
Cyromazine Effects the Reproduction of Drosophila by Decreasing the Number of Germ Cells in the Female Adult Ovary


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Drosophila Strain and Breeding Conditions
2.2. Insecticide Treatment
2.3. Immunohistochemistry and Microscopy
2.4. Counting the Number of GSCs and CBs in the Ovary of 3-Day Old Female Flies
2.5. Effect of Cyromazine on the Expression of Ecdysone Signaling Related Key Genes
2.6. Ecdysone Titer Measurement
2.7. Effect of Cyromazine on the Ovary Morphology and Fecundity
2.8. Statistical Analysis
3. Results
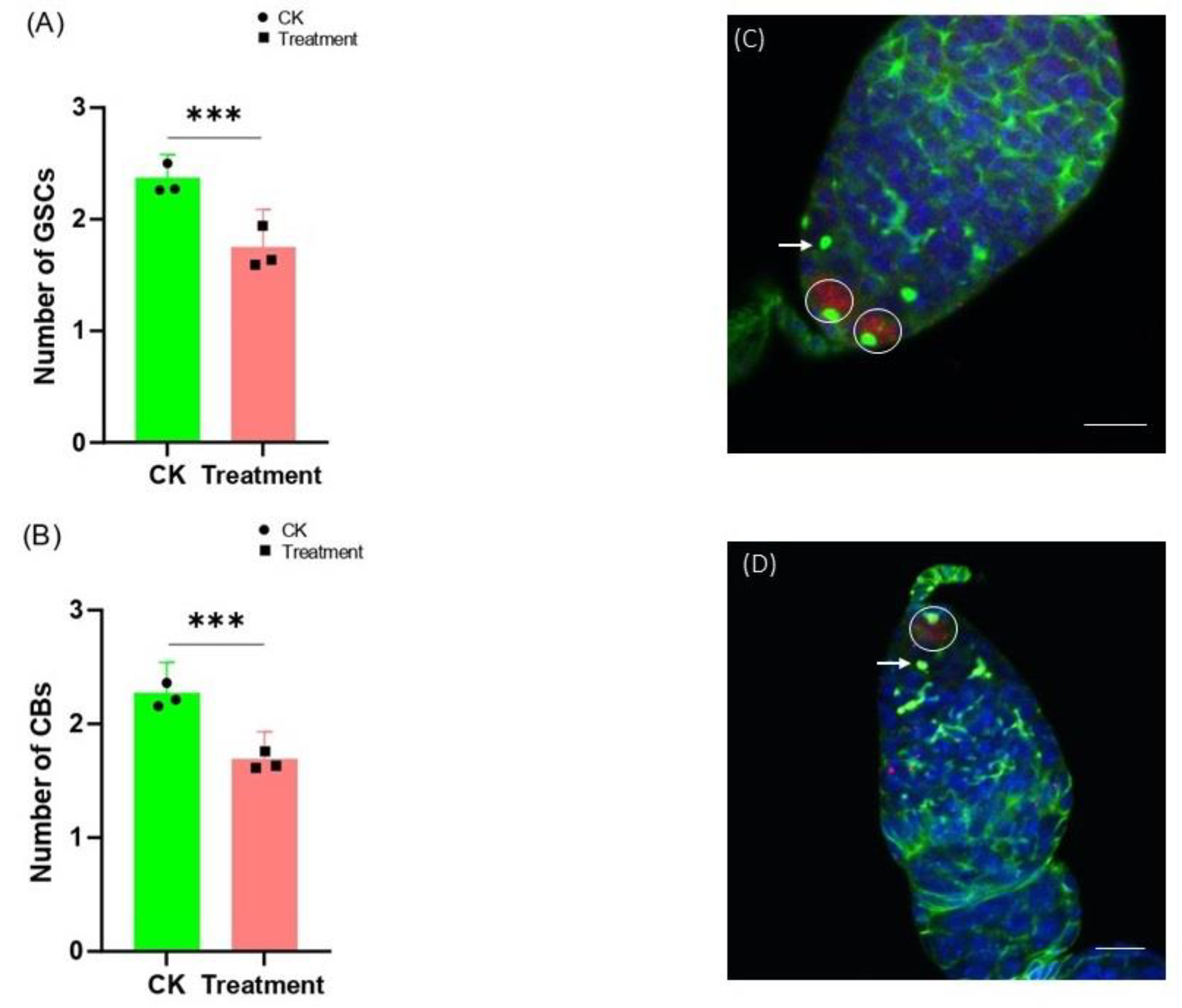
3.1. Cyromazine Effects the GSCs and CBs in the 3-Day Old Female Adult Ovary
3.2. Expression of Ecdysone Signaling Related Key Genes
3.3. Cyromazine Effects the Ecdysone Titer in Both the Larval and Adult Ovaries
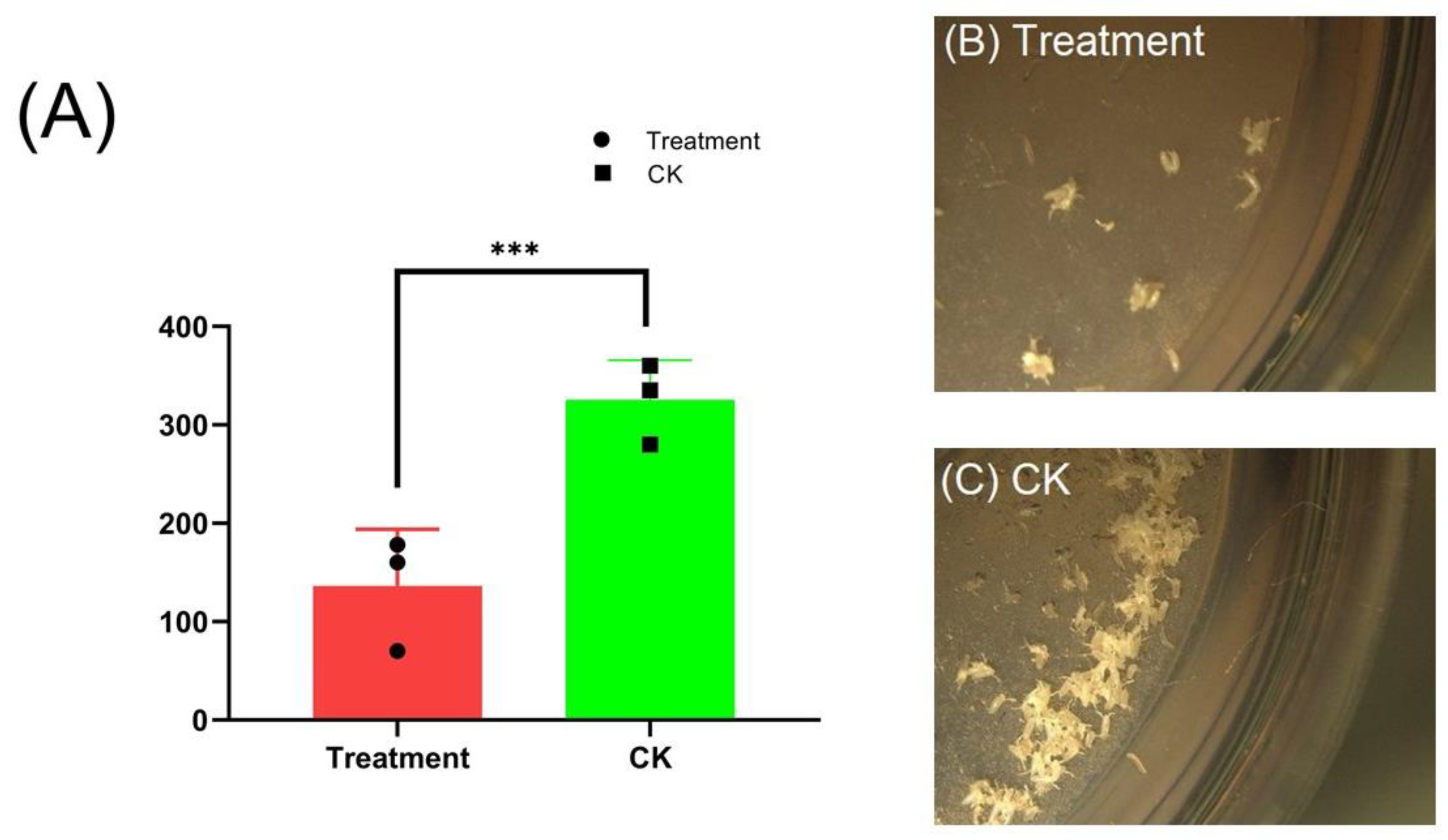
3.4. Effect on the Ovary Morphology and Fecundity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lengil, T.; Gancz, D.; Gilboa, L. Activin signaling balances proliferation and differentiation of ovarian niche precursors and enables adjustment of niche numbers. Development 2015, 142, 883–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Tu, R.; Duan, B.; Song, X.; Chen, S.; Scott, A.; Hall, K.; Blanck, J.; DeGraffenreid, D.; Li, H.; Perera, A. Multiple niche compartments orchestrate stepwise germline stem cell progeny differentiation. Curr. Biol. 2021, 31, 827–839.e3. [Google Scholar] [CrossRef]
- Dansereau, D.A.; Lasko, P. The development of germline stem cells in Drosophila. Germline Stem Cells 2008, 450, 3–26. [Google Scholar]
- Kirilly, D.; Wang, S.; Xie, T. Self-maintained escort cells form a germline stem cell differentiation niche. Development 2011, 138, 5087–5097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rust, K.; Byrnes, L.E.; Yu, K.S.; Park, J.S.; Sneddon, J.B.; Tward, A.D.; Nystul, T.G. A single-cell atlas and lineage analysis of the adult Drosophila ovary. Nat. Commun. 2020, 11, 5628. [Google Scholar] [CrossRef]
- Page, S.W. Chapter 10-Antiparasitic drugs. In Small Animal Clinical Pharmacology, 2nd ed.; Maddison, J.E., Page, S.W., Church, D.B., Eds.; W.B. Saunders: Edinburgh, UK, 2008; pp. 198–260. [Google Scholar]
- National Center for Biotechnology Information. PubChem Compound Summary for CID 47866, Cyromazine. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Cyromazine (accessed on 14 April 2022).
- Taylor, D.B.; Friesen, K.; Zhu, J.J.; Sievert, K. Efficacy of cyromazine to control immature stable flies (Diptera: Muscidae) developing in winter hay feeding sites. J. Econ. Entomol. 2012, 105, 726–731. [Google Scholar] [CrossRef]
- Van De Wouw, A.P.; Batterham, P.; Daborn, P.J. The insect growth regulator insecticide cyromazine causes earlier emergence in Drosophila melanogaster. Arch. Insect Biochem. Physiol. 2006, 63, 101–109. [Google Scholar] [CrossRef]
- Uryu, O.; Ameku, T.; Niwa, R. Recent progress in understanding the role of ecdysteroids in adult insects: Germline development and circadian clock in the fruit fly Drosophila melanogaster. Zool. Lett. 2015, 1, 32. [Google Scholar] [CrossRef] [Green Version]
- Khalid, M.Z.; Ahmad, S.; Ngegba, P.M.; Zhong, G. Role of Endocrine System in the Regulation of Female Insect Reproduction. Biology 2021, 10, 614. [Google Scholar] [CrossRef]
- Belles, X.; Piulachs, M.-D. Ecdysone signalling and ovarian development in insects: From stem cells to ovarian follicle formation. Biochim. Biophys. Acta Gene Regul. Mech. 2015, 1849, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Ables, E.T.; Drummond-Barbosa, D. The steroid hormone ecdysone functions with intrinsic chromatin remodeling factors to control female germline stem cells in Drosophila. Cell Stem Cell 2010, 7, 581–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ables, E.T.; Hwang, G.H.; Finger, D.S.; Hinnant, T.D.; Drummond-Barbosa, D. A genetic mosaic screen reveals ecdysone-responsive genes regulating Drosophila oogenesis. Genes Genomes Genet. 2016, 6, 2629–2642. [Google Scholar] [CrossRef]
- Buszczak, M.; Freeman, M.R.; Carlson, J.R.; Bender, M.; Cooley, L.; Segraves, W.A. Ecdysone response genes govern egg chamber development during mid-oogenesis in Drosophila. Development 1999, 126, 4581–4589. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Chepelev, I.; Wei, G.; Tarayrah, L.; Cui, K.; Zhao, K.; Chen, X. Dynamic regulation of alternative splicing and chromatin structure in Drosophila gonads revealed by RNA-seq. Cell Res. 2010, 20, 763–783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durham, M.F.; Magwire, M.M.; Stone, E.A.; Leips, J. Genome-wide analysis in Drosophila reveals age-specific effects of SNPs on fitness traits. Nat. Commun. 2014, 5, 4338. [Google Scholar] [CrossRef] [Green Version]
- Ables, E.T.; Drummond-Barbosa, D. Cyclin E Controls Drosophila female germline stem cell maintenance independently of its role in proliferation by modulating responsiveness to niche signals. Development 2013, 140, 530–540. [Google Scholar] [CrossRef] [Green Version]
- Yan, D.; Neumüller, R.A.; Buckner, M.; Ayers, K.; Li, H.; Hu, Y.; Yang-Zhou, D.; Pan, L.; Wang, X.; Kelley, C. A regulatory network of Drosophila germline stem cell self-renewal. Dev. Cell 2014, 28, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Xu, Y.; Lin, X. Role of Broad-Complex (Br) and Krüppel homolog 1 (Kr-h1) in the ovary development of Nilaparvata lugens. Front. Physiol. 2017, 8, 1013. [Google Scholar] [CrossRef]
- Zhu, Z.; Tong, C.; Qiu, B.; Yang, H.; Xu, J.; Zheng, S.; Song, Q.; Feng, Q.; Deng, H. 20E-mediated regulation of BmKr-h1 by BmKRP promotes oocyte maturation. BMC Biol. 2021, 19, 39. [Google Scholar] [CrossRef]
- Bastock, R.; St Johnston, D. Drosophila oogenesis. Curr. Biol. 2008, 18, R1082–R1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Zhong, G.; Chai, P.C.; Luo, L.; Liu, S.; Yang, Y.; Baeg, G.-H.; Cai, Y. Coordinated niche-associated signals promote germline homeostasis in the Drosophila ovary. J. Cell Biol. 2015, 211, 469–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafar, J.; Zhang, Y.; Huang, J.; Freed, S.; Shoukat, R.F.; Xu, X.; Jin, F. Spatio-Temporal Profiling of Metarhizium anisopliae—Responsive microRNAs Involved in Modulation of Plutella xylostella Immunity and Development. J. Fungi 2021, 7, 942. [Google Scholar] [CrossRef] [PubMed]
- Zafar, J.; Shoukat, R.F.; Zhang, Y.; Freed, S.; Xu, X.; Jin, F. Metarhizium anisopliae challenges immunity and demography of Plutella xylostella. Insects 2020, 11, 694. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Sun, T.; Sun, Z.; Li, H.; Qi, X.; Zhong, G.; Yi, X. Azadirachtin acting as a hazardous compound to induce multiple detrimental effects in Drosophila melanogaster. J. Hazard. Mater. 2018, 359, 338–347. [Google Scholar] [CrossRef]
- König, A.; Yatsenko, A.S.; Weiss, M.; Shcherbata, H.R. Ecdysteroids affect Drosophila ovarian stem cell niche formation and early germline differentiation. EMBO J. 2011, 30, 1549–1562. [Google Scholar] [CrossRef] [Green Version]
- Enya, S.; Ameku, T.; Igarashi, F.; Iga, M.; Kataoka, H.; Shinoda, T.; Niwa, R. A Halloween gene noppera-bo encodes a glutathione S-transferase essential for ecdysteroid biosynthesis via regulating the behaviour of cholesterol in Drosophila. Sci. Rep. 2014, 4, 6586. [Google Scholar] [CrossRef] [Green Version]
- Chávez, V.M.; Marqués, G.; Delbecque, J.P.; Kobayashi, K.; Hollingsworth, M.; Burr, J.; Natzle, J.E.; O’Connor, M.B. The Drosophila disembodied gene controls late embryonic morphogenesis and codes for a cytochrome P450 enzyme that regulates embryonic ecdysone levels. Development 2000, 127, 4115–4126. [Google Scholar] [CrossRef]
- Romani, P.; Bernardi, F.; Hackney, J.; Dobens, L.; Gargiulo, G.; Cavaliere, V. Cell survival and polarity of Drosophila follicle cells require the activity of ecdysone receptor B1 isoform. Genetics 2009, 181, 165–175. [Google Scholar] [CrossRef] [Green Version]
- Lenaerts, C.; Marchal, E.; Peeters, P.; Vanden Broeck, J. The ecdysone receptor complex is essential for the reproductive success in the female desert locust, Schistocerca gregaria. Sci. Rep. 2019, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Ameku, T.; Niwa, R. Mating-induced increase in germline stem cells via the neuroendocrine system in female Drosophila. PLoS Genet. 2016, 12, e1006123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domanitskaya, E.; Anllo, L.; Schüpbach, T. Phantom, a cytochrome P450 enzyme essential for ecdysone biosynthesis, plays a critical role in the control of border cell migration in Drosophila. Dev. Biol. 2014, 386, 408–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, M.P.; Yin, C.M.; Tatar, M. Impaired ovarian ecdysone synthesis of Drosophila melanogaster insulin receptor mutants. Aging Cell 2002, 1, 158–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameku, T.; Yoshinari, Y.; Fukuda, R.; Niwa, R. Ovarian ecdysteroid biosynthesis and female germline stem cells. Fly 2017, 11, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Morris, L.X.; Spradling, A.C. Steroid signaling within Drosophila ovarian epithelial cells sex-specifically modulates early germ cell development and meiotic entry. PLoS ONE 2012, 7, e46109. [Google Scholar] [CrossRef] [Green Version]
- König, A.; Shcherbata, H.R. Soma influences GSC progeny differentiation via the cell adhesion-mediated steroid-let-7-Wingless signaling cascade that regulates chromatin dynamics. Biol. Open 2015, 4, 285–300. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Zhu, G.; Zhao, H.; Wang, Z.; Xue, M.; Li, X.; Xu, H.; Ma, X.; Liu, Y. Sterilization effects of adult-targeted baits containing insect growth regulators on Delia antiqua. Sci. Rep. 2016, 6, 32855. [Google Scholar] [CrossRef]
- Park, K.; Kwak, I.-S. Disrupting effects of antibiotic sulfathiazole on developmental process during sensitive life-cycle stage of Chironomus riparius. Chemosphere 2018, 190, 25–34. [Google Scholar] [CrossRef]
- Yao, Q.; Xu, S.; Dong, Y.; Quan, L.; Chen, B. Ecdysone receptor (EcR) and ultraspiracle protein (USP) genes from Conopomorpha sinensis Bradley eggs: Identification and expression in response to insecticides. Front. Physiol. 2020, 11, 851. [Google Scholar] [CrossRef]
- Yoshiyama, T.; Namiki, T.; Mita, K.; Kataoka, H.; Niwa, R. Neverland is an evolutionally conserved Rieske-domain protein that is essential for ecdysone synthesis and insect growth. Development 2006, 133, 2565–2574. [Google Scholar] [CrossRef] [Green Version]
- Carney, G.E.; Bender, M. The Drosophila ecdysone receptor (EcR) gene is required maternally for normal oogenesis. Genetics 2000, 154, 1203–1211. [Google Scholar] [CrossRef] [PubMed]
- Ting, X. Control of germline stem cell self-renewal and differentiation in the Drosophila ovary: Concerted actions of niche signals and intrinsic factors. Wiley Interdiscip. Rev. Dev. Biol. 2013, 2, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Zipper, L.; Jassmann, D.; Burgmer, S.; Görlich, B.; Reiff, T. Ecdysone steroid hormone remote controls intestinal stem cell fate decisions via the PPARγ-homolog Eip75B in Drosophila. eLife 2020, 9, e55795. [Google Scholar] [CrossRef] [PubMed]
- Yatsenko, A.S.; Shcherbata, H.R. Stereotypical architecture of the stem cell niche is spatiotemporally established by miR-125-dependent coordination of Notch and steroid signaling. Development 2018, 145, dev159178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gleason, R.J.; Anand, A.; Kai, T.; Chen, X. Protecting and diversifying the germline. Genetics 2018, 208, 435–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayukawa, T.; Jouraku, A.; Ito, Y.; Shinoda, T. Molecular mechanism underlying juvenile hormone-mediated repression of precocious larval–adult metamorphosis. Proc. Natl. Acad. Sci. USA 2017, 114, 1057–1062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Song, W.; Li, Z.; Qian, W.; Wei, L.; Yang, Y.; Wang, W.; Zhou, X.; Meng, M.; Peng, J. Krüppel homolog 1 represses insect ecdysone biosynthesis by directly inhibiting the transcription of steroidogenic enzymes. Proc. Natl. Acad. Sci. USA 2018, 115, 3960–3965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ojani, R.; Fu, X.; Ahmed, T.; Liu, P.; Zhu, J. Krüppel homologue 1 acts as a repressor and an activator in the transcriptional response to juvenile hormone in adult mosquitoes. Insect Mol. Biol. 2018, 27, 268–278. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khalid, M.Z.; Sun, Z.; Chen, Y.; Zhang, J.; Zhong, G. Cyromazine Effects the Reproduction of Drosophila by Decreasing the Number of Germ Cells in the Female Adult Ovary. Insects 2022, 13, 414. https://doi.org/10.3390/insects13050414
Khalid MZ, Sun Z, Chen Y, Zhang J, Zhong G. Cyromazine Effects the Reproduction of Drosophila by Decreasing the Number of Germ Cells in the Female Adult Ovary. Insects. 2022; 13(5):414. https://doi.org/10.3390/insects13050414
Chicago/Turabian StyleKhalid, Muhammad Zaryab, Zhipeng Sun, Yaoyao Chen, Jing Zhang, and Guohua Zhong. 2022. "Cyromazine Effects the Reproduction of Drosophila by Decreasing the Number of Germ Cells in the Female Adult Ovary" Insects 13, no. 5: 414. https://doi.org/10.3390/insects13050414