
Association of Fig Pollinating Wasps and Fig Nematodes inside Male and Female Figs of a Dioecious Fig Tree in Sumatra, Indonesia

 , ,
, , 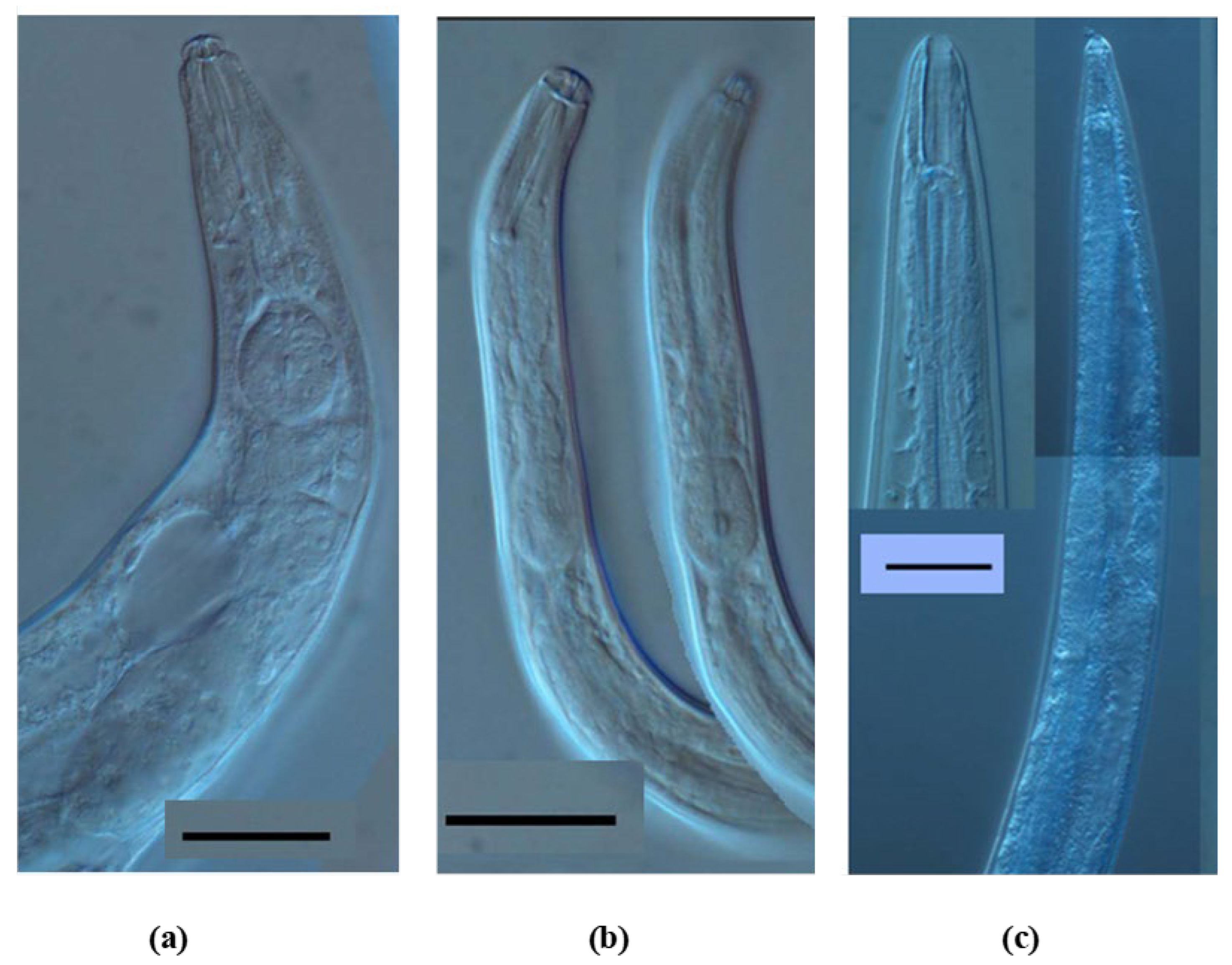{kind=link}
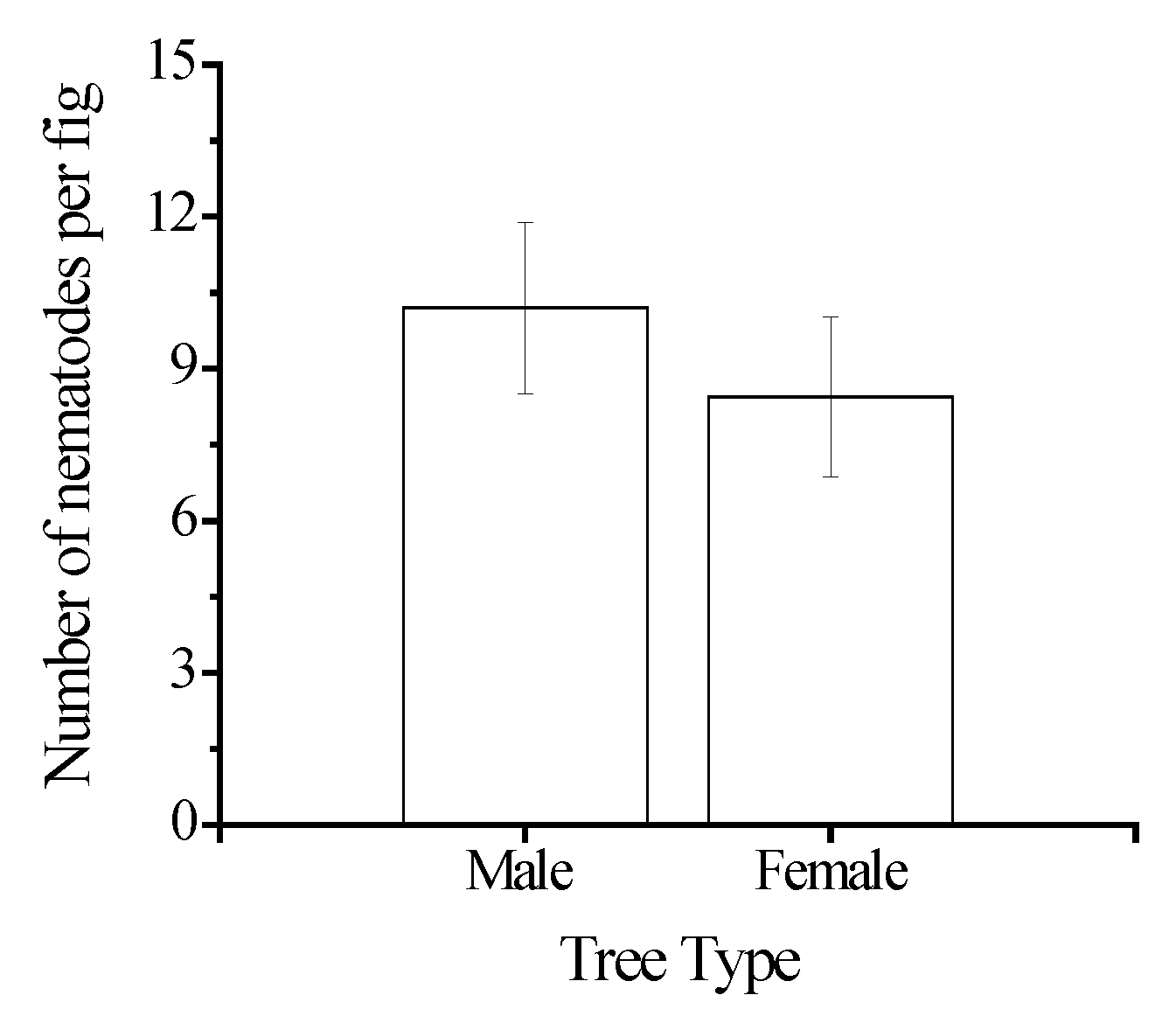{kind=link}
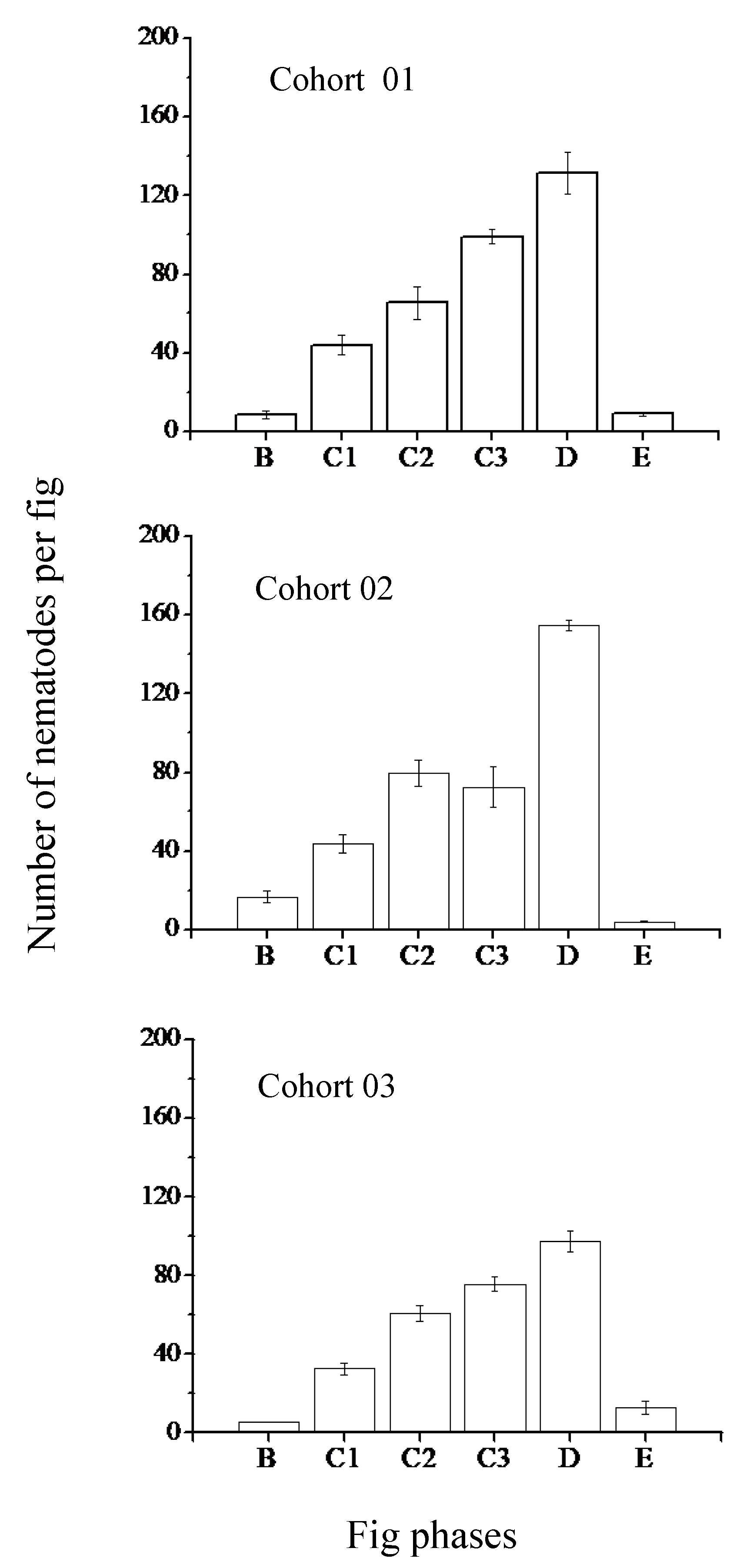{kind=link}
{kind=link}
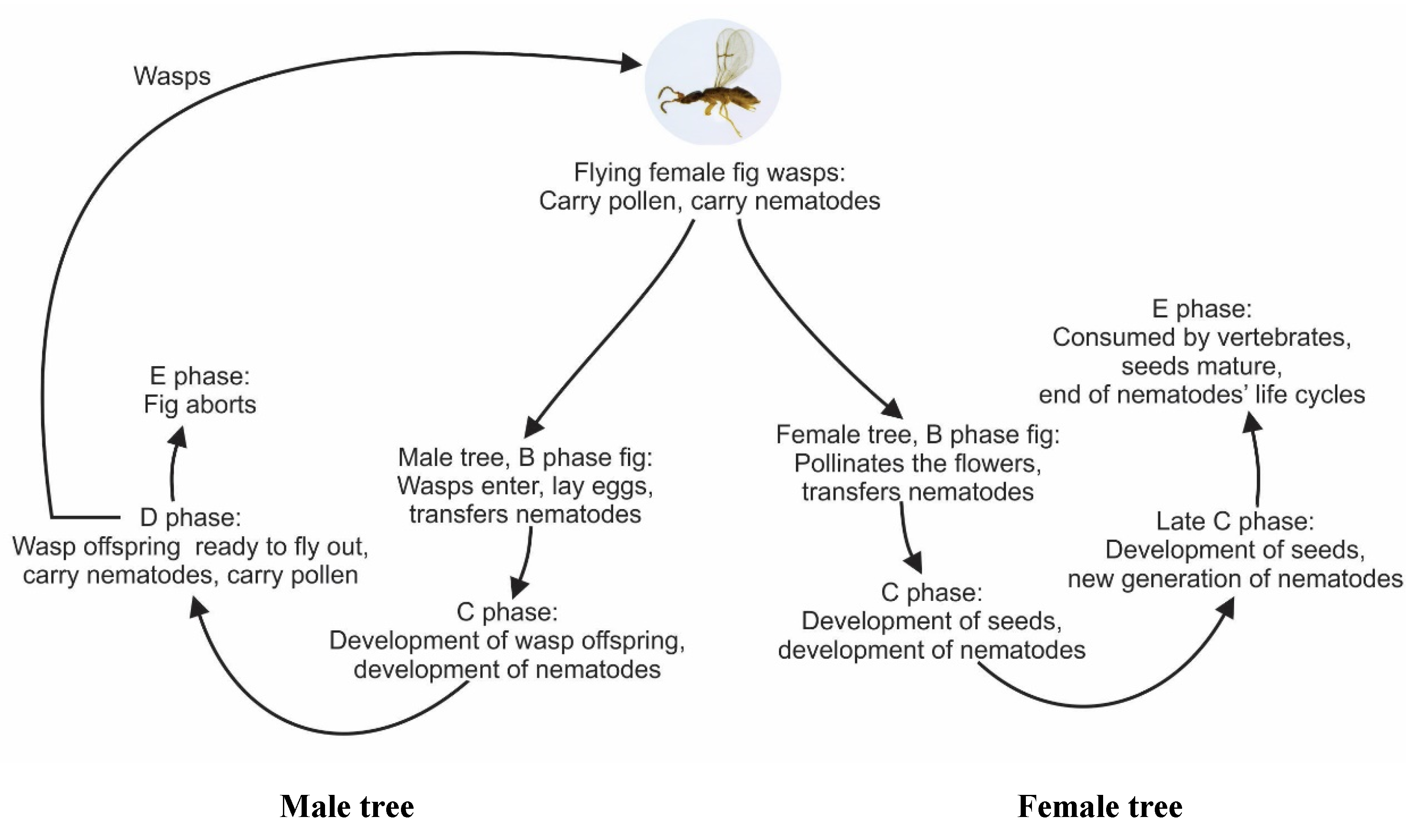{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species and Site
2.2. Sampling Methods and Fig Extraction
2.3. Statistical Analysis
3. Results
3.1. Fruiting Phenology and Fig Wasp Entrants
3.2. Fig Nematodes and Their Life Cycles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shanahan, M.; So, S.; Compton, S.G.; Corlett, R. Fig-eating by vertebrate frugivores: A global review. Biol. Rev. 2001, 76, 529–572. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Compton, S.G.; Liu, M.; Chen, X.Y. Fig trees at the Northern limit of their range: The distributions of cryptic pollinators indicate multiple glacial refugia. Mol. Ecol. 2012, 21, 1687–1701. [Google Scholar] [CrossRef] [PubMed]
- Compton, S.G. One way to be a fig. Afr. Entomol. 1993, 1, 151–158. [Google Scholar]
- Jauharlina, J.; Lindquist, E.E.; Quinnell, R.J.; Robertson, H.G.; Compton, S.G. Fig wasps as vectors of mites and nematodes. Afr. Entomol. 2012, 20, 101–110. [Google Scholar] [CrossRef]
- Pereira, R.A.S.; Semir, J.; Menezes Junior, A.d.O. Pollination and other biotic interactions in figs of Ficus eximia Schott (Moraceae). Rev. Bras. Bot. 2000, 23, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Lachaise, D. Niche separation of African Lissocephala within Ficus drosophilid Community. Oecologia 1977, 31, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Compton, S.G. Moving your sons to safety: Galls containing male fig wasps expand into the centre of figs, away from enemies. PLoS ONE 2012, 7, e30833. [Google Scholar] [CrossRef]
- Kjellberg, F.; Jousselin, E.; Bronstein, J.L.; Patel, A.; Yokoyama, J.; Rasplus, J.Y. Pollination mode in fig wasps: The Predictive power of correlated traits. Proc. R. Soc. Lond. B Biol. Sci. 2001, 268, 1113–1121. [Google Scholar] [CrossRef]
- Weiblen, G.D. How to be a fig wasp. Ann. Rev. Entomol. 2002, 47, 299–330. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.; Muralidharan, S.; Sharma, L.; Borges, R.M. A hitchhiker’s guide to a crowded Syconium: How do fig nematodes find the right ride? Funct. Ecol. 2010, 24, 741–749. [Google Scholar] [CrossRef]
- Michailides, T.J.; Morgan, D.P.; Subbarao, K.V. Fig endospesis: An old disease still a dilemma for California growers. Plant Dis. 1996, 80, 828–840. [Google Scholar] [CrossRef]
- Janzen, D.H. How to be a fig. Annu. Rev. Ecol. Syst. 1979, 10, 13–51. [Google Scholar] [CrossRef]
- Verkerke, W. Structure and Function of the Fig. Experientia 1989, 45, 612–622. [Google Scholar] [CrossRef]
- Jauharlina, J. Fig Trees and Fig Wasps: Their Interactions with Non-Mutualists. Ph.D. Thesis, University of Leeds, Leeds, UK, 2014. [Google Scholar]
- Poinar, G.J.; Poinar, R. The Amber Forest: A Reconstruction of a Vanished World; Princeton University Press: Princeton, NJ, USA, 1999. [Google Scholar] [CrossRef]
- Kanzaki, N.; Giblin-Davis, R.M.; Davies, K.A.; Center, B.J. Teratodiplogaster martini n. Sp. and Parasitodiplogaster doliostoma n. sp. (Nematoda: Diplogastridae) from the syconia of Ficus species from Africa. Nematology 2012, 14, 529–546. [Google Scholar] [CrossRef]
- Poinar, G.O.J.; Herre, E.A. Speciation and adaptive radiation in the fig wasp nematode Parasitodiplogaster Diplogasteridae Rhabditida in Panama. Rev. Nematol. 1991, 14, 361–374. [Google Scholar]
- Vovlas, N.; Inserra, R.N.; Greco, N. Schistonchus caprifici parasitizing caprifig (Ficus carica sylvestris) florets and the relationship with its fig wasp (Blastophaga Psenes) vector. Nematologica 1992, 38, 215–226. [Google Scholar] [CrossRef]
- Wohr, M.; Greeff, J.M.; Kanzaki, N.; Ye, W.; Giblin-Davis, R.M. Molecular and morphological observations on Parasitodiplogaster sycophilon Poinar, 1979 (Nematoda: Diplogastrina) associated with Ficus burkei in Africa. Nematology 2014, 16, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Davies, K.A.; Ye, W.; Kanzaki, N.; Bartholomaeus, F.; Zeng, Y.; Giblin-Davis, R.M. A review of the taxonomy, phylogeny, distribution and co-evolution of Schistonchus Cobb, 1927 with proposal of Ficophagus n. gen. and Martininema n. gen. (Nematoda: Aphelenchoididae). Nematology 2015, 17, 761–829. [Google Scholar] [CrossRef]
- Davies, K.A.; Bartholomaeus, F.; Kanzaki, N.; Ye, W.; Giblin-Davis, R.M. Three new species of Schistonchus (Aphelenchoididae) from the Ficus Subgenus Sycomorus (Moraceae) in Northern Australia. Nematology 2013, 15, 347–362. [Google Scholar] [CrossRef]
- Center, B.J.; Giblin-Davis, R.M.; Herre, E.A.; Chung-Schickler, G.C. Histological comparisons of Parasitism by Schistonchus spp. (Nemata: Aphelenchoididae) in Neotropical Ficus spp. J. Nematol. 1999, 31, 393–406. [Google Scholar]
- Lambert, K.; Bekal, S. Introduction to Plant-Parasitic Nematodes. Plant Health Instr. 2002, 10, 1094–1218, pagination. [Google Scholar] [CrossRef]
- Herre, E.A. Factors affecting the evolution of virulence: Nematode parasites of fig wasps as a case study. Parasitology 1995, 111, S179–S191. [Google Scholar] [CrossRef]
- Compton, S.G.; McLaren, F.A.C. Respiratory adaptations in some male fig wasps. Proc. K. Ned. Akad. Wet. Ser. C Biol. Med. Sci. 1989, 92, 57–71. [Google Scholar]
- Berg, C.C.; Corner, E.J.H. Moraceae-Ficus. Flora Malesiana Series I (Seed Plants). Volume 17, Part 2; National Herbarium of The Netherlands: Leiden, The Netherlands, 2005; ISBN 9071236617. [Google Scholar]
- Galil, J. Fig biology. Endeavour 1977, 1, 52–56. [Google Scholar] [CrossRef]
- Valdeyron, G.; Lloyd, D.G. Sex-differences and flowering phenology in the common fig, Ficus carica L. Evolution 1979, 33, 673–685. [Google Scholar] [CrossRef]
- Sriwati, R.; Takemoto, S.; Futai, K. Seasonal changes in the nematode fauna in pine trees killed by the pinewood nematode, Bursaphelenchus xylophilus. Jpn. J. Nematol. 2006, 36, 87–100. [Google Scholar] [CrossRef] [Green Version]
- Ihaka, R.; Gentelman, R. R: A Language for data analysis and graphics. J. Comput. Graph. Stat. 1996, 5, 299–314. [Google Scholar] [CrossRef]
- Sriwati, R.; Takeuchi-Kaneko, Y.; Jauharlina, J.; Kanzaki, N. Aphelenchoidid nematodes associated with two dominant Ficus species in Aceh, Indonesia. Nematology 2017, 19, 323–331. [Google Scholar] [CrossRef]
- Kanzaki, N.; Tanaka, R.; Giblin-Davis, R.M.; Ragsdale, E.J.; Nguyen, C.N.; Li, H.F.; Lan, Y.C. A preliminary survey of Fig-associated nematodes in the asian subtropics. J. Nematol. 2012, 44, 470. [Google Scholar]
- Woodruff, G.C.; Phillips, P.C. Field studies reveal a close relative of C. elegans thrives in the fresh figs of Ficus septica and disperses on its Ceratosolen pollinating wasps. BMC Ecol. 2018, 18, 26. [Google Scholar] [CrossRef]
- Kiontke, K.; Sudhaus, W. Ecology of Caenorhabditis Species. WormBook: The Online Review of C. elegans Biology; WormBook: Pasadena, CA, USA, 2006. [Google Scholar] [CrossRef]
- Kanzaki, N.; Tsai, I.J.; Tanaka, R.; Hunt, V.L.; Liu, D.; Tsuyama, K.; Maeda, Y.; Namai, S.; Kumagai, R.; Tracey, A.; et al. Biology and genome of a newly discovered sibling species of Caenorhabditis elegans. Nat. Commun. 2018, 9, 3216. [Google Scholar] [CrossRef] [PubMed]
- Weiblen, G.D.; Yu, D.W.; West, S.A. Pollination and parasitism in functionally dioecious figs. Proc. R. Soc. B Biol. Sci. 2001, 268, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Anand, L.R. Association of a nematode Ceratosolenus racemosa Gen. n. Sp.n (Cylindrocorporidae: Rhabditida Goodey, 1939) a wasp (Ceratosolen Sp.n.) and fig Ficus racemosa. Uttar Pradesh J. Zool. 2005, 25, 59–62. [Google Scholar] [CrossRef]
- Davies, K.; Bartholomaeus, F.; Ye, W.; Kanzaki, N.; Giblin-Davis, R. Schistonchus (Aphelenchoididae) from ficus (Moraceae) in Australia, with description of S. aculeata Sp N. Nematology 2010, 12, 935–958. [Google Scholar] [CrossRef]
- Shi, R.-R.; Miao, B.-G.; Segar, S.T.; Zeng, Y.; Wang, B.; Peng, Y.-Q. Are nematodes costly to fig tree–fig wasp mutualists? Entomol. Exp. Appl. 2019, 167, 1000–1011. [Google Scholar] [CrossRef]
- Van Goor, J.; Piatscheck, F.; Houston, D.D.; Nason, J.D. Figs, Pollinators, and Parasites: A longitudinal study of the effects of nematode infection on fig wasp fitness. Acta Oecologica 2018, 90, 140–150. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jauharlina; Oktarina, H.; Sriwati, R.; Sayuthi, M.; Kanzaki, N.; Quinnell, R.J.; Compton, S.G. Association of Fig Pollinating Wasps and Fig Nematodes inside Male and Female Figs of a Dioecious Fig Tree in Sumatra, Indonesia. Insects 2022, 13, 320. https://doi.org/10.3390/insects13040320
Jauharlina, Oktarina H, Sriwati R, Sayuthi M, Kanzaki N, Quinnell RJ, Compton SG. Association of Fig Pollinating Wasps and Fig Nematodes inside Male and Female Figs of a Dioecious Fig Tree in Sumatra, Indonesia. Insects. 2022; 13(4):320. https://doi.org/10.3390/insects13040320
Chicago/Turabian StyleJauharlina, Hartati Oktarina, Rina Sriwati, Muhammad Sayuthi, Natsumi Kanzaki, Rupert J. Quinnell, and Stephen G. Compton. 2022. "Association of Fig Pollinating Wasps and Fig Nematodes inside Male and Female Figs of a Dioecious Fig Tree in Sumatra, Indonesia" Insects 13, no. 4: 320. https://doi.org/10.3390/insects13040320