
CCHamide-2 Signaling Regulates Food Intake and Metabolism in Gryllus bimaculatus


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Tissue-Distribution Analyses Using Reverse Transcription-PCR (RT-PCR)
2.3. RNA Interference
2.4. Chemical Synthesis of GbCCHa-2
2.5. Food Intake Assays
2.6. Measurement of Lipid and Carbohydrate Concentrations
2.7. Food Choice Assay
3. Results
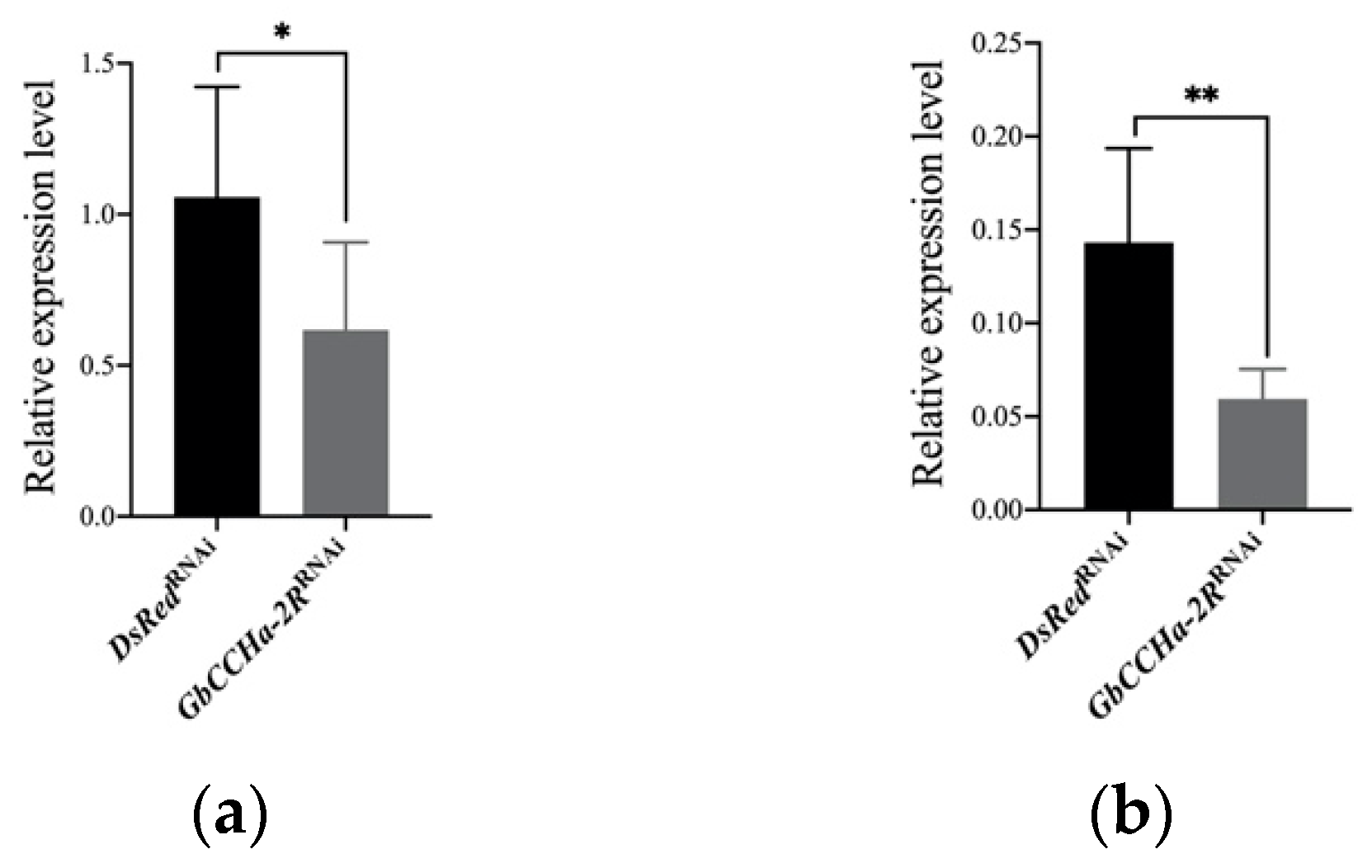
3.1. Tissue Distributions of GbCCHa-2 and GbCCHa-2R
3.2. Effects of GbCCHa-2 Signaling on Food Intake and Body Weight
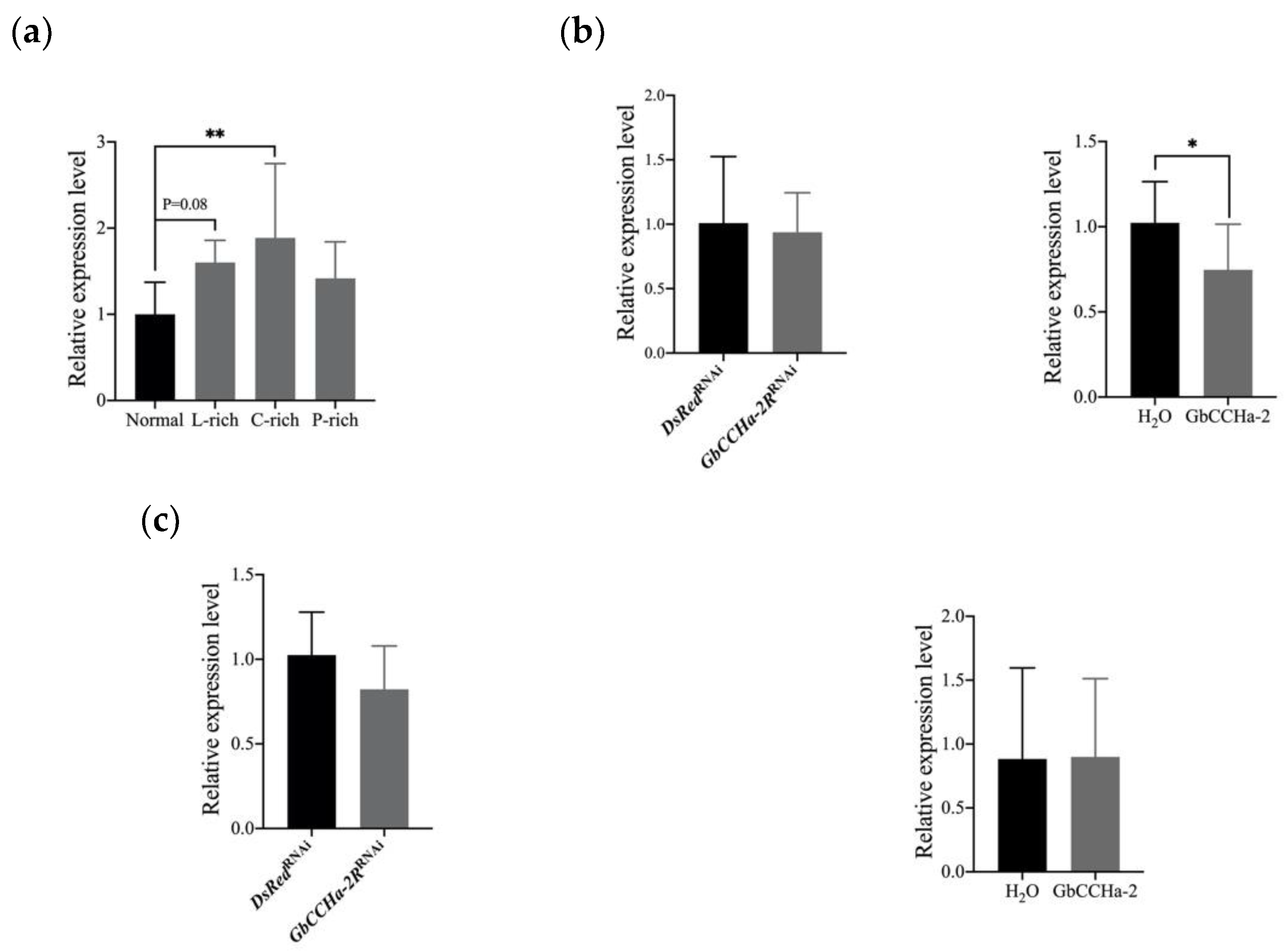
3.3. Effects of GbCCHa-2 Signaling on Lipid and Carbohydrate Concentrations in the Hemolymph and Fat Body
3.4. Effects of Dietary Nutrients on the Transcript Level of GbCCHa-2R
3.5. Effects of GbCCHa-2 Signaling on Food Preference
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Waterson, M.J.; Horvath, T.L. Neuronal regulation of energy homeostasis: Beyond the hypothalamus and feeding. Cell Metab. 2015, 22, 962–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unniappan, S.; Orchard, I.; Delgado, M.J. Neuroendocrine control of energy homeostasis in non-mammalian vertebrates and invertebrates. Front. Endocrinol. 2020, 11, 404. [Google Scholar] [CrossRef] [PubMed]
- Després, J.-P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Schoofs, L.; De Loof, A.; Van Hiel, M.B. Neuropeptides as regulators of behavior in insects. Annu. Rev. Entomol. 2017, 62, 35–52. [Google Scholar] [CrossRef] [PubMed]
- Woods, S.C.; Rushing, P.A.; Seeley, R.J. Neuropeptides and the control of energy homeostasis. In Nutrition and Brain; Nestlé Nutrition Workshop Series: Clinical & Performance Programme; Karger Medical and Scientific Publishers: Basel, Switzerland, 2001; Volume 5, pp. 93–112. [Google Scholar] [CrossRef]
- Taguchi, A.; White, M.F. Insulin-like signaling, nutrient homeostasis, and life span. Annu. Rev. Physiol. 2008, 70, 191–212. [Google Scholar] [CrossRef] [PubMed]
- Morton, G.; Cummings, D.; Baskin, D.; Barsh, G.; Schwartz, M. Central nervous system control of food intake and body weight. Nature 2006, 443, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, A.M.; Torres-Alemán, I. The many faces of insulin-like peptide signalling in the brain. Nat. Rev. Neurosci. 2012, 13, 225–239. [Google Scholar] [CrossRef]
- Chung, B.Y.; Ro, J.; Hutter, S.A.; Miller, K.M.; Guduguntla, L.S.; Kondo, S.; Pletcher, S.D. Drosophila neuropeptide F signaling independently regulates feeding and sleep-wake behavior. Cell Rep. 2017, 19, 2441–2450. [Google Scholar] [CrossRef] [Green Version]
- Fukumura, K.; Konuma, T.; Tsukamoto, Y.; Nagata, S. Adipokinetic hormone signaling determines dietary fatty acid preference through maintenance of hemolymph fatty acid composition in the cricket Gryllus bimaculatus. Sci. Rep. 2018, 8, 4737. [Google Scholar] [CrossRef]
- Gáliková, M.; Diesner, M.; Klepsatel, P.; Hehlert, P.; Xu, Y.; Bickmeyer, I.; Predel, R.; Kühnlein, R.P. Energy homeostasis control in Drosophila adipokinetic hormone mutants. Genetics 2015, 201, 665–683. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Zhang, X.; Chen, X.; Li, Y.; Li, W.; Cheng, Y.; Zhou, J.; You, K.; Zhou, Q. Adipokinetic hormone receptor mediates lipid mobilization to regulate starvation resistance in the brown planthopper, Nilaparvata lugens. Front. Physiol. 2018, 9, 1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zdárek, J.; Nachman, R.; Denlinger, D. Parturition hormone in the tsetse Glossina morsitans: Activity in reproductive tissues from other species and response of tsetse to identified neuropeptides and other neuroactive compounds. J. Insect Physiol. 2000, 46, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Thiel, D.; Bauknecht, P.; Jékely, G.; Hejnol, A. A nemertean excitatory peptide/CCHamide regulates ciliary swimming in the larvae of Lineus longissimus. Front. Zool. 2019, 16, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, K.K.; Hauser, F.; Williamson, M.; Weber, S.B.; Grimmelikhuijzen, C.J. The Drosophila genes CG14593 and CG30106 code for G-protein-coupled receptors specifically activated by the neuropeptides CCHamide-1 and CCHamide-2. Biochem. Biophys. Res. Commun. 2011, 404, 184–189. [Google Scholar] [CrossRef]
- Mains, R.E.; Eipper, B.A. Neuropeptide Receptors. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects, 6th ed.; Siegel, G.J., Agranoff, B.W., Albers, R.W., Fisher, S.K., Uhler, M.D., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1999. Available online: https://www.ncbi.nlm.nih.gov/books/NBK27941/ (accessed on 21 February 2022).
- Tian, Y.; Jiang, C.; Pan, Y.; Guo, Z.; Wang, W.; Luo, X.; Cao, Z.; Zhang, B.; Yang, J.; Shi, Y.; et al. Bombyx neuropeptide G protein-coupled receptor A14 and A15 are two functional G protein-coupled receptors for CCHamide neuropeptides. Insect Biochem. Mol. Biol. 2021, 131, 103553. [Google Scholar] [CrossRef]
- Farhan, A.; Gulati, J.; Groβe-Wilde, E.; Vogel, H.; Hansson, B.S.; Knaden, M. The CCHamide 1 receptor modulates sensory perception and olfactory behavior in starved Drosophila. Sci. Rep. 2013, 3, 2765. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, Y.; Hermann-Luibl, C.; Katsura, M.; Sekiguchi, M.; Ida, T.; Helfrich-Förster, C.; Yoshii, T. The CCHamide1 neuropeptide expressed in the anterior dorsal neuron 1 conveys a circadian signal to the ventral lateral neurons in Drosophila melanogaster. Front. Physiol. 2018, 9, 1276. [Google Scholar] [CrossRef] [Green Version]
- Rogulja, D.; Titos, I. A gut-secreted peptide controls arousability through modulation of dopaminergic neurons in the brain. BioRxiv 2020. [Google Scholar] [CrossRef]
- Ren, G.R.; Hauser, F.; Rewitz, K.F.; Kondo, S.; Engelbrecht, A.F.; Didriksen, A.K.; Schjøtt, S.R.; Sembach, F.E.; Li, S.; Søgaard, K.C.; et al. CCHamide-2 is an orexigenic brain-gut peptide in Drosophila. PLoS ONE 2015, 10, e0133017. [Google Scholar] [CrossRef] [Green Version]
- Ida, T.; Takahashi, T.; Tominaga, H.; Sato, T.; Sano, H.; Kume, K.; Ozaki, M.; Hiraguchi, T.; Shiotani, H.; Terajima, S.; et al. Isolation of the bioactive peptides CCHamide-1 and CCHamide-2 from Drosophila and their putative role in appetite regulation as ligands for G protein-coupled receptors. Front. Endocrinol. 2012, 3, 177. [Google Scholar] [CrossRef] [Green Version]
- Sano, H.; Nakamura, A.; Texada, M.J.; Truman, J.W.; Ishimoto, H.; Kamikouchi, A.; Nibu, Y.; Kume, K.; Ida, T.; Kojima, M. The nutrient-responsive hormone CCHamide-2 controls growth by regulating insulin-like peptides in the brain of Drosophila melanogaster. PLoS Genet. 2015, 11, e1005209. [Google Scholar] [CrossRef] [Green Version]
- Millington, J.W.; Chao, C.; Sun, Z.; Basner-Collins, P.J.; Brownrigg, G.P.; Wat, L.W.; Hudry, B.; Miguel-Aliaga, I.; Rideout, E.J. Female-specific upregulation of insulin pathway activity mediates the sex difference in Drosophila body size plasticity. BioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Capriotti, N.; Ianowski, J.P.; Gioino, P.; Ons, S. The neuropeptide CCHamide2 regulates diuresis in the Chagas disease vector Rhodnius prolixus. J. Exp. Biol. 2019, 222, jeb203000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capriotti, N.; Gioino, P.; Ons, S.; Ianowski, J.P. The neuropeptide RhoprCCHamide2 inhibits serotonin-stimulated transcellular Na+ transport across the anterior midgut of the vector of Chagas disease, Rhodnius prolixus. J. Exp. Biol. 2021, 224, jeb242272. [Google Scholar] [CrossRef] [PubMed]
- Shahid, S.; Shi, Y.; Yang, C.; Li, J.; Ali, M.Y.; Smagghe, G.; Liu, T.X. CCHamide2-receptor regulates feeding behavior in the pea aphid, Acyrthosiphon pisum. Peptides 2021, 14, 170596. [Google Scholar] [CrossRef]
- Fukumura, K.; Nagata, S. Behavioral tracing demonstrates dietary nutrient discrimination in two-spotted crickets Gryllus bimaculatus. Biosci. Biotechnol. Biochem. 2017, 81, 1990–1993. [Google Scholar] [CrossRef] [Green Version]
- Mito, T.; Noji, S. The two-spotted cricket Gryllus bimaculatus: An emerging model for developmental and regeneration studies. Cold Spring Harb. Protoc. 2008, 144, 27–35. [Google Scholar] [CrossRef]
- Zhou, Y.J.; Fukumura, K.; Nagata, S. Effects of adipokinetic hormone and its related peptide on maintaining hemolymph carbohydrate and lipid levels in the two-spotted cricket, Gryllus bimaculatus. Biosci. Biotechnol. Biochem. 2018, 82, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Nagata, S.; Nagasawa, H. Calcitonin-like peptide hormone (CT/DH) in the frontal ganglia as a feeding regulatory peptide of the silkworm, Bombyx mori. Peptides 2017, 98, 23–28. [Google Scholar] [CrossRef]
- Lorenz, M.W. Adipokinetic hormone inhibits the formation of energy stores and egg production in the cricket Gryllus bimaculatus. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 136, 197–206. [Google Scholar] [CrossRef]
- Raubenheimer, D.; Simpson, S.J. Integrative models of nutrient balancing: Application to insects and vertebrates. Nutr. Res. Rev. 1997, 10, 151–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Torre-Muruzabal, T.; Søgaard, K.C.; Ren, G.R.; Hauser, F.; Engelsen, S.M.; Pødenphanth, M.D.; Desjardins, A.; Grimmelikhuijzen, C.J. Expression patterns of the Drosophila neuropeptide CCHamide-2 and its receptor may suggest hormonal signaling from the gut to the brain. PLoS ONE 2013, 8, e76131. [Google Scholar] [CrossRef] [PubMed]
- Christie, A.E. Prediction of Scylla olivacea (Crustacea; Brachyura) peptide hormones using publicly accessible transcriptome shotgun assembly (TSA) sequences. Gen. Comp. Endocrinol. 2016, 230–231, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagata, S.; Zhou, Y.J. Feeding-modulating neuropeptides and peptide hormones in insects. Adv. Insect Physiol. 2019, 7, 137–172. [Google Scholar] [CrossRef]
- Scriber, J.; Slansky, F., Jr. The nutritional ecology of immature insects. Annu. Rev. Entomol. 1981, 26, 183–211. [Google Scholar] [CrossRef]
- Lee, G.; Park, J.H. Hemolymph sugar homeostasis and starvation-induced hyperactivity affected by genetic manipulations of the adipokinetic hormone-encoding gene in Drosophila melanogaster. Genetics 2004, 167, 311–323. [Google Scholar] [CrossRef] [Green Version]
- Gruber, F.; Knapek, S.; Fujita, M.; Matsuo, K.; Bräcker, L.; Shinzato, N.; Siwanowicz, I.; Tanimura, T.; Tanimoto, H. Suppression of conditioned odor approach by feeding is independent of taste and nutritional value in Drosophila. Curr. Biol. 2013, 23, 507–514. [Google Scholar] [CrossRef] [Green Version]
- Konuma, T.; Tsukamoto, Y.; Nagasawa, H.; Nagata, S. Imbalanced hemolymph lipid levels affect feeding motivation in the two-spotted cricket, Gryllus bimaculatus. PLoS ONE 2016, 11, e0154841. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′-3′ |
|---|---|
| GbCCHa-2 RT-F | CAGGCAGTAGCAGCAGCA |
| GbCCHa-2 RT-R | GAAGCACGAGTGG CCGAA |
| GbCCHa-2R RT-F | CATGGAGGTGGACGTGGAG |
| GbCCHa-2R RT-R | ATGGGGTTGATGCACGAGTT |
| EF RT-F | ATGCCTGTATCTTGACTGCTCA |
| EF RT-R | ATGGTTTGCTTCCAGTTTCAGT |
| DsRed2 RT-F | AGAACGTCATCACCGAGTTCAT |
| DsRed2 RT-R | CCGATGAACTTCACCTTGTAGA |
| T7 promoter-F | CTTCTAATACGACTCACTATAG |
| T7 promoter-R | CTTCTAATACGACTCACTATAG |
| GbCCHa-2R q-F | AGCTGCTCACCTACATCGTG |
| GbCCHa-2R q-R | GAAGATGAACACCAGCGTGC |
| Gbilp q-F | CTGAGAAAGAGCCAGAGCC |
| Gbilp q-R | ATTGCACATGACTTCCGAGA |
| GbAKH q-F | CCCACAGTGCACAGGATCAT |
| GbAKH q-R | CGCCAAAACCAGAACCAAGG |
| ß-actin q-F | TTGACAATGGATCCGGAATGT |
| ß-actin q-R | AAAACTGCCCTGGGTGCAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Z.; Tsuchimoto, M.; Nagata, S. CCHamide-2 Signaling Regulates Food Intake and Metabolism in Gryllus bimaculatus. Insects 2022, 13, 324. https://doi.org/10.3390/insects13040324
Zhu Z, Tsuchimoto M, Nagata S. CCHamide-2 Signaling Regulates Food Intake and Metabolism in Gryllus bimaculatus. Insects. 2022; 13(4):324. https://doi.org/10.3390/insects13040324
Chicago/Turabian StyleZhu, Zhen, Maho Tsuchimoto, and Shinji Nagata. 2022. "CCHamide-2 Signaling Regulates Food Intake and Metabolism in Gryllus bimaculatus" Insects 13, no. 4: 324. https://doi.org/10.3390/insects13040324