
Testing Phylogeographic Hypotheses in Mepraia (Hemiptera: Reduviidae) Suggests a Complex Spatio-Temporal Colonization in the Coastal Atacama Desert

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples, Localities and Obtaining Mitochondrial Gene Sequences
2.2. Phylogenetic Analyses and Estimates of Time of the Most Recent Common Ancestor (TMRCA)
2.3. Test for Phylogeographic Scenarios Using Coalescent Simulations
2.4. Test of Phylogenetic Hypotheses Using Bayes Factor
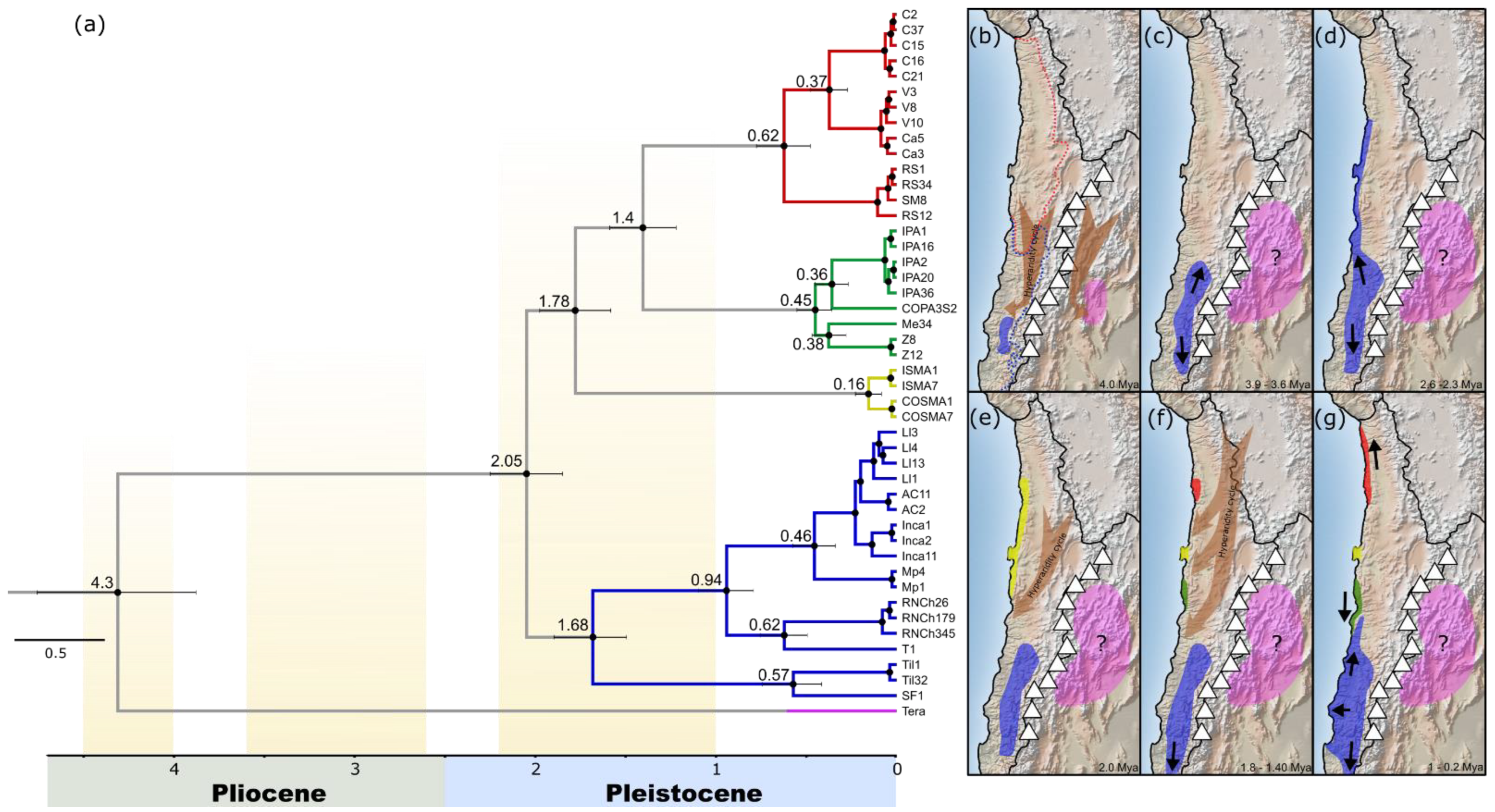
3. Results
3.1. Phylogenetic Relationships and Divergences Time into Mepraia Species
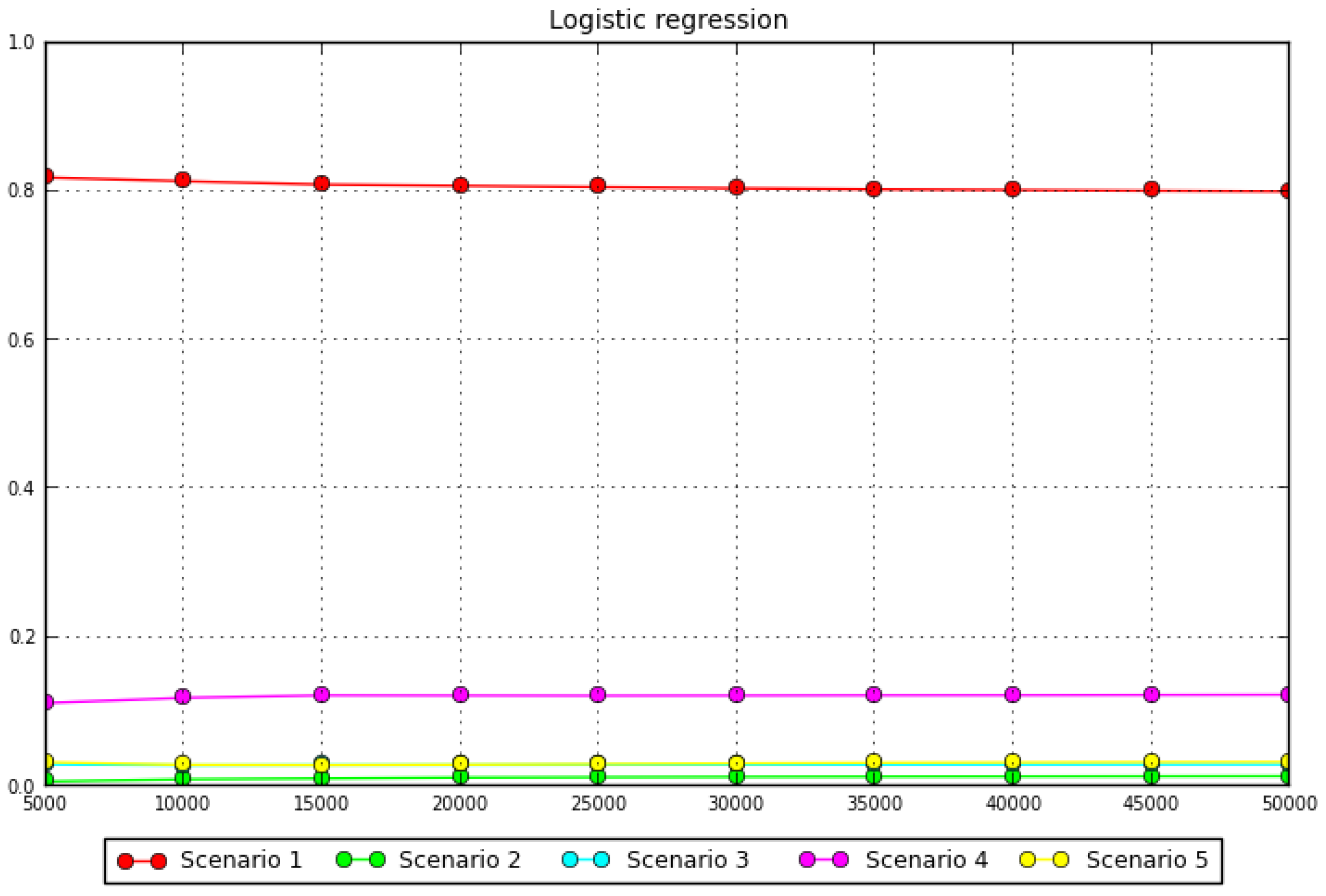
3.2. Test of Phylogeographic Hypotheses with Coalescent Simulations
3.3. Test of Phylogenetic Hypotheses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Graham, A. The Andes: A geological overview from a biological perspective. Ann. Mo. Bot. Gard. 2009, 96, 371–385. [Google Scholar] [CrossRef]
- Hughes, C.; Eastwood, R. Island radiation on a continental scale: Exceptional rates of plant diversification after uplift of the Andes. Proc. Natl. Acad. Sci. USA 2006, 103, 10334–10339. [Google Scholar] [CrossRef] [Green Version]
- Zúñiga-Reinoso, A.; Ritter, B.; Predel, R. The colonization of the Puna and Atacama Biogeographic Province by sister clades of Psectrascelis (Coleoptera: Tenebrionidae): Synchronous expansion without spatial overlap. J. Biogeogr. 2021, 48, 1930–1940. [Google Scholar] [CrossRef]
- Monteiro, F.; Weirauch, C.; Felix, M.; Lazoski, C.; Abad-Franch, F. Evolution, Systematics, and Biogeography of the Triatominae, Vectors of Chagas Disease. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 265–344. [Google Scholar]
- Campos-Soto, R.; Ortiz, S.; Ivan, C.; Bruneau, N.; Botto-Mahan, C.; Solari, A. Interactions between Trypanosoma cruzi the Chagas disease parasite and naturally infected wild Mepraia vectors of Chile. Vector-Borne Zoonotic Dis. 2016, 16, 165–171. [Google Scholar] [CrossRef] [Green Version]
- Botto-Mahan, C.; Bacigalupo, A.; Correa, J.P.; Fontúrbel, F.E.; Cattan, P.E.; Solari, A. Prevalence, infected density or individual probability of infection? Assessing vector infection risk in the wild transmission of Chagas disease. Proc. R. Soc. B Biol. Sci. 2020, 287, 20193018. [Google Scholar] [CrossRef] [Green Version]
- Campos, R.; Torres-Pérez, F.; Botto-Mahan, C.; Coronado, X.; Solari, A. High phylogeographic structure in sylvatic vectors of Chagas disease of the genus Mepraia (Hemiptera: Reduviidae). Infect. Genet. Evol. 2013, 19, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Frías-Lasserre, D. A new species and karyotype variation in the bordering distribution of Mepraia spinolai (Porter) and Mepraia gajardoi Frías et al. (Hemiptera: Reduviidae: Triatominae) in Chile and its parapatric model of speciation. Neotrop. Entomol. 2010, 39, 572–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Soto, R.; Díaz-Campusano, G.; Cianferoni, F.; Torres-Pérez, F. Biogeographic origin and phylogenetic relationships of Mepraia (Hemiptera, Reduviidae) on islands of northern Chile. PLoS ONE 2020, 15, e0234056. [Google Scholar] [CrossRef] [PubMed]
- Campos-Soto, R.; Díaz-Campusano, G.; Quiroga, N.; Muñoz-San Martín, C.; Rives-Blanchard, N.; Torres-Pérez, F. Trypanosoma cruzi-infected triatomines and rodents co-occur in a coastal island of northern Chile. PeerJ 2020, 8, e9967. [Google Scholar] [CrossRef] [PubMed]
- Calleros, L.; Panzera, F.; Bargues, M.D.; Monteiro, F.A.; Klisiowicz, D.R.; Zuriaga, M.A.; Mas-Coma, S.; Pérez, R. Systematics of Mepraia (Hemiptera-Reduviidae): Cytogenetic and molecular variation. Infect. Genet. Evol. 2010, 10, 221–228. [Google Scholar] [CrossRef]
- Grisales, N.; Triana, O.; Angulo, V.; Jaramillo, N.; Parra-Henao, G.; Panzera, F.; Gómez-Palacio, A. Diferenciación genética de tres poblaciones colombianas de Triatoma dimidiata (Latreille, 1811) mediante análisis molecular del gen mitocondrial ND4. Biomédica 2010, 30, 207–214. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Lefort, V.; Longueville, J.E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Morrone, J.J. Biogeographical regionalisation of the Andean region. Zootaxa 2015, 3936, 207–236. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Arensburger, P.; Buckley, T. Biogeography and phylogeny of the New Zealand cicada genera (Hemiptera: Cicadidae) based on nuclear and mitochondrial DNA data. J. Biogeogr. 2004, 31, 557–569. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, F.; Barrett, T.V.; Fitzpatrick, S.; Cordon-Rosales, C.; Feliciangeli, D.; Beard, C.B. Molecular phylogeography of the Amazonian Chagas disease vectors Rhodnius prolixus and R. robustus. Mol. Ecol. 2003, 12, 997–1006. [Google Scholar] [CrossRef]
- Pech-May, A.; Mazariegos-Hidalgo, C.J.; Izeta-Alberdi, A.; López-Cancino, S.A.; Tun-Ku, E.; de la Cruz-Félix, K.; Ibarra-Cerdeña, C.N.; González Ittig, R.E.; Ramsey, J.M. Genetic variation and phylogeography of the Triatoma dimidiata complex evidence a potential center of origin and recent divergence of haplogroups having differential Trypanosoma cruzi and DTU infections. PLoS Negl. Trop. Dis. 2019, 13, e0007044. [Google Scholar] [CrossRef] [Green Version]
- Maturana, P.; Brewer, B.J.; Klaere, S.; Bouckaert, R. Model selection and parameter inference in phylogenetics using Nested Sampling. Syst. Biol. 2019, 68, 219–233. [Google Scholar] [CrossRef]
- Kass, R.E.; Raftery, A.E. Bayes Factors. J. Am. Stat. Assoc. 1995, 90, 773–795. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. Molecular Evolution, Phylogenetics and Epidemiology. FigTree. Tree Figure Drawing Tool Software v1.4.4. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 18 February 2022).
- Cornuet, J.-M.; Pudlo, P.; Veyssier, J.; Dehne-Garcia, A.; Gautier, M.; Leblois, R.; Marin, J.-M.; Estoup, A. DIYABC v2.0: A software to make approximate Bayesian computation inferences about population history using single nucleotide polymorphism, DNA sequence and microsatellite data. Bioinformatics 2014, 30, 1187–1189. [Google Scholar] [CrossRef] [Green Version]
- Jordan, T.E.; Kirk-Lawlor, N.E.; Blanco, N.P.; Rech, J.A.; Cosentino, N.J. Landscape modification in response to repeated onset of hyperarid paleoclimate states since 14 Ma, Atacama Desert, Chile. GSA Bull. 2014, 126, 1016–1046. [Google Scholar] [CrossRef]
- Hinojosa, L.F.; Villagrán, C. Historia de los bosques del sur de Sudamérica, I: Antecedentes paleobotánicos, geológicos y climáticos del Terciario del cono sur de América. Rev. Chil. Hist. Nat. 1997, 70, 225–239. [Google Scholar]
- Latorre, C.; Moreno, P.I.; Vargas, G.; Maldonado, A.; Villa-Martínez, R.; Armesto, J.J.; Villagrán, C.; Pino, M.; Núñez, L.; Grosjean, M. Late Quaternary environments and palaeoclimate. In The Geology of Chile; Moreno, T., Gibbons, W., Eds.; Geological Society of London: London, UK, 2007; pp. 309–328. [Google Scholar]
- Zúñiga-Reinoso, Á.; Predel, R. Past climatic changes and their effects on the phylogenetic pattern of the Gondwanan relict Maindronia (Insecta: Zygentoma) in the Chilean Atacama Desert. Glob. Planet. Chang. 2019, 182, 103007. [Google Scholar] [CrossRef]
- Boric-Bargetto, D.; Zúñiga-Reinoso, Á.; Inostroza-Michel, O.; Rodríguez-Serrano, E.; González-Acuña, D.; Palma, R.E.; Hernández, C.E. A comprehensive overview of the genetic diversity in Thylamys elegans (Didelphimorphia: Didelphidae): Establishing the phylogeographic determinants. Rev. Chil. Hist. Nat. 2021, 94, 5. [Google Scholar] [CrossRef]
- Ceccarelli, F.S.; Ojanguren-Affilastro, A.A.; Ramírez, M.J.; Ochoa, J.A.; Mattoni, C.I.; Prendini, L. Andean uplift drives diversification of the bothriurid scorpion genus Brachistosternus. J. Biogeogr. 2016, 43, 1942–1954. [Google Scholar] [CrossRef]
- Ortiz-Jaureguizar, E.; Cladera, G.A. Paleoenvironmental evolution of southern South America during the Cenozoic. J. Arid Environ. 2006, 66, 498–532. [Google Scholar] [CrossRef]
- Garzione, C.N.; McQuarrie, N.; Perez, N.D.; Ehlers, T.A.; Beck, S.L.; Kar, N.; Eichelberger, N.; Chapman, A.D.; Ward, K.M.; Ducea, M.N.; et al. Tectonic Evolution of the Central Andean Plateau and Implications for the Growth of Plateaus. Annu. Rev. Earth Planet. Sci. 2017, 45, 529–559. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, M.; Sloan, L.; Thomas, E.; Billups, K. Trends, Rhythms, and Aberrations in Global Climate 65 Ma to Present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef]
- Glade-Vargas, N.S.; Rojas, C.; Jara-Arancio, P.; Vidal, P.; Arroyo, M.T.K.; Hinojosa, L.F. Biogeography of Argylia D. Don (Bignoniaceae): Diversification, Andean Uplift and Niche Conservatism. Front. Plant Sci. 2021, 12, 2270. [Google Scholar] [CrossRef]
- San Juan, E.; Araya-Donoso, R.; Véliz, D.; Quiroga, N.; Botto-Mahan, C. Genetic diversity in a restricted-dispersal kissing bug: The centre–periphery hypothesis halfway. Mol. Ecol. 2021, 30, 4660–4672. [Google Scholar] [CrossRef]
- Canals, M.; Cruzat, L.; Molina, M.C.; Ferreira, A.; Cattan, P.E. Blood host sources of Mepraia spinolai (Heteroptera: Reduviidae), wild vector of Chagas Disease in Chile. J. Med. Entomol. 2001, 38, 303–307. [Google Scholar] [CrossRef] [Green Version]
- Quiroga, N.; Correa, J.P.; Campos-Soto, R.; San Juan, E.; Araya-Donoso, R.; Díaz-Campusano, G.; González, C.R.; Botto-Mahan, C. Blood-Meal Sources and Trypanosoma cruzi Infection in Coastal and Insular Triatomine Bugs from the Atacama Desert of Chile. Microorganisms 2022, 10, 785. [Google Scholar] [CrossRef]
- Torres-Pérez, F.; Acuna-Retamar, M.; Cook, J.A.; Bacigalupo, A.; García, A.; Cattan, P.E. Statistical phylogeography of Chagas disease vector Triatoma infestans: Testing biogeographic hypotheses of dispersal. Infect. Genet. Evol. 2011, 11, 167–174. [Google Scholar] [CrossRef]
- Bacigalupo, A.; Torres-Pérez, F.; Segovia, V.; García, A.; Correa, J.P.; Moreno, L.; Arroyo, P.; Cattan, P.E. Sylvatic foci of the Chagas disease vector Triatoma infestans in Chile: Description of a new focus and challenges for control programs. Memórias do Instituto Oswaldo Cruz 2010, 105, 633–641. [Google Scholar] [CrossRef] [Green Version]
- Baranzelli, M.C.; Johnson, L.A.; Cosacov, A.; Sérsic, A.N. Historical and ecological divergence among populations of Monttea chilensis (Plantaginaceae), an endemic endangered shrub bordering the Atacama Desert, Chile. Evol. Ecol. 2014, 28, 751–774. [Google Scholar] [CrossRef]
- Mayle, F.E.; Burbridge, R.; Killeen, T.J. Millennial-Scale Dynamics of Southern Amazonian Rain Forests. Science 2000, 290, 2291–2294. [Google Scholar] [CrossRef] [Green Version]
- Dézerald, O.; Latorre, C.; Betancourt, J.L.; Brito Vera, G.A.; González, A.L. Ecological fidelity and spatiotemporal resolution of arthropod death assemblages from rodent middens in the central Atacama Desert (northern Chile). Quat. Sci. Rev. 2019, 210, 15–25. [Google Scholar] [CrossRef]
- Latorre, C.; Betancourt, J.L.; Rylander, K.A.; Quade, J. Vegetation invasions into absolute desert: A 45;th000 yr rodent midden record from the Calama–Salar de Atacama basins, northern Chile (lat 22°–24°S). GSA Bull. 2002, 114, 349–366. [Google Scholar] [CrossRef]
- Zúñiga-Reinoso, Á.; Pinto, P.; Predel, R. A New Species of Gyriosomus Guérinmeneville (Coleoptera: Tenebrionidae) from the Chilean Atacama Desert. Ann. Zool. 2019, 69, 105–112. [Google Scholar] [CrossRef]
- Guerrero, P.C.; Rosas, M.; Arroyo, M.T.K.; Wiens, J.J. Evolutionary lag times and recent origin of the biota of an ancient desert (Atacama–Sechura). Proc. Natl. Acad. Sci. USA 2013, 110, 11469–11474. [Google Scholar] [CrossRef] [Green Version]
- Rundel, P.; Dillon, M.; Palma, B.; Mooney, H.; Gulmon, S.; Ehleringer, J. The Phytogeography and Ecology of the Coastal Atacama and Peruvian Deserts. Aliso A J. Syst. Florist. Bot. 1991, 13, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.; Barichivich, J.; Christie, D.A.; Maldonado, A. Interannual variability of the coastal fog at Fray Jorge relict forests in semiarid Chile. J. Geophys. Res. 2008, 113, G04011. [Google Scholar] [CrossRef] [Green Version]
- Garreaud, R.; Molina, A.; Farias, M. Andean uplift, ocean cooling and Atacama hyperaridity: A climate modeling perspective. Earth Planet. Sci. Lett. 2010, 292, 39–50. [Google Scholar] [CrossRef]
- Pedoja, K.; Husson, L.; Johnson, M.E.; Melnick, D.; Witt, C.; Pochat, S.; Nexer, M.; Delcaillau, B.; Pinegina, T.; Poprawski, Y.; et al. Earth-Science Reviews Coastal staircase sequences reflecting sea-level oscillations and tectonic uplift during the Quaternary and Neogene. Earth Sci. Rev. 2014, 132, 13–38. [Google Scholar] [CrossRef]
- Ortlieb, L.; Zazo, C.; Goy, J.L.; Hillaire-Marcel, C.; Ghaleb, B.; Cournoyer, L. Coastal deformation and sea-level changes in the northern Chile subduction area (23 degrees S) during the last 330 ky. Quat. Sci. Rev. 1996, 15, 819–831. [Google Scholar] [CrossRef]
- Quezada, J.; González, G.; Dunai, T.; Jensen, A.; Juez-Larré, J. Alzamiento litoral Pleistoceno del norte de Chile: Edades 21Ne de la terraza costera más alta del área de Caldera-Bahía Inglesa. Rev. Geológica de Chile 2007, 34, 81–96. [Google Scholar] [CrossRef]
- Binnie, A.; Dunai, T.J.; Binnie, S.A.; Victor, P.; González, G.; Bolten, A. Accelerated late quaternary uplift revealed by 10Be exposure dating of marine terraces, Mejillones Peninsula, northern Chile. Quat. Geochronol. 2016, 36, 12–27. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S1 | S2 | S3 | S4 | S5 | |
|---|---|---|---|---|---|
| S1 | −8647.2 | 38.5 * | 23 * | 18.1 * | 8.8 * |
| S2 | −38.5 | −8685.7 | −15.5 | −20.4 | −29.7 |
| S3 | −23 | 15.5 | −8670.2 | −4.9 | −14.2 |
| S4 | −18.1 | 20.4 | 4.9 | −8665.3 | −9.3 |
| S5 | −8.8 | 29.7 | 14.2 | 9.3 | −8656 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campos-Soto, R.; Rodríguez-Valenzuela, E.; Díaz-Campusano, G.; Boric-Bargetto, D.; Zúñiga-Reinoso, Á.; Cianferoni, F.; Torres-Pérez, F. Testing Phylogeographic Hypotheses in Mepraia (Hemiptera: Reduviidae) Suggests a Complex Spatio-Temporal Colonization in the Coastal Atacama Desert. Insects 2022, 13, 419. https://doi.org/10.3390/insects13050419
Campos-Soto R, Rodríguez-Valenzuela E, Díaz-Campusano G, Boric-Bargetto D, Zúñiga-Reinoso Á, Cianferoni F, Torres-Pérez F. Testing Phylogeographic Hypotheses in Mepraia (Hemiptera: Reduviidae) Suggests a Complex Spatio-Temporal Colonization in the Coastal Atacama Desert. Insects. 2022; 13(5):419. https://doi.org/10.3390/insects13050419
Chicago/Turabian StyleCampos-Soto, Ricardo, Evelyn Rodríguez-Valenzuela, Gabriel Díaz-Campusano, Dusan Boric-Bargetto, Álvaro Zúñiga-Reinoso, Franco Cianferoni, and Fernando Torres-Pérez. 2022. "Testing Phylogeographic Hypotheses in Mepraia (Hemiptera: Reduviidae) Suggests a Complex Spatio-Temporal Colonization in the Coastal Atacama Desert" Insects 13, no. 5: 419. https://doi.org/10.3390/insects13050419