth, development and reproduction of Plutella xylostella. Acta Agric. Boreali Occident. Sin. 2019, 28, 475–480. [Google Scholar]
- Jabraeil, R.; Bahram, N.; Seyed, A.H. Comparative performance of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) on various host plants. J. Pest Sci. 2014, 87, 29–37. [Google Scholar]
- Putra, N.S.; Yasuda, H. Effects of prey species and its density on larval performance of two species of hoverfly larvae, Episyrphus balteatus de Geer and Eupeodes corollae Fabricius (Diptera: Syrphidae). Appl. Entomol. Zool. 2006, 41, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Sugiura, K.; Takada, H. Suitability of seven aphid species as prey of Cheilomenes sexmaculata (Fabricius) (Coleoptera: Coccinellidae). Appl. Entomol. Zool. 1998, 42, 7–14. [Google Scholar] [CrossRef] [Green Version]
- Okamoto, H. Laboratory studies on food ecology of aphidophagous lady beetles (Coleoptera: Coccinellidae). Mem. Fac. Eng. Kyushu Univ. 1978, 32, 1–94. [Google Scholar]
- HE, L.M.; WU, Q.L.; GAO, X.W.; Wu, K.-M. Population life tables for the invasive fall armyworm, Spodoptera frugiperda fed on major oil crops planted in China. J. Integr. Agric. 2021, 20, 745–754. [Google Scholar] [CrossRef]
- Dong, K.; Dong, Y.; Li, X.Y.; Luo, Y.Z. Effects of feeding with different aphids on the growth and development of Syrphidae. J. Environ. Entomol. 2003, 4, 165–168. [Google Scholar]
- Yin, Y.Y.; Lin, Q.C.; Zhai, Y.F.; Zheng, L.; Yu, Y.; Chen, H. The flight ability and spreading ability of Encarsia formosa Gahan. J. Environ. Entomol. 2019, 41, 1355–1360. [Google Scholar]
- Guo, J.L.; Fu, X.W.; Zhao, X.C.; Wu, K.M. Study on the flying ability of Agrotis segetum. J. Environ. Entomol. 2016, 38, 888–895. [Google Scholar]
- Wu, K.M.; Guo, Y.Y. Flying activity of Helicoverpa armigera. J. Ecol. Chin. 1996, 6, 612–617. [Google Scholar]
- Nielsen, B.O. On a migration of hoverflies (Dipt., Syrphidae) and sawflies (Hym., Tenthrdinidae) observed in Denmark, August 1967. Acta Entomol. Sin. 1968, 36, 215–224. [Google Scholar]
- Lucas, É.; Coderre, D.; Brodeur, J. Intraguild predation among aphid predators: Characterization and influence of extraguild prey density. J. Ecol. 1998, 79, 1084–1092. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Stage | Sex | Stage Duration (d) | |||||
|---|---|---|---|---|---|---|---|
| A. craccivora | n | M. persicae | n | M. japonica | n | ||
| Egg | ♀ + ♂ | 2.0 ± 0.00 a | 90 | 2.0 ± 0.00 a | 90 | 2.0 ± 0.00 a | 90 |
| ♀ | 2.0 ± 0.00 A | 31 | 2.0 ± 0.00 A | 20 | 2.0 ± 0.00 A | 13 | |
| ♂ | 2.0 ± 0.00 A | 33 | 2.0 ± 0.00 A | 21 | 2.0 ± 0.00 A | 14 | |
| 1st instar | ♀ + ♂ | 2.0 ± 0.06 a | 75 | 1.4 ± 0.06 b | 90 | 1.8 ± 0.09 a | 55 |
| ♀ | 2.1 ± 0.06 A | 31 | 1.5 ± 0.18 A | 20 | 1.5 ± 0.18 B | 13 | |
| ♂ | 1.8 ± 0.09 B | 33 | 1.0 ± 0.00 B | 21 | 2.2 ± 0.11 A | 14 | |
| 2nd instar | ♀ + ♂ | 2.1 ± 0.03 a | 72 | 1.9 ± 0.06 b | 76 | 1.8 ± 0.07 b | 49 |
| ♀ | 2.1 ± 0.04 A | 31 | 1.7 ± 0.14 A | 20 | 1.8 ± 0.15 A | 13 | |
| ♂ | 2.1 ± 0.04 A | 33 | 1.8 ± 0.09 A | 21 | 1.6 ± 0.13 A | 14 | |
| 3rd instar | ♀ + ♂ | 3.5 ± 0.07 a | 65 | 3.6 ± 0.13 a | 55 | 3.0 ± 0.13 b | 41 |
| ♀ | 3.6 ± 0.09 A | 31 | 3.1 ± 0.08 A | 20 | 2.8 ± 0.10 A | 13 | |
| ♂ | 3.5 ± 0.10 A | 33 | 3.4 ± 0.13 A | 21 | 3.0 ± 0.23 A | 14 | |
| Larval stage (1st–3rd instar) | ♀ + ♂ | 7.6 ± 0.09 a | 65 | 6.7 ± 0.16 b | 55 | 6.7 ± 0.16 b | 41 |
| ♀ | 7.8 ± 0.10 A | 31 | 6.4 ± 0.11 A | 20 | 6.2 ± 0.20 A | 13 | |
| ♂ | 7.4 ± 0.14 B | 33 | 6.1 ± 0.16 A | 21 | 6.8 ± 0.23 A | 14 | |
| Pupae | ♀ + ♂ | 6.9 ± 0.04 a | 64 | 6.3 ± 0.07 b | 41 | 6.1 ± 0.07 b | 27 |
| ♀ | 7.0 ± 0.00 A | 31 | 6.7 ± 0.10 A | 20 | 6.1 ± 0.10 A | 13 | |
| ♂ | 6.8 ± 0.07 B | 33 | 6.0 ± 0.00 B | 21 | 6.1 ± 0.10 A | 14 | |
| Adult | ♀ + ♂ | 11.8 ± 0.69 a | 64 | 12.5 ± 0.84 a | 41 | 12.4 ± 0.77 a | 27 |
| ♀ | 14.5 ± 1.11 A | 31 | 14.9 ± 1.25 A | 20 | 13.3 ± 0.91 A | 13 | |
| ♂ | 9.3 ± 0.55 B | 33 | 10.1 ± 0.89 B | 21 | 11.6 ± 1.21 A | 14 | |
| Egg–adult | ♀ + ♂ | 28.3 ± 0.70 a | 64 | 27.1 ± 0.88 a | 41 | 27.1 ± 0.83 a | 27 |
| ♀ | 31.3 ± 1.07 A | 31 | 30.0 ± 1.26 A | 20 | 27.7 ± 1.08 A | 13 | |
| ♂ | 25.5 ± 0.59 B | 33 | 24.3 ± 0.86 B | 21 | 26.6 ± 1.26 A | 14 | |
| Variable | A. craccivora | n | M. persicae | n | M. japonica | n |
|---|---|---|---|---|---|---|
| Larval survival rate (%) | 80.0 ± 5.77 a | 3 | 84.4 ± 2.94 a | 3 | 54.4 ± 4.84 b | 3 |
| Pupation rate (%) | 89.9 ± 5.52 a | 3 | 72.8 ± 6.78 a | 3 | 83.1 ± 7.62 a | 3 |
| Emergence rate (%) | 98.0 ± 1.96 a | 3 | 73.9 ± 6.29 b | 3 | 64.7 ± 4.05 b | 3 |
| Hatching rate of egg (%) | 90.0 ± 5.09 a | 3 | 88.89 ± 4.01 a | 3 | 82.2 ± 6.76 a | 3 |
| Mass (g) | ||||||
| ♀ | 0.0564 ± 0.0133 A | 31 | 0.0317 ± 0.0009 A | 20 | 0.0315 ± 0.0006 A | 13 |
| ♂ | 0.0332 ± 0.0005 A | 33 | 0.0307 ± 0.0009 A | 21 | 0.0309 ± 0.0009 A | 14 |
| ♀ + ♂ | 0.0444 ± 0.0066 a | 64 | 0.0312 ± 0.0006 b | 41 | 0.0312 ± 0.0005 b | 27 |
| Prey | A. craccivora | M. persicae | M. japonica |
|---|---|---|---|
| T | 24.85 ± 0.34 a | 23.12 ± 0.41 b | 21.05 ± 0.33 c |
| λ | 1.2052 ± 0.0089 a | 1.2167 ± 0.0137 a | 1.2335 ± 0.0182 a |
| r | 0.1866 ± 0.0074 a | 0.1961 ± 0.0113 a | 0.2098 ± 0.0149 a |
| R0 | 105.38 ± 19.65 a | 96.39 ± 24.90 a | 86.16 ± 24.42 a |
| Prey | A. craccivora | n | M. persicae | n | M. japonica | n |
|---|---|---|---|---|---|---|
| No. of ovipositions | 305.9 ± 36.23 b | 31 | 433.7 ± 73.80 ab | 20 | 596.5 ± 74.85 a | 13 |
| Oviposition duration (d) | 10.2 ± 1.11 a | 30 | 11.6 ± 1.18 a | 19 | 11.0 ± 0.89 a | 13 |
| Total pre- oviposition (d) | 20.3 ± 0.18 a | 30 | 18.9 ± 0.29 b | 19 | 16.6 ± 0.37 c | 13 |
| Pre-oviposition (d) | 3.5 ± 0.11 a | 30 | 3.8 ± 0.26 a | 19 | 2.2 ± 0.17 b | 13 |
| Prey | A. craccivora (n = 32) | M. persicae (n = 36) | M. japonica (n = 37) | |
|---|---|---|---|---|
| Duration (s) | Max | 6217.2 | 4305.6 | 5079.6 |
| Mean ± SE | 2442.3 ± 240.3 a | 2037.4 ± 151.3 a | 2521.9 ± 186.8 a | |
| Distance (m) | Max | 1622.7 | 950.2 | 1407.3 |
| Mean ± SE | 582.4 ± 69.2 a | 448.2 ± 35.4 a | 624.0 ± 52.0 a | |
| Velocity (m/s) | Max | 0.38 | 0.33 | 0.36 |
| Mean ± SE | 0.23 ± 0.01 a | 0.22 ± 0.01 a | 0.25 ± 0.01 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, S.; Li, H.; He, L.; Wu, K. Population Fitness of Eupeodes corollae Fabricius (Diptera: Syrphidae) Feeding on Different Species of Aphids. Insects 2022, 13, 494. https://doi.org/10.3390/insects13060494
Jiang S, Li H, He L, Wu K. Population Fitness of Eupeodes corollae Fabricius (Diptera: Syrphidae) Feeding on Different Species of Aphids. Insects. 2022; 13(6):494. https://doi.org/10.3390/insects13060494
Chicago/Turabian StyleJiang, Shanshan, Hui Li, Limei He, and Kongming Wu. 2022. "Population Fitness of Eupeodes corollae Fabricius (Diptera: Syrphidae) Feeding on Different Species of Aphids" Insects 13, no. 6: 494. https://doi.org/10.3390/insects13060494