
Cross-Latitude Behavioural Axis in an Adult Damselfly Calopteryx splendens (Harris, 1780)



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Experimental Setup
2.3. Statistical Analyses
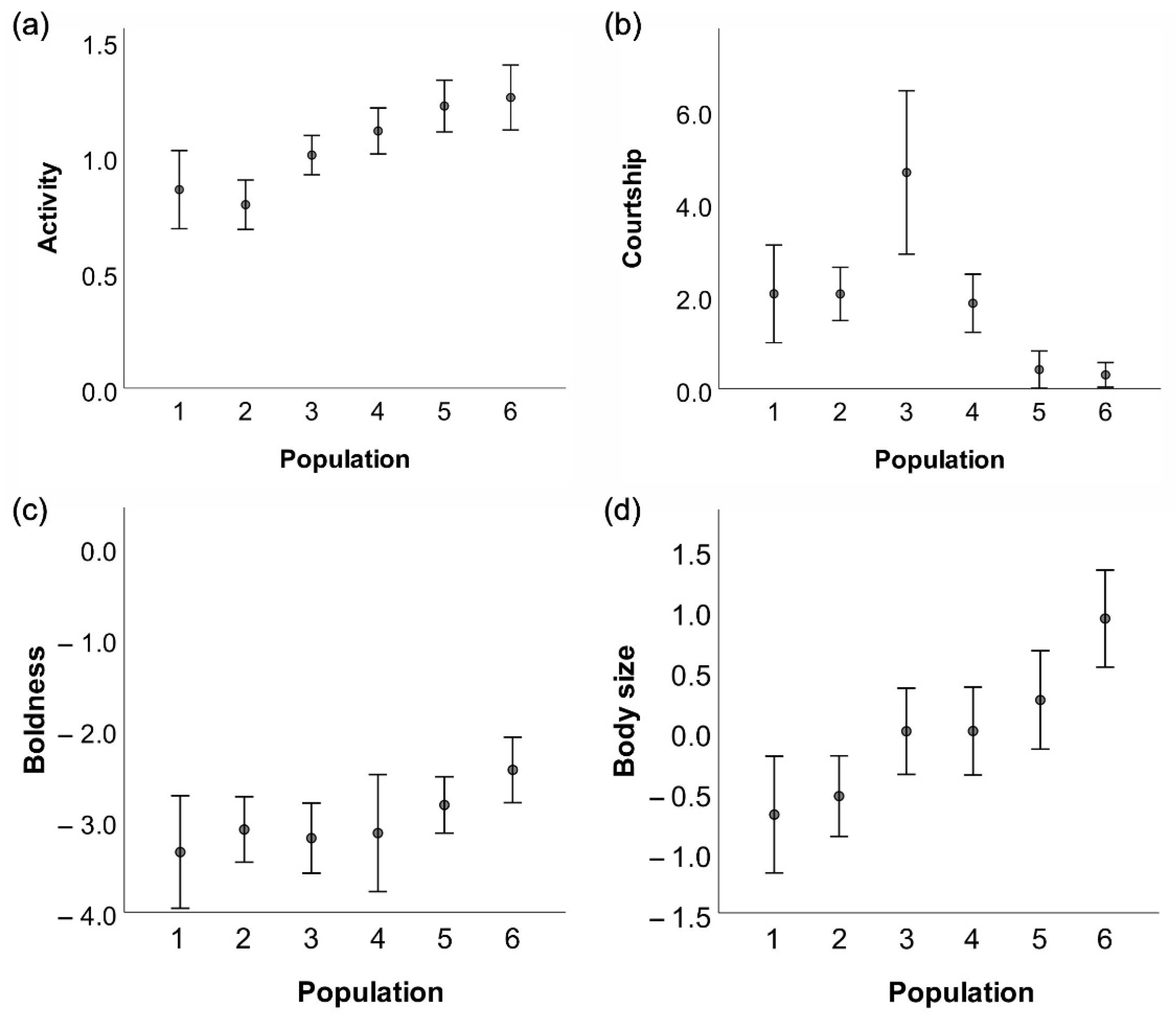
3. Results
3.1. Descriptive Statistics
3.2. Behavioural Syndromes
3.3. Body Size
3.4. Behaviours
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Breed, M.; Moore, J. Animal Behavior; Academic Press: Burlington, MA, USA, 2011; ISBN 978-0-08-091992-8. [Google Scholar]
- Puniamoorthy, N.; Ismail, M.R.B.; Tan, D.S.H.; Meier, R. From Kissing to Belly Stridulation: Comparative Analysis Reveals Surprising Diversity, Rapid Evolution, and Much Homoplasy in the Mating Behaviour of 27 Species of Sepsid Flies (Diptera: Sepsidae). J. Evol. Biol. 2009, 22, 2146–2156. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.J.; Brodie, E.D.; Wolf, J.B. Interacting Phenotypes and the Evolutionary Process: I. Direct and Indirect Genetic Effects of Social Interactions. Evolution 1997, 51, 1352–1362. [Google Scholar] [CrossRef]
- Candolin, U.; Wong, B.B.M. Behavioural Responses to a Changing World: Mechanisms and Consequences; Oxford University Press: Oxford, UK, 2012; ISBN 978-0-19-960257-5. [Google Scholar]
- Bae, M.-J.; Park, Y.-S. Biological Early Warning System Based on the Responses of Aquatic Organisms to Disturbances: A Review. Sci. Total Environ. 2014, 466–467, 635–649. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C.; Patton, J.L.; Conroy, C.J.; Parra, J.L.; White, G.C.; Beissinger, S.R. Impact of a Century of Climate Change on Small-Mammal Communities in Yosemite National Park, USA. Science 2008, 322, 261–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, P.; Lundrigan, B.; Hoffman, S.; Haraminac, A.; SETO, S. Climate-Induced Changes in Small Mammal Communities of the Northern Great Lakes Region. Glob. Change Biol. 2009, 15, 1434–1454. [Google Scholar] [CrossRef]
- Coulson, T.; Tuljapurkar, S. The Dynamics of a Quantitative Trait in an Age-Structured Population Living in a Variable Environment. Am. Nat. 2008, 172, 599–612. [Google Scholar] [CrossRef] [Green Version]
- Sih, A.; Bell, A.M.; Johnson, J.C.; Ziemba, R.E. Behavioral Syndromes: An Integrative Overview. Q. Rev. Biol. 2004, 79, 241–277. [Google Scholar] [CrossRef]
- Horváth, G.; Kriska, G. Polarization Vision in Aquatic Insects and Ecological Traps for Polarotactic Insects; CAB International Publishing: Wallingford, UK, 2008; pp. 204–229. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.J.; O’Connor, M.I.; Poloczanska, E.S.; Schoeman, D.S.; Buckley, L.B.; Burrows, M.T.; Duarte, C.M.; Halpern, B.S.; Pandolfi, J.M.; Parmesan, C.; et al. Ecological and Methodological Drivers of Species’ Distribution and Phenology Responses to Climate Change. Glob. Change Biol. 2016, 22, 1548–1560. [Google Scholar] [CrossRef] [Green Version]
- Musolin, D.L. Insects in a Warmer World: Ecological, Physiological and Life-History Responses of True Bugs (Heteroptera) to Climate Change. Glob. Change Biol. 2007, 13, 1565–1585. [Google Scholar] [CrossRef]
- Foster, S.A.; Endler, J.A. Geographic Variation in Behavior: Perspectives on Evolutionary Mechanisms: Perspectives on Evolutionary Mechanisms; Oxford University Press: New York, NY, USA, 1999; ISBN 978-0-19-535948-0. [Google Scholar]
- De Frenne, P.; Graae, B.J.; Rodríguez-Sánchez, F.; Kolb, A.; Chabrerie, O.; Decocq, G.; De Kort, H.; De Schrijver, A.; Diekmann, M.; Eriksson, O.; et al. Latitudinal Gradients as Natural Laboratories to Infer Species’ Responses to Temperature. J. Ecol. 2013, 101, 784–795. [Google Scholar] [CrossRef]
- Culumber, Z.W. Variation in Behavioral Traits across a Broad Latitudinal Gradient in a Livebearing Fish. Evol. Ecol. 2022, 36, 75–91. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Wright, J.; Kazem, A.J.N.; Thomas, D.K.; Hickling, R.; Dawnay, N. Behavioural Syndromes Differ Predictably between 12 Populations of Three-Spined Stickleback. J. Anim. Ecol. 2007, 76, 1128–1138. [Google Scholar] [CrossRef]
- Aspin, T.W.H.; Khamis, K.; Matthews, T.J.; Milner, A.M.; O’Callaghan, M.J.; Trimmer, M.; Woodward, G.; Ledger, M.E. Extreme Drought Pushes Stream Invertebrate Communities over Functional Thresholds. Glob. Change Biol. 2019, 25, 230–244. [Google Scholar] [CrossRef]
- Koski, S.E. Broader Horizons for Animal Personality Research. Front. Ecol. Evol. 2014, 2, 70. [Google Scholar] [CrossRef] [Green Version]
- Sih, A.; Bell, A.M.; Johnson, J.C. Behavioral Syndromes. In Evolutiornary Behavioural Ecology; Westneat, D., Fox, C., Eds.; Oxford University Press: New York, NY, USA, 2010; pp. 516–530. [Google Scholar]
- Myles-Gonzalez, E.; Burness, G.; Yavno, S.; Rooke, A.; Fox, M.G. To Boldly Go Where No Goby Has Gone before: Boldness, Dispersal Tendency, and Metabolism at the Invasion Front. Behav. Ecol. 2015, 26, 1083–1090. [Google Scholar] [CrossRef] [Green Version]
- Chapple, D.G.; Simmonds, S.M.; Wong, B.B.M. Can Behavioral and Personality Traits Influence the Success of Unintentional Species Introductions? Trends Ecol. Evol. 2012, 27, 57–64. [Google Scholar] [CrossRef]
- Monceau, K.; Moreau, J.; Poidatz, J.; Bonnard, O.; Thiéry, D. Behavioral Syndrome in a Native and an Invasive Hymenoptera Species. Insect Sci. 2015, 22, 541–548. [Google Scholar] [CrossRef]
- Stevens, V.M.; Whitmee, S.; Le Galliard, J.-F.; Clobert, J.; Böhning-Gaese, K.; Bonte, D.; Brändle, M.; Matthias Dehling, D.; Hof, C.; Trochet, A.; et al. A Comparative Analysis of Dispersal Syndromes in Terrestrial and Semi-Terrestrial Animals. Ecol. Lett. 2014, 17, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Bell, A.M. Insights for Behavioral Ecology from Behavioral Syndromes. In Advances in the Study of Behavior; Brockmann, H.J., Roper, T.J., Naguib, M., WynneEdwards, K.E., Barnard, C., Mitani, J.C., Eds.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2008; Volume 38, pp. 227–281. ISBN 978-0-12-004538-9. [Google Scholar]
- Way, G.P.; Kiesel, A.L.; Ruhl, N.; Snekser, J.L.; McRobert, S.P. Sex Differences in a Shoaling-Boldness Behavioral Syndrome, but No Link with Aggression. Behav. Processes 2015, 113, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Candolin, U.; Wong, B. Sexual Selection in Changing Environments: Consequences for Individuals and Populations. In Behavioural Responses to a Changing World; Oxford University Press: Oxford, UK, 2012; ISBN 978-0-19-960256-8. [Google Scholar]
- Denoël, M.; Doellen, J. Displaying in the Dark: Light-Dependent Alternative Mating Tactics in the Alpine Newt. Behav. Ecol. Sociobiol. 2010, 64, 1171–1177. [Google Scholar] [CrossRef]
- Fleisher, P. Evolution; Twenty-First Century Books: Minneapolis, MN, USA, 2006; ISBN 978-0-8225-2134-1. [Google Scholar]
- Dijkstra, K.; Schröter, A.; Lewington, R. Field Guide to the Dragonflies of Britain and Europe, 2nd ed.; Bloomsbury Publishing: London, UK, 2020; ISBN 978-1-4729-4399-6. [Google Scholar]
- Williams, C.M.; Henry, H.A.L.; Sinclair, B.J. Cold Truths: How Winter Drives Responses of Terrestrial Organisms to Climate Change. Biol. Rev. 2015, 90, 214–235. [Google Scholar] [CrossRef] [Green Version]
- Askew, R.R. Dragonflies of Europe, 2nd ed.; Harley Books: Essex, UK, 2004; ISBN 0-946589-75-5. [Google Scholar]
- Stettmer, C. Colonisation and Dispersal Patterns of Banded (Calopteryx splendens) and Beautiful Demoiselles (C-Virgo) (Odonata: Calopterygidae) in South-East German Streams. Eur. J. Entomol. 1996, 93, 579–593. [Google Scholar]
- Golab, M.J.; Śniegula, S.; Drobniak, S.M.; Zając, T.; Serrano-Meneses, M.A. Where Do Floaters Settle? An Experimental Approach in Odonates. Anim. Behav. 2013, 86, 1069–1075. [Google Scholar] [CrossRef]
- Golab, M.J.; Gołąb, P.A.; Contreras-Garduño, J.; Zając, T.; Sniegula, S. The Effects of Habitat Deterioration and Social Status on Patrolling Behavior in the Territorial Damselfly Calopteryx splendens. Pol. J. Ecol. 2017, 65, 122–131. [Google Scholar] [CrossRef]
- Zahner, R. Über Die Bindung Der Mitteleuropäischen Calopteryx-Arten (Odonata, Zygoptera) an Den Lebensraum Des Strömenden Wassers II. Der Anteil Der Imagines an Der Biotopbindung. Int. Rev. Der Gesamten Hydrobiol. Hydrogr. 1960, 45, 101–123. [Google Scholar] [CrossRef]
- Corbet, P. Dragonflies: Behaviour and Ecology of Odonata; Harley Books: Alpharetta, GA, USA, 2004; ISBN 978-0-946589-77-7. [Google Scholar]
- Rüppell, G.; Rehfeldt, G.; Schütte, C.; Hilfert-Rüppell, D. Die Prachtlibellen Europas: Gattung Calopteryx; Westarp Wissenschaften: Hohenwarsleben, Germany, 2005; ISBN 3-89432-883-5. [Google Scholar]
- Meek, S.B.; Herman, T.B. A Comparison of the Reproductive Behaviours of Three Calopteryx Species (Odonata: Calopterygidae) in Nova Scotia. Can. J. Zool. 1990, 68, 10–16. [Google Scholar] [CrossRef]
- Tynkkynen, K.; Rantala, M.; Suhonen, J. Interspecific Aggression and Character Displacement in the Damselfly Calopteryx splendens. J Evol. Biol 2004, 17, 759–767. [Google Scholar] [CrossRef]
- Golab, M.J.; Sniegula, S.; Antoł, A.; Brodin, T. Adult Insect Personality in the Wild—Calopteryx splendens as a Model for Field Studies. Ecol. Evol. 2021, 11, 18467–18476. [Google Scholar] [CrossRef] [PubMed]
- Réale, D.; Reader, S.; Sol, D.; McDougall, P.; Dingemanse, N. Integrating Animal Temperament within Ecology and Evolution. Biol. Rev. 2007, 82, 291–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dougherty, L.R.; Guillette, L.M. Linking Personality and Cognition: A Meta-Analysis. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Killen, S.S.; Marras, S.; Metcalfe, N.B.; McKenzie, D.J.; Domenici, P. Environmental Stressors Alter Relationships between Physiology and Behaviour. Trends Ecol. Evol. 2013, 28, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Brodin, T. Behavioral Syndrome over the Boundaries of Life Carryovers from Larvae to Adult Damselfly. Behav. Ecol. 2009, 20, 30–37. [Google Scholar] [CrossRef] [Green Version]
- IBM Corp, N. IBM SPSS Statistics for Windows; IBM Corp.: Armon, NY, USA, 2019. [Google Scholar]
- Golab, M.J.; Brodin, T.; Sniegula, S. Two Experimental Designs Generate Contrasting Patterns of Behavioral Differentiation along a Latitudinal Gradient in Lestes Sponsa—Common-Garden Not so Common after All? Ecol. Evol. 2020, 10, 10242–10253. [Google Scholar] [CrossRef] [PubMed]
- Mason, T.H.E.; Brivio, F.; Stephens, P.A.; Apollonio, M.; Grignolio, S. The Behavioral Trade-off between Thermoregulation and Foraging in a Heat-Sensitive Species. Behav. Ecol. 2017, 28, 908–918. [Google Scholar] [CrossRef] [Green Version]
- Choe, J.C.; Crespi, B.J. The Evolution of Mating Systems in Insects and Arachnids; Cambridge University Press: Cambridge, UK, 1997; ISBN 978-0-521-58976-5. [Google Scholar]
- Emlen, S.T.; Oring, L.W. Ecology, Sexual Selection, and the Evolution of Mating Systems. Science 1977, 197, 215–223. [Google Scholar] [CrossRef] [Green Version]
- Benton, T.G. Dispersal Ecology and Evolution; OUP Oxford: Oxford, UK, 2012; ISBN 978-0-19-960890-4. [Google Scholar]
- Hickling, R.; Roy, D.B. A Northward Shift of Range Margins in British Odonata. Glob. Change Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- Atwell, J.W.; Cardoso, G.C.; Whittaker, D.J.; Campbell-Nelson, S.; Robertson, K.W.; Ketterson, E.D. Boldness Behavior and Stress Physiology in a Novel Urban Environment Suggest Rapid Correlated Evolutionary Adaptation. Behav. Ecol. 2012, 23, 960–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golab, M.; Johansson, F.; Sniegula, S. Let’s Mate Here and Now–Seasonal Constraints Increase Mating Efficiency. Ecol. Entomol. 2019, 44, 623–629. [Google Scholar] [CrossRef]
- Godin, J.G.J.; Dugatkin, L.A. Female Mating Preference for Bold Males in the Guppy, Poecilia reticulata. Proc. Natl. Acad. Sci. USA 1996, 93, 10262–10267. [Google Scholar] [CrossRef] [Green Version]
- Schuett, W.; Tregenza, T.; Dall, S.R.X. Sexual Selection and Animal Personality. Biol. Rev. 2010, 85, 217–246. [Google Scholar] [CrossRef]
- Bell, A.M.; Sih, A. Exposure to Predation Generates Personality in Threespined Sticklebacks (Gasterosteus aculeatus). Ecol. Lett. 2007, 10, 828–834. [Google Scholar] [CrossRef] [PubMed]
- Huntingford, F.A. The Relationship between Anti-Predator Behaviour and Aggression among Conspecifics in the Three-Spined Stickleback, Gasterosteus aculeatus. Anim. Behav. 1976, 24, 245–260. [Google Scholar] [CrossRef]
- Schuett, W.; Godin, J.-G.J.; Dall, S.R.X. Do Female Zebra Finches, Taeniopygia guttata, Choose Their Mates Based on Their “Personality”? Ethology 2011, 117, 908–917. [Google Scholar] [CrossRef]
- Roslin, T.; Hardwick, B.; Novotny, V.; Petry, W.K.; Andrew, N.R.; Asmus, A.; Barrio, I.C.; Basset, Y.; Boesing, A.L.; Bonebrake, T.C.; et al. Higher Predation Risk for Insect Prey at Low Latitudes and Elevations. Science 2017, 356, 742–744. [Google Scholar] [CrossRef] [Green Version]
- Goodchild, C.G.; Schmidt, L.M.; DuRant, S.E. Evidence for the ‘Behavioural Character’ Hypothesis: Does Boldness Programme Disparate Antipredator Strategies? Anim. Behav. 2020, 164, 123–132. [Google Scholar] [CrossRef]
- Schmidt-Kloiber, A.; Hering, D. Www.Freshwaterecology.Info—An Online Tool That Unifies, Standardises and Codifies More than 20,000 European Freshwater Organisms and Their Ecological Preferences. Ecol. Indic. 2015, 53, 271–282. [Google Scholar] [CrossRef]
- Gallesi, M.M.; Sacchi, R. Voltinism and Larval Population Structure of Calopteryx splendens (Odonata: Calopterygidae) in the Po Valley. Int. J. Odonatol. 2019, 22, 21–30. [Google Scholar] [CrossRef]
- Corbet, P.; Suhling, F.; Soendgerath, D. Voltinism of Odonata: A Review. Int. J. Odonatol. 2006, 9, 1–44. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Jackson, J.K.; Funk, D.H. Semivoltinism, Seasonal Emergence, and Adult Size Variation in a Tropical Stream Mayfly (Euthyplocia hecuba). J. North Am. Benthol. Soc. 1995, 14, 131–146. [Google Scholar] [CrossRef]
- Purse, B.V.; Thompson, D.J. Voltinism and Larval Growth Pattern in Coenagrion mercuriale (Odonata: Coenagrionidae) at Its Northern Range Margin. EJE 2013, 99, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Raczyński, M.; Stoks, R.; Johansson, F.; Sniegula, S. Size-Mediated Priority Effects Are Trait-Dependent and Consistent across Latitudes in a Damselfly. Oikos 2021, 130, 1535–1547. [Google Scholar] [CrossRef]
- Laakso, L.K.; Ilvonen, J.J.; Suhonen, J. Phenotypic Variation in Male Calopteryx splendens Damselflies: The Role of Wing Pigmentation and Body Size in Thermoregulation. Biol. J. Linn. Soc. 2021, 134, 685–696. [Google Scholar] [CrossRef]
- May, M. Thermoregulation and Adaptation to Temperature in Dragonflies (Odonata: Anisoptera). Ecol. Monogr. 1976, 46, 1–32. [Google Scholar] [CrossRef]
- Peters, R.H.; Peters, R.H. The Ecological Implications of Body Size; Cambridge University Press: Cambridge, UK, 1986; ISBN 978-0-521-28886-6. [Google Scholar]
- Kleiber, M. Body Size and Metabolic Rate. Physiol. Rev. 1947, 27, 511–541. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
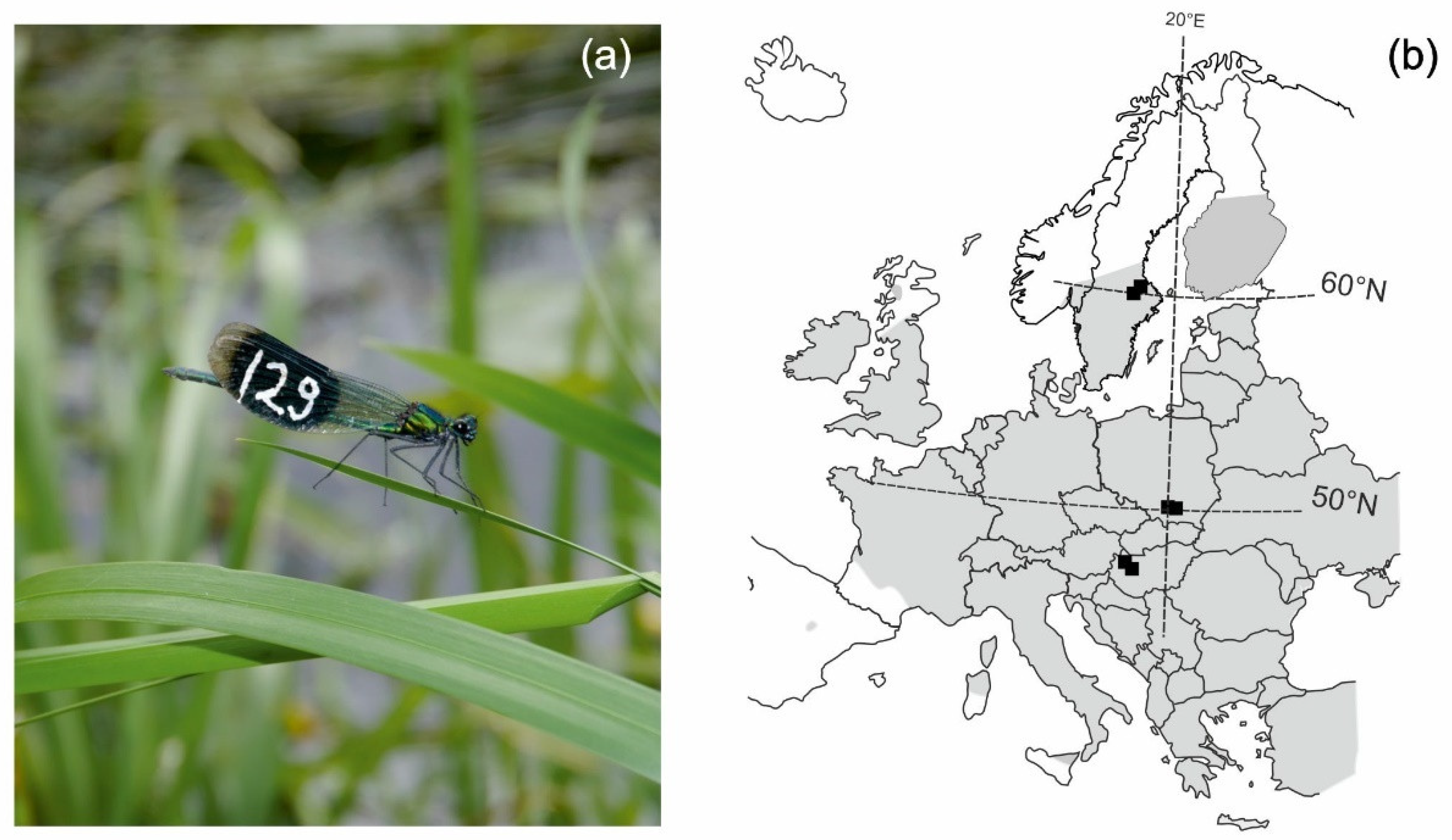
| Region | Population | Coordinates | Sampling Dates | Sample Size |
|---|---|---|---|---|
| Southern | 1 | 47.59087° N 16.75141° E | 27–29 July | 23 males |
| 2 | 47.573524° N 17.000544° E | 31 July–3 August | 23 males | |
| Central | 3 | 50.0769° N 19.85464° E | 21–23 July | 21 males |
| 4 | 50.0805° N 20.62278° E | 8–11 July | 19 males | |
| Northern | 5 | 59.71278° N 16.65955° E | 29–30 June | 23 males |
| 6 | 59.91231° N 16.87902° E | 26–28 June | 32 males |
| Behavioural Axis | Trait Measured | Description |
|---|---|---|
| Activity | Patrol flights | Number of patrol flights performed during observation period |
| Male chases | Number of chases of intruder males approaching or passing a territory defended by a focal territorial male | |
| Courtships | Courtship | Number of courtship displays performed during 20 min observation period |
| Boldness | Immobility | Total time [s] of the territorial male remaining motionless after the predatory attack simulation. The motionless periods between consequtive movements toward the territory are summed up |
| First move | Time [s] until the territorial male first moved after the motionless behaviour | |
| Time to return | Time [s] passed until the territorial male returned to his territory after the predatory attack. The maximum time the observer waited for the male to come back was 180 s |
| Trait | Pairwise Contrast | Estimate | t | df | p |
|---|---|---|---|---|---|
| Activity | S–C | −0.18 | −3.59 | 131 | <0.001 |
| S–N | −0.34 | −7.07 | 131 | <0.001 | |
| C–N | −0.16 | −3.19 | 131 | 0.002 | |
| Courtship | S–C | −0.96 | −0.89 | 131 | 0.372 |
| S–N | 1.69 | 2.74 | 131 | 0.007 | |
| C–N | 0.96 | 2.98 | 131 | 0.003 | |
| Boldness | S–C | 0.05 | 0.22 | 121 | 0.823 |
| S–N | 0.57 | 2.81 | 121 | 0.006 | |
| C–N | 0.52 | 2.49 | 121 | 0.014 | |
| Body size | S–C | −0.54 | −2.12 | 120 | 0.031 |
| S–N | −1.07 | −4.43 | 120 | <0.001 | |
| C–N | −0.54 | −2.17 | 120 | 0.032 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Golab, M.J.; Sniegula, S.; Brodin, T. Cross-Latitude Behavioural Axis in an Adult Damselfly Calopteryx splendens (Harris, 1780). Insects 2022, 13, 342. https://doi.org/10.3390/insects13040342
Golab MJ, Sniegula S, Brodin T. Cross-Latitude Behavioural Axis in an Adult Damselfly Calopteryx splendens (Harris, 1780). Insects. 2022; 13(4):342. https://doi.org/10.3390/insects13040342
Chicago/Turabian StyleGolab, Maria J., Szymon Sniegula, and Tomas Brodin. 2022. "Cross-Latitude Behavioural Axis in an Adult Damselfly Calopteryx splendens (Harris, 1780)" Insects 13, no. 4: 342. https://doi.org/10.3390/insects13040342