
Safety Evaluation of Chemical Insecticides to Tetrastichus howardi (Hymenoptera: Eulophidae), a Pupal Parasitoid of Spodoptera frugiperda (Lepidoptera: Noctuidae) Using Three Exposure Routes

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Insecticides and Chemicals
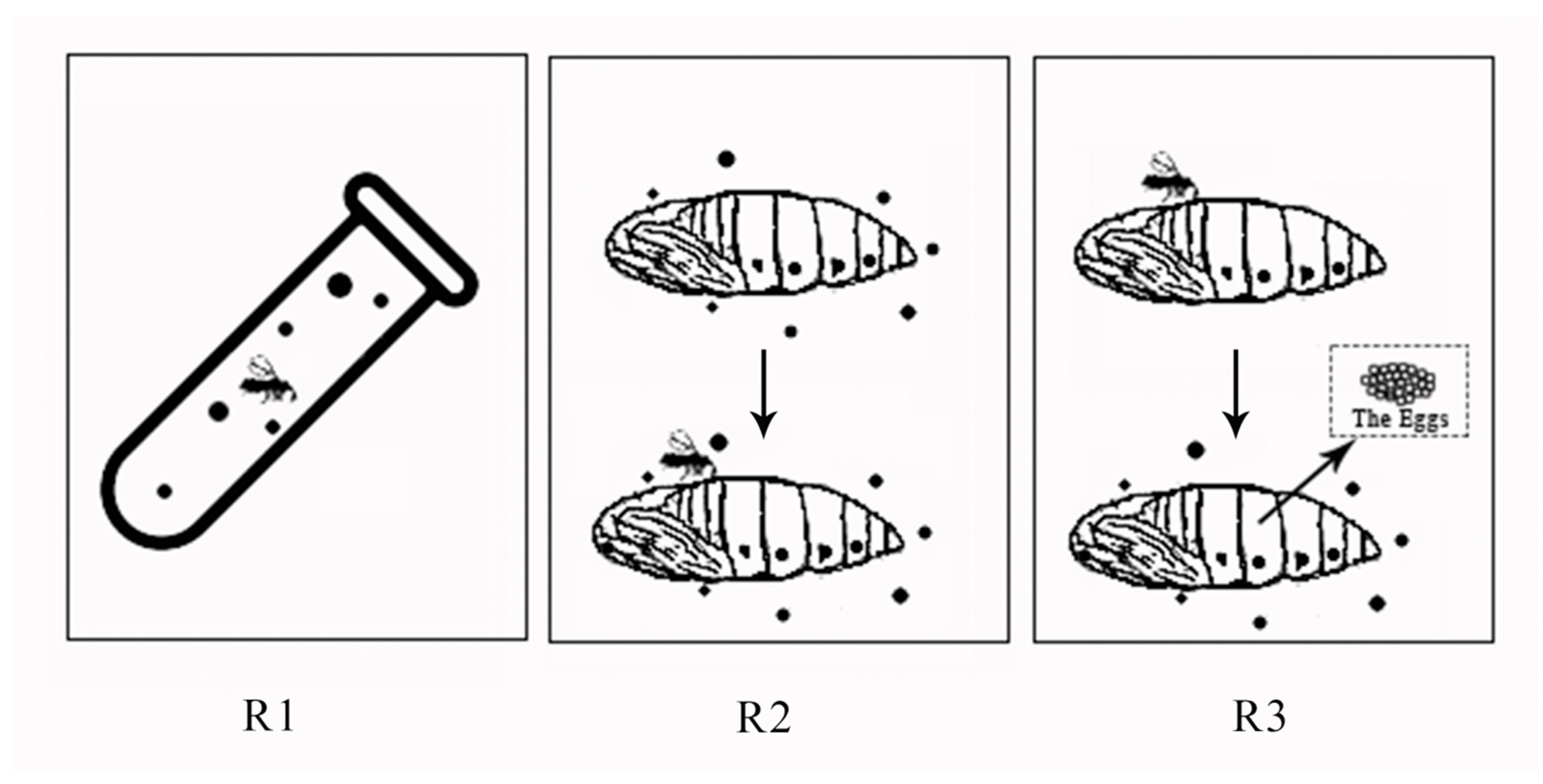
2.3. Risk Assessment of Six Insecticides against T. howardi
2.4. Sublethal Effects of Indoxacarb on T. howardi
2.5. Data Analysis
3. Results
3.1. Risk Assessment of Six Insecticides against T. howardi
3.2. Sublethal Effects of Indoxacarb on T. howardi Development and Fecundity
3.2.1. Percentage of Parasitism and Emergence
3.2.2. Offspring Female Ratio
3.2.3. Number of Emerged Offspring
3.2.4. Developmental Duration
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stenberg, J.A. A Conceptual Framework for Integrated Pest Management. Trends. Plant Sci. 2017, 22, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Longley, M.; Jepson, P.C. Effects of honeydew and insecticide residues on the distribution of foraging aphid parasitoids under glasshouse and field conditions. Entomol. Exp. Appl. 1996, 81, 189–198. [Google Scholar] [CrossRef]
- Alcántara-de la Cruz, R.; Zanuncio, J.C.; Lacerda, M.C.; Wilcken, C.F.; Fernandes, F.L.; Tavares, W.D.; Soares, M.A.; Sediyama, C.S. Side-effects of pesticides on the generalist endoparasitoid Palmistichus elaeisis (Hymenoptera: Eulophidae). Sci. Rep. 2017, 7, 557–561. [Google Scholar]
- Khan, R.R.; Al-Ghafri, T.H.A.; Al-Khatri, S.A.H.; Al-Mazidi, I.S.S.; Al-Rawahi, F.G. Resistance to deltamethrin and fenitrothion in dubas bug, Ommatissus lybicus de Bergevin (Homoptera: Tropiduchidae) and possible biochemical mechanisms. Science 2020, 10, 13220. [Google Scholar] [CrossRef]
- Biondi, A.; Campolo, O.; Desneux, N.; Siscaro, G.; Palmeri, V.; Zappalà, L. Life stage-dependent susceptibility of Aphytis melinus DeBach (Hymenoptera: Aphelinidae) to two pesticides commonly used in citrus orchards. Chemosphere 2015, 128, 142–147. [Google Scholar] [CrossRef]
- Desneux, N.; Fauvergue, X.; Dechaume-Moncharmont, F.X.; Kerhoas, L.; Ballanger, Y.; Kaiser, L. Diaeretiella rapae limits myzus persicae populations after applications of deltamethrin in oilseed rape. J. Econ. Entomol. 2005, 98, 9–17. [Google Scholar] [CrossRef]
- Desneux, N.; Decourtye, A.; Delpuech, J.M. The Sublethal Effects of Pesticides on Beneficial Arthropods. Annu. Rev. Entomol. 2007, 52, 81–106. [Google Scholar] [CrossRef]
- Biondi, A.; Mommaerts, V.; Smagghe, G.; Viñuela, E.; Zappalà, L.; Desneux, N. The non-target impact of spinosyns on beneficial arthropods. Pest. Manag. Sci. 2012, 68, 1523–1536. [Google Scholar] [CrossRef]
- Desneux, N.; Denoyelle, R.; Kaiser, L. A multi-step bioassay to assess the effect of the deltamethrin on the parasitic wasp Aphidius ervi. Chemosphere 2006, 65, 1697–1706. [Google Scholar] [CrossRef]
- Rodrigues, A.; Pereira, F.F.; Barbosa, P.R.R.; Silva-Torres, C.S.A.; Torres, J.B. Parasitism Behavior of Tetrastichus howardi (Hymenoptera: Eulophidae) on Larvae and Pupae of Sugarcane Borers. J. Insect Behav. 2021, 34, 71–81. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Q.L.; Zhang, H.W.; Wu, K.M. Spread of invasive migratory pest Spodoptera frugiperda and management practices throughout China. J. Integr. Agric. 2021, 20, 637–645. [Google Scholar] [CrossRef]
- Lu, H.; Tang, J.H.; Lyu, B.Q.; Ma, Z.L.; He, X.; Chen, Q.; Su, H. Recent advances in biological control and invasion risk of Spodoptera frugiperda. Chin. J. Trop. Crops. 2019, 40, 1237–1244. [Google Scholar]
- Li, X.J.; Wu, M.F.; Ma, J.; Gao, B.Y.; Wu, Q.L.; Chen, A.D.; Liu, J.; Jiang, Y.Y.; Zhai, B.P.; Early, R.; et al. Prediction of migratory routes of the invasive fall armyworm in eastern China using a trajectory analytical approach. Pest. Manag. Sci. 2020, 76, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.F.; Qi, G.J.; Chen, H.; Ma, J.; Liu, J.; Jiang, Y.Y.; Lee, G.S.; Otuka, A.; Hu, G. Overseas immigration of fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) invading Korea and Japan in 2019. Insect. Sci. 2022, 29, 505–520. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Lu, B.Q.; Lu, H.; Tang, J.H.; Liu, Y.L.; Yan, S.Q. Analysis of cannibalism predation among larvae of Spodoptera frugiperda. J. Environ. Entomol. 2021. Available online: http://kns.cnki.net/kcms/detail/44.1640.q.20210812.0935.002.html (accessed on 21 November 2021).
- Tang, J.H.; Lyu, B.Q.; Lu, H.; Ji, X.C.; Yang, P.Y.; Su, H.; Cai, B. Investigation and preliminary study of biological characteristic of parasitic wasp of Spodoptera frugiperda in Hainan. Chin. J. Trop. Crops. 2020, 41, 1189–1195. [Google Scholar]
- Yan, S.Q.; Su, H.; Lyu, B.Q.; Lu, H.; Tang, J.H.; Zhuo, Z.H.; Yang, F. Effects of temperature and supplementary food on the development and fecundity of the Eulophid parasitoid Tetrastichus howardi on Spodoptera frugiperda (Lepidoptera: Noctuidae). Int. J. Pest. Manag. 2021. Available online: https://doi.org/10.1080/09670874.2021.1968535 (accessed on 27 January 2022). [CrossRef]
- Fontes, J.; Roja, I.S.; Tavares, J.; Oliveira, L. Lethal and Sublethal Effects of Various Pesticides on Trichogramma achaeae (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2018, 111, 1219–1226. [Google Scholar] [CrossRef]
- Jiang, J.G.; Liu, X.; Zhang, Z.Q.; Liu, F.; Mu, W. Lethal and sublethal impact of sulfoxaflor on three species of Trichogramma parasitoid wasps (Hymenoptera: Trichogrammatidae). Biol. Control 2019, 134, 32–37. [Google Scholar] [CrossRef]
- Wang, Q.Q.; Cui, L.; Wang, L.; Yang, D.B.; Yan, X.J.; Yuan, H.Z.; Dong, F.S.; Zheng, Y.Q.; Rui, C.H. Prevention and Control of Fall Armyworm, Spodoptera frugiperda: Experience in Foreign Countries and Suggestion in China. Mod. Agrochem. 2020, 19, 1–6. [Google Scholar]
- Jatsch, A.S.; Ruther, J. Acetone application for administration of bioactive substances has no negative effects on longevity, fitness, and sexual communication in a parasitic wasp. PLoS ONE 2021, 16, e0245698. [Google Scholar] [CrossRef]
- Preetha, G.; Stanley, J.; Suresh, S.; Samiyappan, R. Risk assessment of insecticides used in rice on miridbug, Cyrtorhinus lividipennis Reuter, the important predator of brown planthopper, Nilaparvata lugens (Stal.). Chemosphere 2010, 80, 498–503. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Li, X.Y.; Zhou, C.; Liu, F.; Mu, W. Toxicity of nine insecticides on four natural enemies of Spodoptera exigua. Sci. Rep. 2016, 6, 39060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desneux, N.; Pham-Delègue, M.H.; Kaiser, L. Effects of sub-lethal and lethal doses of lambda-cyhalothrin on oviposition experience and host-searching behaviour of a parasitic wasp, Aphidius ervi. Pest. Manag. Sci. 2004, 60, 381–389. [Google Scholar] [CrossRef]
- Ham, E.H.; Lee, J.S.; Jang, M.Y.; Park, J.H. Toxic effects of 12 pesticides on green lacewing, Chrysoperla nipponensis (Okamoto) (Neuroptera: Chrysopidae). Entomol. Res. 2019, 49, 305–312. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Tang, Q.Y.; Zhang, C.X. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef]
- Nozad-Bonab, Z.; Hejazi, M.J.; Iranipour, S.; Arzanlou, M.; Biondi, A. Lethal and sublethal effects of synthetic and bio-insecticides on Trichogramma brassicae parasitizing Tuta absoluta. PLoS ONE 2021, 16, e0243334. [Google Scholar]
- Chen, X.F.; Song, M.; Qi, S.Z.; Wang, C.J. Safety Evaluation of Eleven Insecticides to Trichogramma nubilale (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2013, 106, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Wing, K.D.; Sacher, M.; Kagaya, Y.; Tsurubuchi, Y.; Mulderig, L.; Connair, M.; Schnee, M. Bioactivation and mode of action of the oxadiazine indoxacarb in insects. Crop. Prot. 2000, 19, 537–545. [Google Scholar] [CrossRef]
- Kaiser, I.S.; Kanno, R.H.; Bolzan, A.; Amaral, F.S.A.; Lira, E.C.; Guidolin, A.S.; Omoto, C. Baseline Response, Monitoring, and Cross-Resistance of Spodoptera frugiperda (Lepidoptera: Noctuidae) to Sodium Channel Blocker Insecticides in Brazil. J. Econ. Entomol. 2021, 114, 903–913. [Google Scholar] [CrossRef]
- Souza, J.R.; Carvalho, G.A.; Moura, A.P.; Couto, M.H.G.; Maia, J.B. Impact of insecticides used to control Spodoptera frugiperda (J.E. Smith) in corn on survival, sex ratio, and reproduction of Trichogramma pretiosum Riley offspring. Chil. J. Agric. Res. 2013, 73, 122–127. [Google Scholar] [CrossRef] [Green Version]
- Costa, E.S.P.; Soares, M.A.; Caldeira, Z.V.; Veloso, R.V.; da Silva, L.A.; da Silva, D.J.H.; Santos, I.C.D.; Castro, B.M.D.E.; Zanuncio, J.C.; Legaspi, J.C. Selectivity of deltamethrin doses on Palmistichus elaeisis (Hymenoptera: Eulophidae) parasitizing Tenebrio molitor (Coleoptera: Tenebrionidae). Sci. Rep. 2020, 10, 72–87. [Google Scholar]
- Parsaeyan, E.; Safavi, S.A.; Saber, M.; Poorjavad, N. Effects of emamectin benzoate and cypermethrin on the demography of Trichogramma brassicae Bezdenko. Crop. Prot. 2018, 110, 269–274. [Google Scholar] [CrossRef]
- Karatolos, N.; Hatcher, P.E. The effect of acetylsalicylic acid and oxalic acid on Myzus persicae and Aphidius colemani. Entomol. Exp. Appl. 2009, 130, 98–105. [Google Scholar] [CrossRef]
- Zou, Y.; Geuverink, E.; Beukeboom, L.W.; Verhulst, E.C.; van de Zande, L. A chimeric gene paternally instructs female sex determination in the haplodiploid wasp Nasonia. Science 2020, 370, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Broadley, H.J.; Gould, J.R.; Sullivan, L.T.; Wang, X.Y.; Hoelmer, K.A.; Hickin, M.L.; Elkinton, J.S. Life History and Rearing of Anastatus orientalis (Hymenoptera: Eupelmidae), an Egg Parasitoid of the Spotted Lanternfly (Hemiptera: Fulgoridae). Environ. Entomol. 2021, 50, 28–35. [Google Scholar] [CrossRef]
- Ramos, R.S.; de Araújo, V.C.R.; Pereira, R.R.; Martins, J.C.; Queiroz, O.S.; Silva, R.S.; Picanço, M.C. Investigation of the lethal and behavioral effects of commercial insecticides on the parasitoid wasp Copidosoma truncatellum. Chemosphere 2018, 191, 770–778. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Pesticide | LC50 (mg a.i L−1) | Regression Equation | χ2 Value | p a | 95% Confidence Limits | Recommended Dose (g a.i ha−1) | Risk Quotient (RQ) | Category |
|---|---|---|---|---|---|---|---|---|
| Emamectin benzoate | 0.09 | y = 2.140 + 2.065x | 11.08 | 0.60 | 0.075~0.110 | 12 | 133.33 | 2 |
| Chlorfenapyr | 0.29 | y = 1.117 + 0.900x | 5.37 | 0.97 | 0.225~0.355 | 72 | 248.28 | 2 |
| Indoxacarb | 5.38 | y = −1.292 + 0.768x | 20.59 | 0.20 | 4.406~6.541 | 40 | 7.43 | 1 |
| Chlorantraniliprole | 0.84 | y = 0.145 + 1.896x | 11.01 | 0.61 | 0.663~1.034 | 30 | 35.71 | 1 |
| Bisultap | 0.30 | y = 1.186 + 2.283x | 8.10 | 0.84 | 0.243~0.366 | 675 | 2250.00 | 2 |
| Lufenuron | 0.91 | y = 0.036 + 0.373x | 10.53 | 0.65 | 0.457~1.867 | 45 | 49.45 | 1 |
| Treatments | Parasitism Rate (%) | Emergence Rate (%) | ||||
|---|---|---|---|---|---|---|
| Concentration (mg L−1) | R1 | R2 | R3 | R1 | R2 | R3 |
| LC40 | 24.83 ± 1.76 cB | 43.97 ± 1.77 cA | --------- | 65.77 ± 2.20 cA | 72.67 ± 2.24 cA | 64.03 ± 2.78 cA |
| LC30 | 34.57 ± 1.79 bB | 50.43 ± 1.24 cA | --------- | 72.9 ± 2.20 bcA | 77.37 ± 1.13 bcA | 72.57 ± 2.28 bcA |
| LC20 | 43.17 ± 2.18 bB | 59.27 ± 1.99 bA | --------- | 80.00 ± 1.93 bA | 82.73 ± 0.90 bA | 78.73 ± 0.84 bA |
| Control | 69.77 ± 2.84 aA | 70.77 ± 1.39 aA | --------- | 89.13 ± 1.24 aA | 88.37 ± 1.04 aA | 88.23 ± 0.76 aA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Su, H.; Lyu, B.; Yan, S.; Lu, H.; Tang, J. Safety Evaluation of Chemical Insecticides to Tetrastichus howardi (Hymenoptera: Eulophidae), a Pupal Parasitoid of Spodoptera frugiperda (Lepidoptera: Noctuidae) Using Three Exposure Routes. Insects 2022, 13, 443. https://doi.org/10.3390/insects13050443
Liu Z, Su H, Lyu B, Yan S, Lu H, Tang J. Safety Evaluation of Chemical Insecticides to Tetrastichus howardi (Hymenoptera: Eulophidae), a Pupal Parasitoid of Spodoptera frugiperda (Lepidoptera: Noctuidae) Using Three Exposure Routes. Insects. 2022; 13(5):443. https://doi.org/10.3390/insects13050443
Chicago/Turabian StyleLiu, Zhuo, Hao Su, Baoqian Lyu, Sanqiang Yan, Hui Lu, and Jihong Tang. 2022. "Safety Evaluation of Chemical Insecticides to Tetrastichus howardi (Hymenoptera: Eulophidae), a Pupal Parasitoid of Spodoptera frugiperda (Lepidoptera: Noctuidae) Using Three Exposure Routes" Insects 13, no. 5: 443. https://doi.org/10.3390/insects13050443