
Wing Phenotypic Variation among Stomoxys calcitrans (Diptera: Muscidae) Populations in Thailand



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
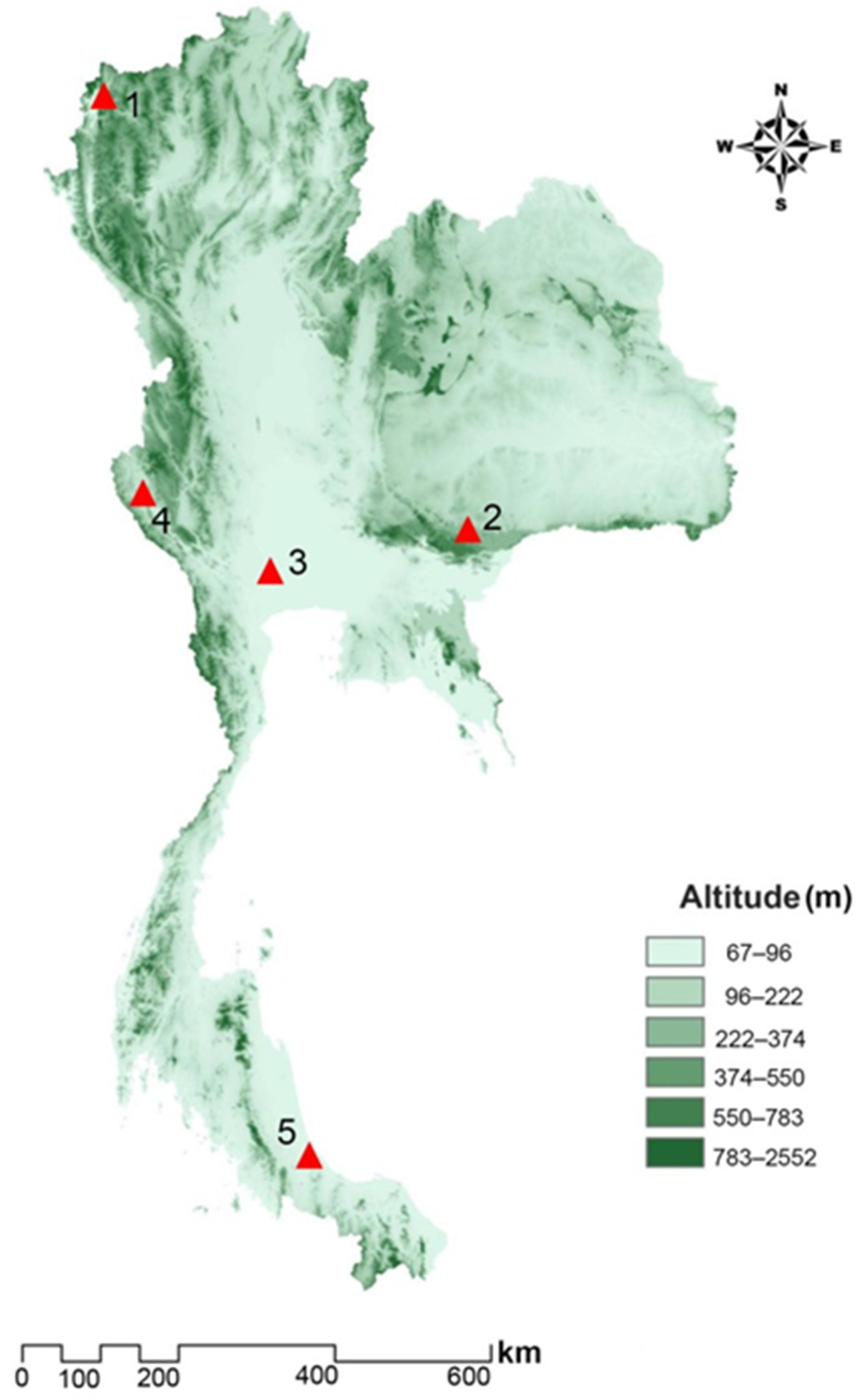
2.2. Stable Fly Specimens
2.3. Sample Preparation and Landmark Digitization
2.4. Repeatability and Allometry
2.5. Wing Size Analyses
2.6. Wing Shape Analyses
2.7. Validated Classification
3. Results
3.1. Repeatability of Wing Image
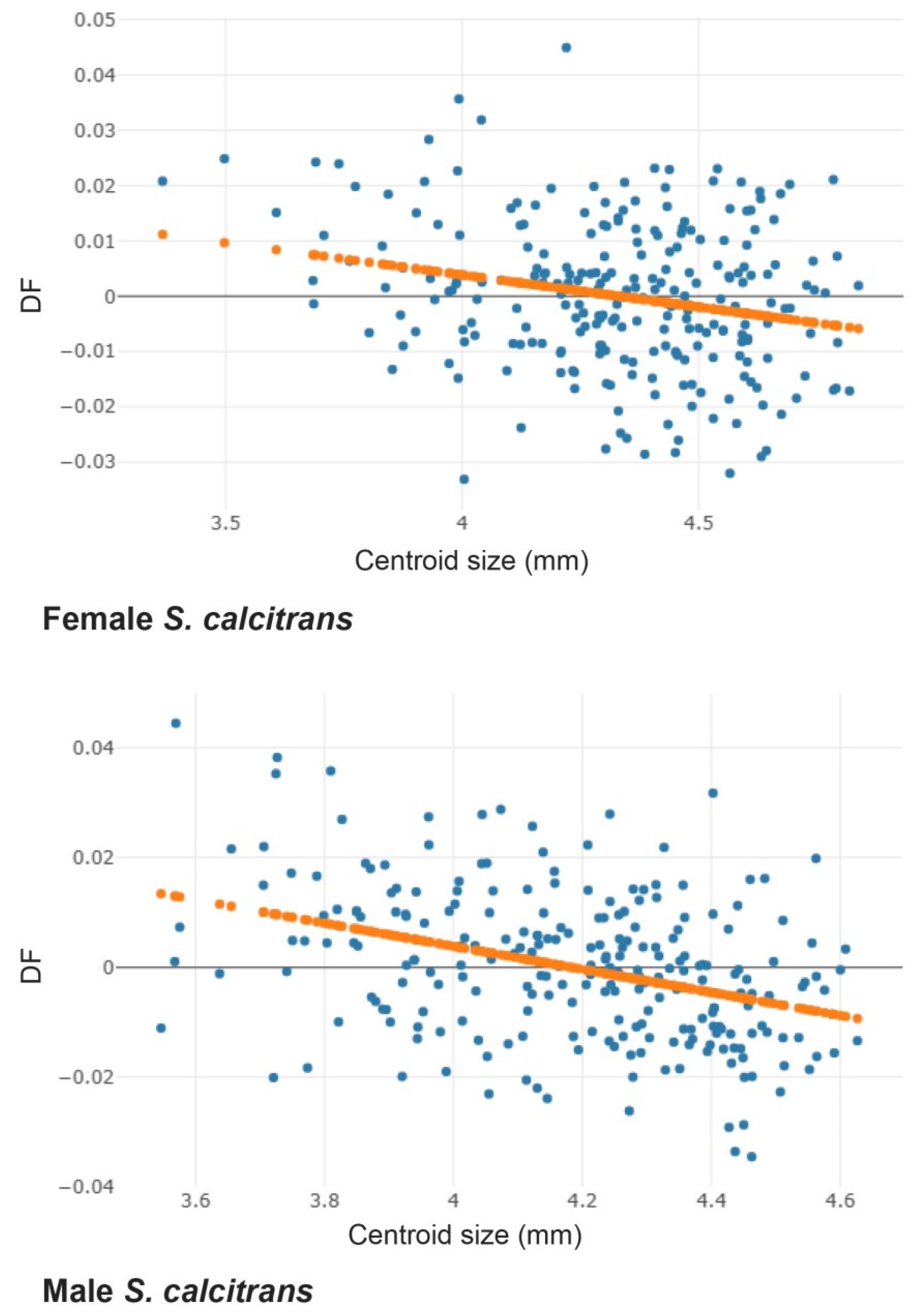
3.2. Allometric Effect
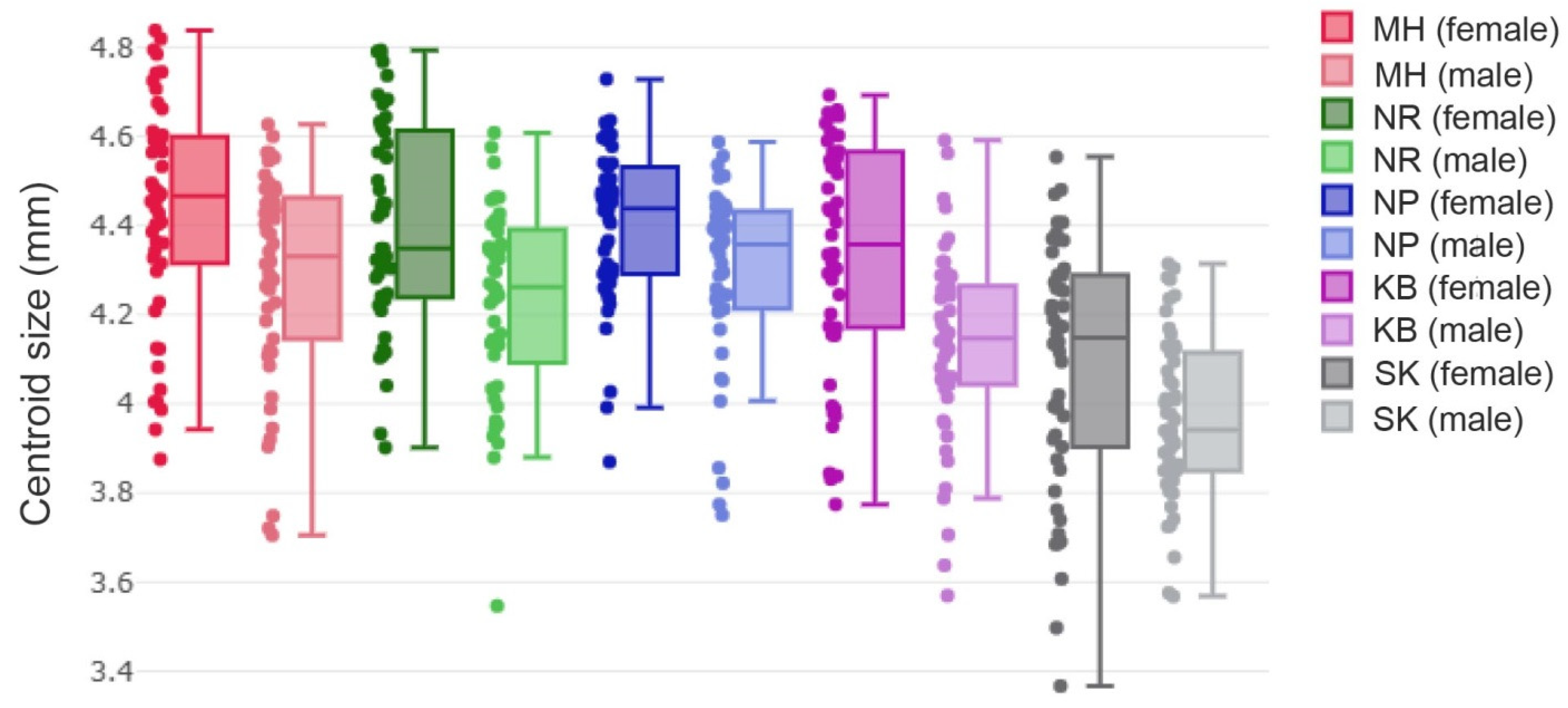
3.3. Wing Size Variation
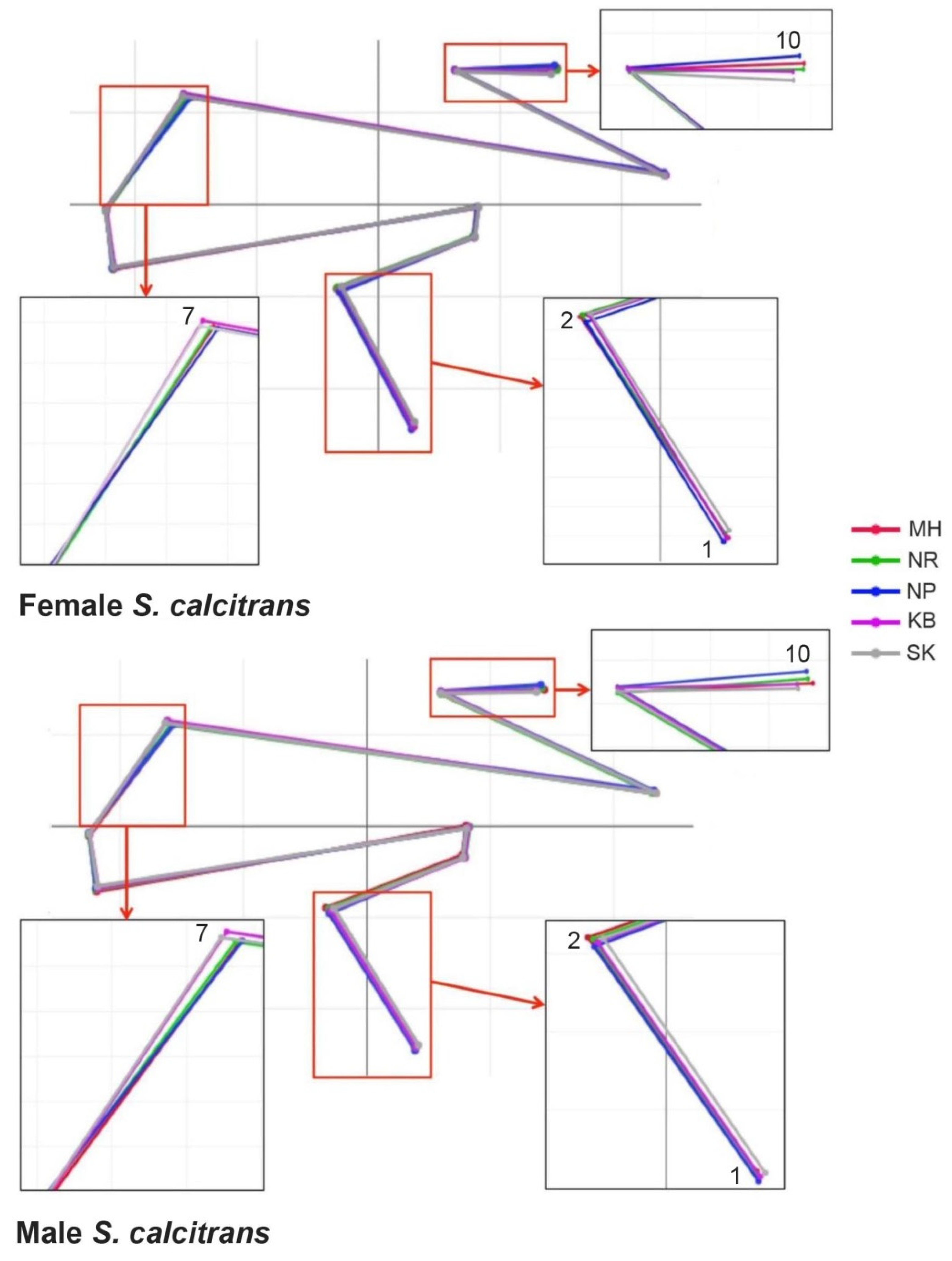
3.4. Wing Shape Variation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zumpt, F. The Stomoxyine Biting Flies of the World; Gustav Fisher Verlag: Stuttgart, Germany, 1973; pp. 97–137. [Google Scholar]
- Baldacchino, F.; Muenworn, V.; Desquesnes, M.; Desoli, F.; Charoenviriyaphap, T.; Duvallet, G. Transmission of pathogens by Stomoxys flies (Diptera, Muscidae): A review. Parasite 2013, 20, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibbs, E.P.J.; Johnson, R.H.; Gatehouse, A.G. A laboratory technique for studying the mechanical transmission of bovine herpes mammillitis virus by the stable fly (Stomoxys calcitrans L.). Res. Vet. Sci. 1973, 14, 145–147. [Google Scholar] [PubMed]
- Foil, L.D.; Meek, C.L.; Adams, W.V.; Issel, C.J. Mechanical transmission of equine infectious anemia virus by deer flies (Chrysops flavidus) and stable flies (Stomoxys calcitrans). Am. J. Vet. Res. 1983, 44, 155–156. [Google Scholar] [PubMed]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Campe, W.V.; De Vleeschauwer, A.; Tuppurainen, E.S.M.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical lumpy skin disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 20076. [Google Scholar] [CrossRef]
- Vergne, T.; Andraud, M.; Bonnet, S.; De Regge, N.; Desquesnes, M.; Fite, J.; Etore, F.; Garigliany, M.M.; Jori, F.; Lempereur, L.; et al. Mechanical transmission of African swine fever virus by Stomoxys calcitrans: Insights from a mechanistic model. Transbound. Emerg. Dis. 2021, 68, 1541–1549. [Google Scholar] [CrossRef]
- Desquesnes, M.; Holzmuller, P.; Lai, D.H.; Dargantes, A.; Lun, Z.R.; Jittaplapong, S. Trypanosoma evansi and surra: A review and perspectives on origin, history, distribution, taxonomy, morphology, hosts, and pathogenic effects. Biomed. Res. Int. 2013, 2013, 194176. [Google Scholar] [CrossRef] [Green Version]
- Sharif, S.; Jacquiet, P.; Prevot, F.; Grisez, C.; Raymond-Letron, I.; Semin, M.O.; Geffré, A.; Trumel, C.; Franc, M.; Bouhsira, É.; et al. Stomoxys calcitrans, mechanical vector of virulent Besnoitia besnoiti from chronically infected cattle to susceptible rabbit. Med. Vet. Entomol. 2019, 33, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Turell, M.J.; Knudson, G.B. Mechanical transmission of Bacillus anthracis by stable flies (Stomoxys calcitrans) and mosquitoes (Aedes aegypti and Aedes taeniorhynchus). Infect. Immun. 1987, 55, 1859–1861. [Google Scholar] [CrossRef] [Green Version]
- Traversa, D.; Otranto, D.; Iorio, R.; Carluccio, A.; Contri, A.; Paoletti, B.; Bartolini, R.; Giangaspero, A. Identification of the intermediate hosts of Habronema microstoma and Habronema muscae under field conditions. Med. Vet. Entomol. 2008, 22, 283–287. [Google Scholar] [CrossRef]
- Cook, D. A historical review of management options used against the stable fly (Diptera: Muscidae). Insects 2020, 11, 313. [Google Scholar] [CrossRef]
- Nattero, J.; Leonhard, G.; Gürtler, R.E.; Crocco, L.B. Evidence of selection on phenotypic plasticity and cost of plasticity in response to host-feeding sources in the major Chagas disease vector Triatoma infestans. Acta Trop. 2015, 152, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Dong, J.J.; Guan, D.L.; Xie, J.Y.; Xu, S.Q. Geographic variation in wing size and shape of the grasshopper Trilophidia annulata (Orthoptera: Oedipodidae): Morphological trait variations follow an ecogeographical rule. Sci. Rep. 2016, 6, 32680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dujardin, J.P. Morphometrics applied to medical entomology. Infect. Genet. Evol. 2008, 8, 875–890. [Google Scholar] [CrossRef]
- Alves, V.M.; Moura, M.O.; de Carvalho, C.J.B. Wing shape is influenced by environmental variability in Polietina orbitalis (Stein) (Diptera: Muscidae). Rev. Bras. Entomol. 2016, 60, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Oguz, G.; Kasap, O.E.; Alten, B. Wing morphology variations in a natural population of Phlebotomus tobbi Adler and Theodor 1930. J. Vector Ecol. 2017, 42, 223–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiphongpachara, T.; Juijayen, N.; Chansukh, K.K. Wing geometry analysis of Aedes aegypti (Diptera, Culicidae), a dengue virus vector, from multiple geographical locations of Samut Songkhram, Thailand. J. Arthropod-Borne Dis. 2018, 12, 351–360. [Google Scholar] [PubMed]
- Gilchrist, A.S.; Partridge, L. The contrasting genetic architecture of wing size and shape in Drosophila melanogaster. Heredity 2001, 86, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Dellicour, S.; Gerard, M.; Prunier, J.G.; Dewulf, A.; Kuhlmann, M.; Michez, D. Distribution and predictors of wing shape and size variability in three sister species of solitary bees. PLoS ONE 2017, 12, e0173109. [Google Scholar] [CrossRef]
- Barreaux, A.M.G.; Barreaux, P.; Thievent, K.; Koella, J.C. Larval environment influences vector competence of the malaria mosquito Anopheles gambiae. Malar. World J. 2016, 7, 1–6. [Google Scholar]
- Barreaux, A.M.G.; Stone, C.M.; Barreaux, P.; Koella, J.C. The relationship between size and longevity of the malaria vector Anopheles gambiae (s.s.) depends on the larval environment. Parasit. Vectors 2018, 11, 485. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, K.S.; Westby, K.M.; Medley, K.A. Genetic and environmental influences on the size-fecundity relationship in Aedes albopictus (Diptera: Culicidae): Impacts on population growth estimates? PLoS ONE 2018, 13, e0201465. [Google Scholar] [CrossRef] [PubMed]
- Baleba, S.B.S.; Masiga, D.; Torto, B.; Weldon, C.W.; Getahun, M.N. Effect of larval density and substrate quality on the wing geometry of Stomoxys calcitrans L. (Diptera: Muscidae). Parasit. Vectors 2019, 12, 222. [Google Scholar] [CrossRef] [PubMed]
- Baimai, V. Biodiversity in Thailand. J. R. Inst. Thail. 2010, 2, 107–114. [Google Scholar]
- Kermel-Torres, D. Atlas of Thailand: Spatial Structures and Development; IRD Editions: Paris, France, 2004; pp. 13–33. [Google Scholar]
- Muenworn, V.; Duvallet, G.; Thainchum, K.; Tuntakom, S.; Tanasilchayakul, S.; Prabaripai, A.; Akratanakul, P.; Sukonthabhirom, S.; Chareonviriyaphap, T. Geographic distribution of stomoxyine flies (Diptera: Muscidae) and diurnal activity of Stomoxys calcitrans in Thailand. J. Med. Entomol. 2010, 47, 791–797. [Google Scholar] [CrossRef]
- Changbunjong, T.; Weluwanarak, T.; Ratanakorn, P.; Maneeon, P.; Ganpanakngan, M.; Apiwathnasorn, C.; Sungvornyothin, S.; Sriwichai, P.; Sumruayphol, S.; Ruangsittichai, J. Distribution and abundance of Stomoxyini flies (Diptera: Muscidae) in Thailand. Southeast Asian J. Trop. Med. Public Health 2012, 43, 1400–1410. [Google Scholar]
- Gibert, P.; Debat, V.; Ghalambor, C.K. Phenotypic plasticity, global change, and the speed of adaptive evolution. Curr. Opin. Insect Sci. 2019, 35, 34–40. [Google Scholar] [CrossRef]
- Dujardin, J.P.; Kaba, D.; Solano, P.; Dupraz, M.; McCoy, K.D.; Jaramillo-O, N. Outline-based morphometrics, an overlooked method in arthropod studies? Infect. Genet. Evol. 2014, 28, 704–714. [Google Scholar] [CrossRef]
- Dujardin, S.; Dujardin, J.P. Geometric morphometrics in the cloud. Infect. Genet. Evol. 2019, 70, 189–196. [Google Scholar] [CrossRef]
- Changbunjong, T.; Sumruayphol, S.; Weluwanarak, T.; Ruangsittichai, J.; Dujardin, J.P. Landmark and outline-based geometric morphometrics analysis of three Stomoxys flies (Diptera: Muscidae). Folia Parasitol. 2016, 63, 37. [Google Scholar] [CrossRef] [Green Version]
- Changbunjong, T.; Prakaikowit, N.; Maneephan, P.; Kaewwiset, T.; Weluwanarak, T.; Chaiphongpachara, T.; Dujardin, J.P. Landmark data to distinguish and identify morphologically close Tabanus spp. (Diptera: Tabanidae). Insects 2021, 12, 974. [Google Scholar] [CrossRef]
- Morales Vargas, R.E.; Phumala-Morales, N.; Tsunoda, T.; Apiwathnasorn, C.; Dujardin, J.P. The phenetic structure of Aedes albopictus. Infect. Genet. Evol. 2013, 13, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Prudhomme, J.; Cassan, C.; Hide, M.; Toty, C.; Rahola, N.; Vergnes, B.; Dujardin, J.P.; Alten, B.; Sereno, D.; Bañuls, A.L. Ecology and morphological variations in wings of Phlebotomus ariasi (Diptera: Psychodidae) in the region of Roquedur (Gard, France): A geometric morphometrics approach. Parasit. Vectors 2016, 14, 578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Changbunjong, T.; Ruangsittichai, J.; Duvallet, G.; Pont, A.C. Molecular identification and geometric morphometric analysis of Haematobosca aberrans (Diptera: Muscidae). Insects 2020, 16, 451. [Google Scholar] [CrossRef] [PubMed]
- Mihok, S. The development of a multipurpose trap (the Nzi) for tsetse and other biting flies. Bull. Entomol. Res. 2002, 92, 385–403. [Google Scholar] [CrossRef]
- Tumrasvin, W.; Shinonaga, S. Studies on medically important flies in Thailand. V. On 32 species belonging to the subfamilies Muscinae and Stomoxyini including the taxonomic keys (Diptera: Muscidae). Bull. Tokyo Med. Dent. Univ. 1978, 25, 201–227. [Google Scholar]
- Arnqvist, G.; Martensson, T. Measurement error in geometric morphometrics: Empirical strategies to assess and reduce its impact on measures of shape. Acta Zool. Academ. Sci. Hung. 1998, 44, 73–96. [Google Scholar]
- Bookstein, F.L. Morphometric Tools for Landmark Data. Geometry and Biology; Cambridge University Press: Cambridge, UK, 1991. [Google Scholar]
- Sumruayphol, S.; Chittsamart, B.; Polseela, R.; Sriwichai, P.; Samung, Y.; Apiwathnasorn, C.; Dujardin, J.P. Wing geometry of Phlebotomus stantoni and Sergentomyia hodgsoni from different geographical locations in Thailand. C. R. Biol. 2017, 340, 37–46. [Google Scholar] [CrossRef]
- Demari-Silva, B.; Suesdek, L.; Sallum, M.A.M.; Marrelli, M.T. Wing geometry of Culex coronator (Diptera: Culicidae) from South and Southeast Brazil. Parasit. Vectors 2014, 7, 174. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Xu, Y.; Jiang, J.; Lavine, M.; Lavine, L.C. Host quality induces phenotypic plasticity in a wing polyphenic insect. Proc. Natl. Acad. Sci. USA 2018, 115, 7563–7568. [Google Scholar] [CrossRef] [Green Version]
- Johansson, F.; Söderquist, M.; Bokma, F. Insect wing shape evolution: Independent effects of migratory and mate guarding flight on dragonfly wings. Biol. J. Linn. Soc. 2009, 97, 362–372. [Google Scholar] [CrossRef] [Green Version]
- Chaiphongpachara, T.; Laojun, S. Wing morphometric variability of the malaria vector Anopheles (Cellia) epiroticus Linton et Harbach (Diptera: Culicidae) for the duration of the rainy season in coastal areas of Samut Songkhram, Thailand. Folia Parasitol. 2020, 67, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Morales Vargas, R.; Tsunoda, T.; Noda, J.; Bousses, P.; Nguyen, T.Y.; Hasebe, F.; Dujardin, J.P. Shape relatedness between geographic populations of Culex tritaeniorhynchus, the primary vector of Japanese encephalitis virus: A landmark study. Infect. Genet. Evol. 2021, 90, 104764. [Google Scholar] [CrossRef] [PubMed]
- Phanitchat, T.; Apiwathnasorn, C.; Sungvornyothin, S.; Samung, Y.; Dujardin, S.; Dujardin, J.P.; Sumruayphol, S. Geometric morphometric analysis of the effect of temperature on wing size and shape in Aedes albopictus. Med. Vet. Entomol. 2019, 33, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Marques, T.C.; Sallum, M.A.M.; Suesdek, L. Altitudinal population structure and microevolution of the malaria vector Anopheles cruzii (Diptera: Culicidae). Parasit. Vectors 2014, 7, 581. [Google Scholar] [CrossRef] [PubMed]
- Marks, D. Climate change and Thailand: Impact and response. Contemp. Southeast Asia 2011, 33, 229–258. [Google Scholar] [CrossRef]
- Virginio, F.; Oliveira Vidal, P.; Suesdek, L. Wing sexual dimorphism of pathogen-vector culicids. Parasit. Vectors 2015, 8, 159. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, C.; Almeida, F.; Almeida-Lopes, F.; Louise, C.; Pereira, S.N.; Petersen, V.; Vidal, P.O.; Virginio, F.; Suesdek, L. Geometric morphometrics in mosquitoes: What has been measured? Infect. Genet. Evol. 2017, 54, 205–215. [Google Scholar] [CrossRef]
- Gidaszewski, N.A.; Baylac, M.; Klingenberg, C.P. Evolution of sexual dimorphism of wing shape in the Drosophila melanogaster subgroup. BMC Evol. Biol. 2009, 9, 110. [Google Scholar] [CrossRef] [Green Version]
- Changbunjong, T.; Weluwanarak, T.; Samung, Y.; Ruangsittichai, J. Molecular identification and genetic variation of stomoxyine flies (Diptera: Muscidae) in Thailand based on cox1 barcode. J. Asia Pac. Entomol. 2016, 19, 1117–1123. [Google Scholar] [CrossRef]
- Tainchum, K.; Sukonthabhirom, S.; Duvallet, G.; Akratanakul, P.; Muenworn, V.; Chareonviriyaphap, T. Population structure of Stomoxys calcitrans (Diptera: Muscidae) from nine regions of Thailand. J. Econ. Entomol. 2010, 103, 1012–1018. [Google Scholar] [CrossRef]
- Long, A.; Mullaney, S.; Reid, L.; Fry, J.; Langley, C.; Mackay, T.F.C. High resolution mapping of genetic factors affecting abdominal bristle number in Drosophila melanogaster. Genetics 1995, 139, 1273–1291. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
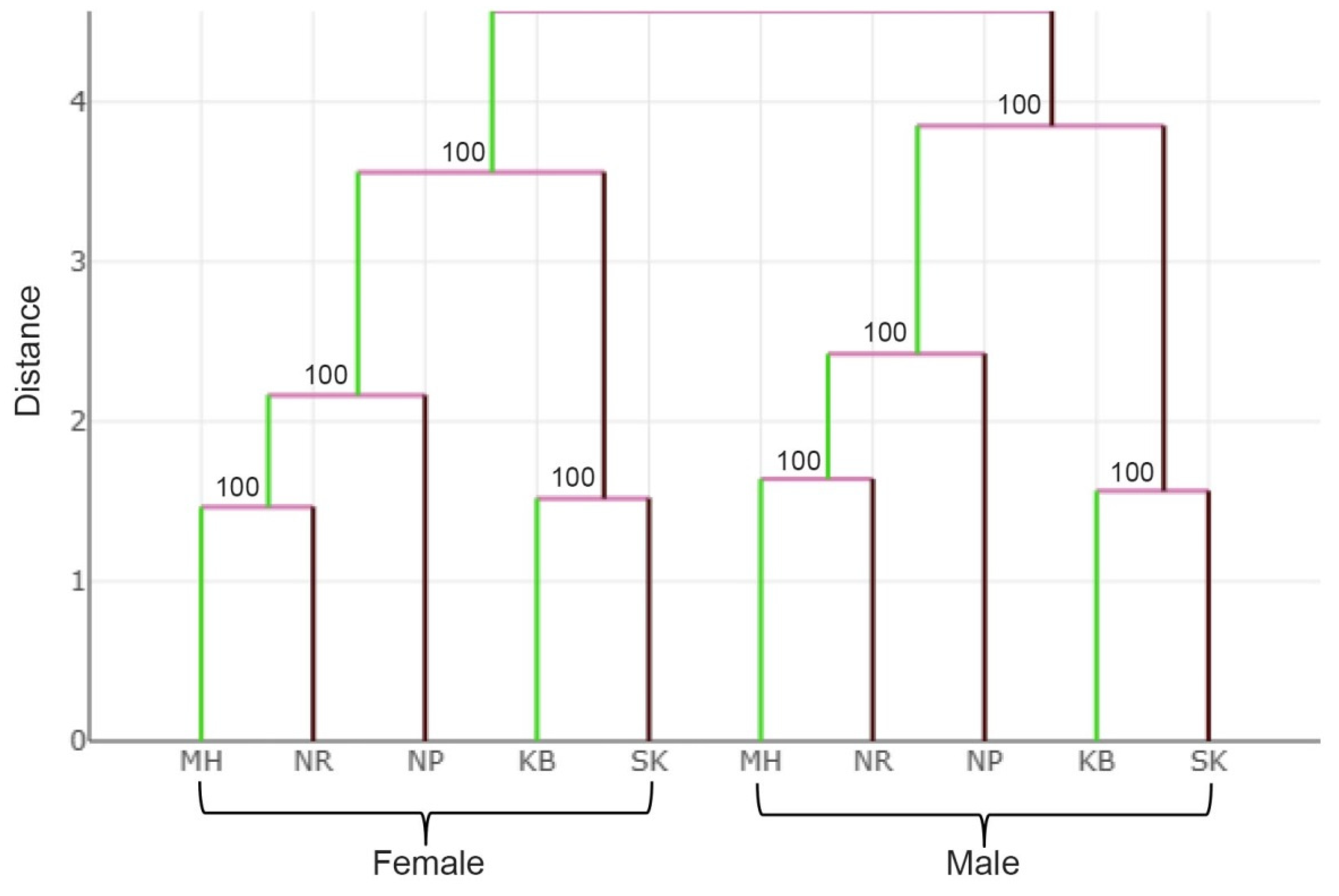{kind=link}
| Population | Code | Region | Date | Biotope | Altitude | Coordinates (Lat/Long) | n |
|---|---|---|---|---|---|---|---|
| Mae Hong Son | MH | Northern | July | Beef cattle farm located in rural area | 452 | 19°31′45″ N, 98°04′48″ E | Female 50, Male 50 |
| Nakhon Ratchasima | NR | Northeastern | February | Beef cattle and buffalo farm located in rural area | 498 | 14°22′23″ N, 101°44′51″ E | Female 45, Male 45 |
| Nakhon Pathom | NP | Central | May | Beef cattle farm located in urban area | 11 | 14°01′10″ N, 99°57′37″ E | Female 50, Male 50 |
| Kanchanaburi | KB | Western | March | Beef cattle farm located in rural area | 174 | 14°39′28″ N, 98°32′19″ E | Female 50, Male 50 |
| Songkhla | SK | Southern | February | Zoo located in urban area | 112 | 07°08′26″ N, 100°36′20″ E | Female 50, Male 50 |
| Population | n | Mean (mm) | (Min–Max) | Variance | SD | SE |
|---|---|---|---|---|---|---|
| Female | ||||||
| MH | 50 | 4.43 a | 3.88−4.84 | 0.06 | 0.25 | 0.04 |
| NR | 45 | 4.40 ac | 3.90−4.79 | 0.05 | 0.24 | 0.04 |
| NP | 50 | 4.40 ad | 3.87−4.73 | 0.03 | 0.18 | 0.02 |
| KB | 50 | 4.34 ab | 3.77−4.69 | 0.07 | 0.26 | 0.04 |
| SK | 50 | 4.09 h | 3.37−4.55 | 0.08 | 0.28 | 0.04 |
| Male | ||||||
| MH | 50 | 4.29 be | 3.71−4.63 | 0.05 | 0.23 | 0.03 |
| NR | 45 | 4.23 efg | 3.55−4.61 | 0.05 | 0.22 | 0.03 |
| NP | 50 | 4.30 bcdf | 3.75−4.59 | 0.04 | 0.20 | 0.03 |
| KB | 50 | 4.14 gh | 3.57−4.59 | 0.05 | 0.21 | 0.03 |
| SK | 50 | 3.97 i | 3.57−4.31 | 0.03 | 0.18 | 0.03 |
| Population | MH | NR | NP | KB | SK |
|---|---|---|---|---|---|
| Female | |||||
| MH | - | 0.005 | <0.001 | <0.001 | <0.001 |
| NR | 1.55 | - | <0.001 | <0.001 | <0.001 |
| NP | 1.94 | 2.51 | - | <0.001 | <0.001 |
| KB | 3.22 | 2.96 | 3.69 | - | <0.001 |
| SK | 3.45 | 2.99 | 3.98 | 1.41 | - |
| Male | |||||
| MH | - | <0.001 | <0.001 | <0.001 | <0.001 |
| NR | 1.68 | - | <0.001 | <0.001 | <0.001 |
| NP | 2.52 | 2.26 | - | <0.001 | <0.001 |
| KB | 4.38 | 3.99 | 4.08 | - | <0.001 |
| SK | 4.69 | 4.35 | 4.17 | 1.61 | - |
| Population | Size | Shape | ||
|---|---|---|---|---|
| % Correctly Assigned Individuals | No. of Correctly Assigned Individuals/Total Numbers | % Correctly Assigned Individuals | No. of Correctly Assigned Individuals/Total Numbers | |
| Female | ||||
| MH | 58 | 29/50 | 52 | 26/50 |
| NR | 0 | 0/45 | 51.11 | 23/45 |
| NP | 2 | 1/50 | 70 | 35/50 |
| KB | 6 | 3/50 | 66 | 33/50 |
| SK | 16 | 8/50 | 54 | 27/50 |
| Male | ||||
| MH | 12 | 6/50 | 58 | 29/50 |
| NR | 13.33 | 6/45 | 53.33 | 24/45 |
| NP | 4 | 2/50 | 72 | 36/50 |
| KB | 18 | 9/50 | 58 | 29/50 |
| SK | 52 | 26/50 | 80 | 40/50 |
| Total | 18.37 | 90/490 | 61.63 | 302/490 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaiphongpachara, T.; Duvallet, G.; Changbunjong, T. Wing Phenotypic Variation among Stomoxys calcitrans (Diptera: Muscidae) Populations in Thailand. Insects 2022, 13, 405. https://doi.org/10.3390/insects13050405
Chaiphongpachara T, Duvallet G, Changbunjong T. Wing Phenotypic Variation among Stomoxys calcitrans (Diptera: Muscidae) Populations in Thailand. Insects. 2022; 13(5):405. https://doi.org/10.3390/insects13050405
Chicago/Turabian StyleChaiphongpachara, Tanawat, Gerard Duvallet, and Tanasak Changbunjong. 2022. "Wing Phenotypic Variation among Stomoxys calcitrans (Diptera: Muscidae) Populations in Thailand" Insects 13, no. 5: 405. https://doi.org/10.3390/insects13050405