
Land Use Influences the Composition and Antimicrobial Effects of Propolis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Propolis Collection and Extraction
2.2. Gas Chromatography Mass Spectroscopy (GC-MS)
2.3. Land Use Analysis
2.4. MIC50
2.5. Statistical Analyses
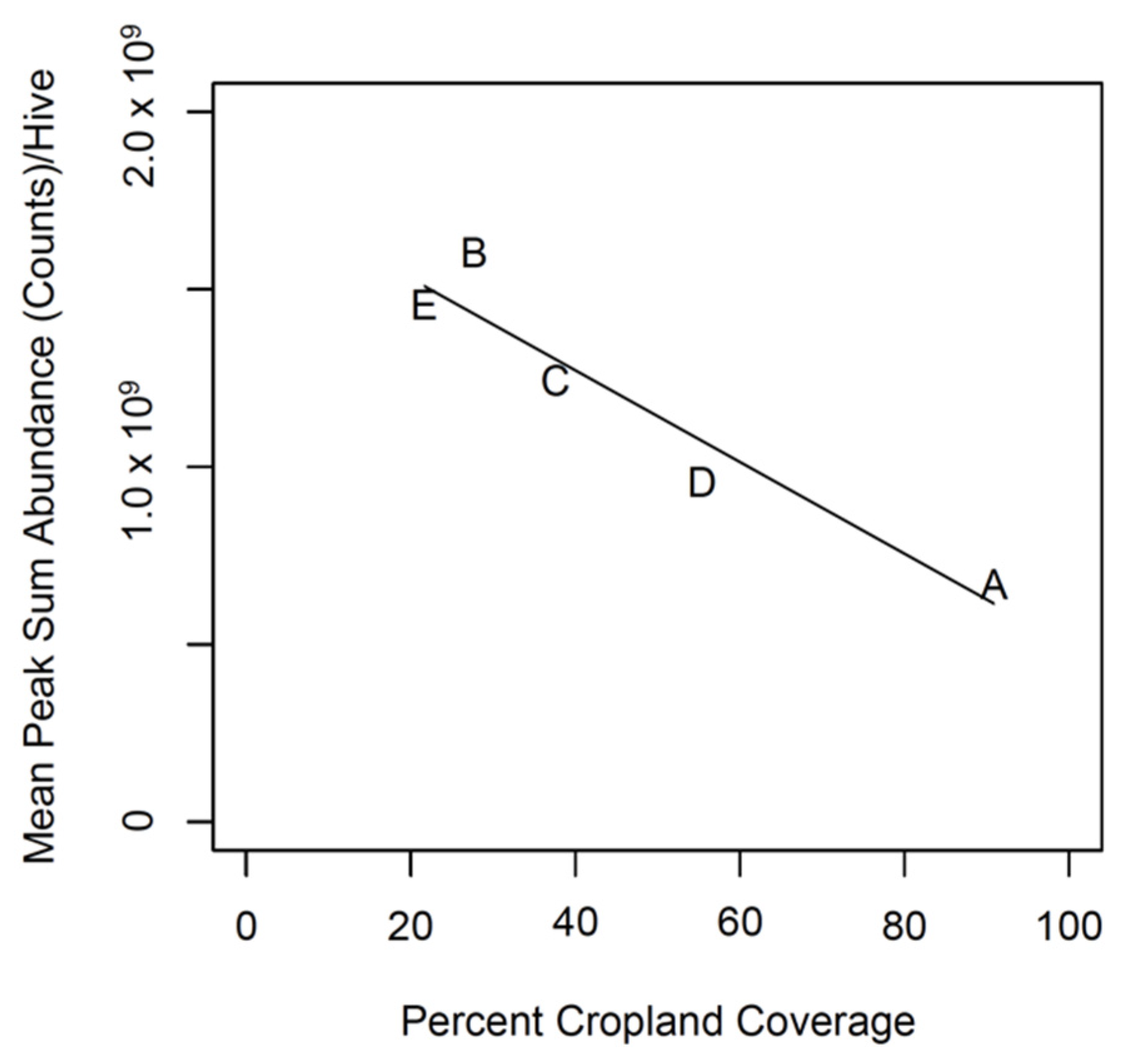
2.5.1. Relationship between Land Use and Propolis Production
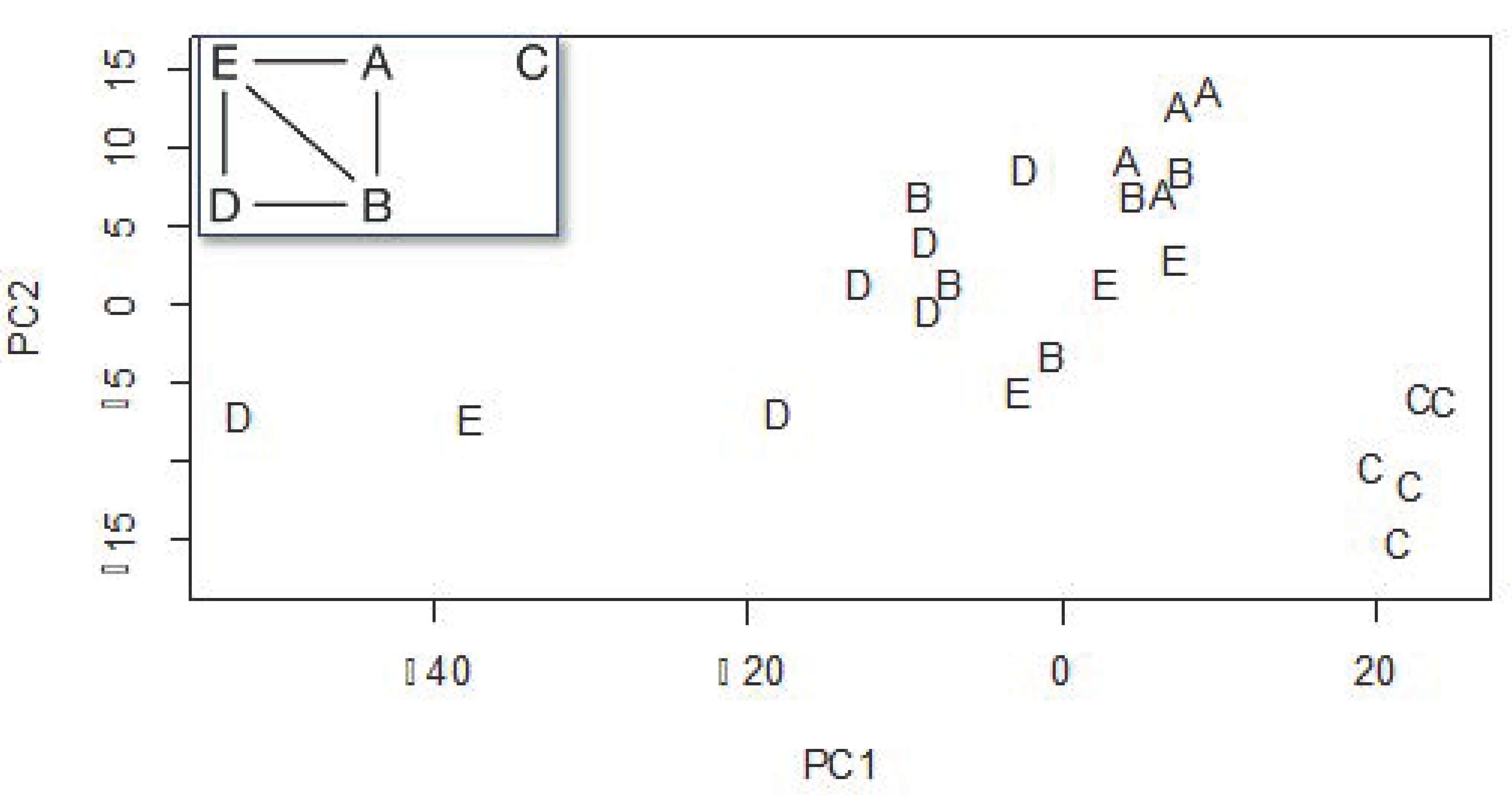
2.5.2. Comparison of Propolis Compounds among Apiaries
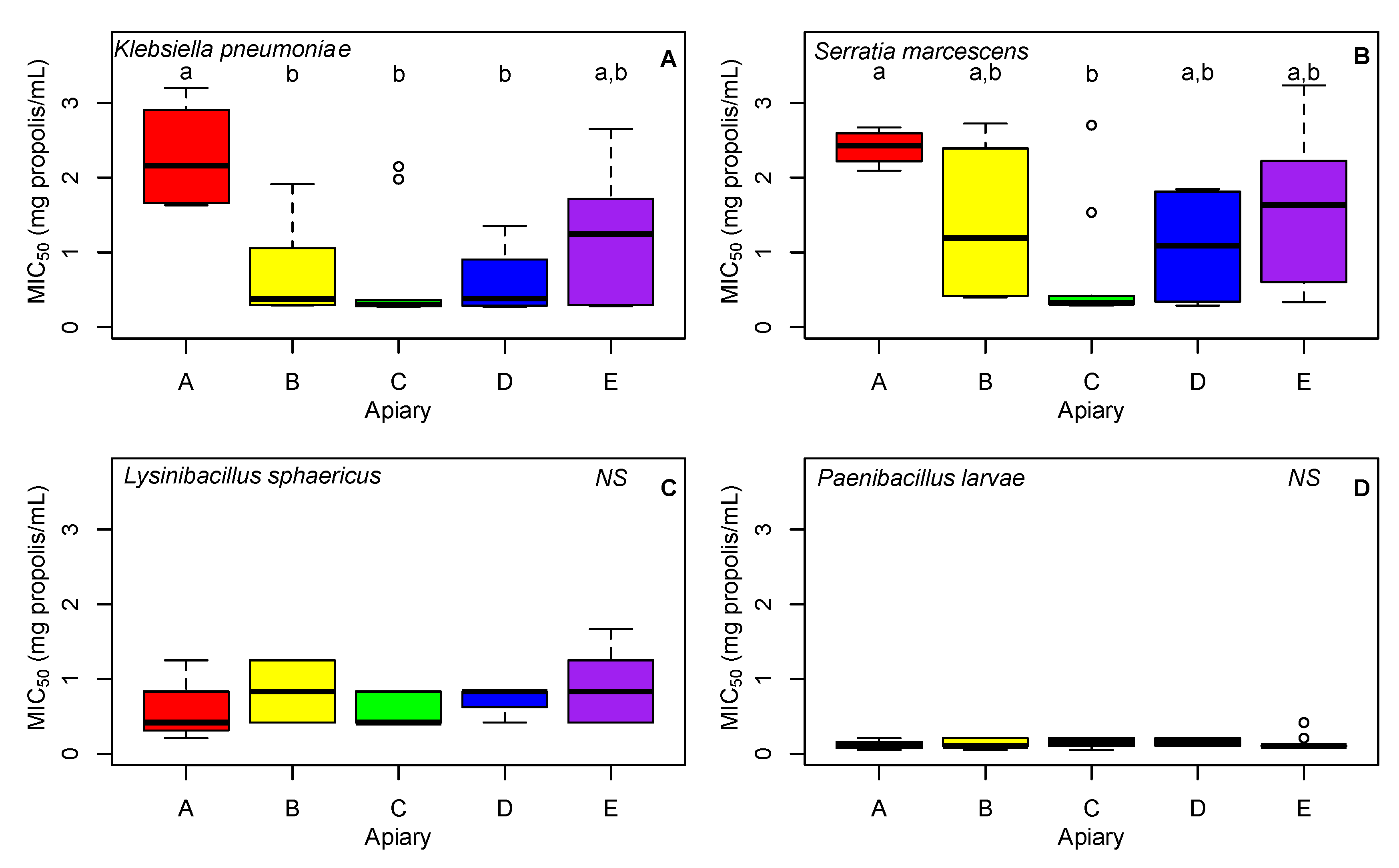
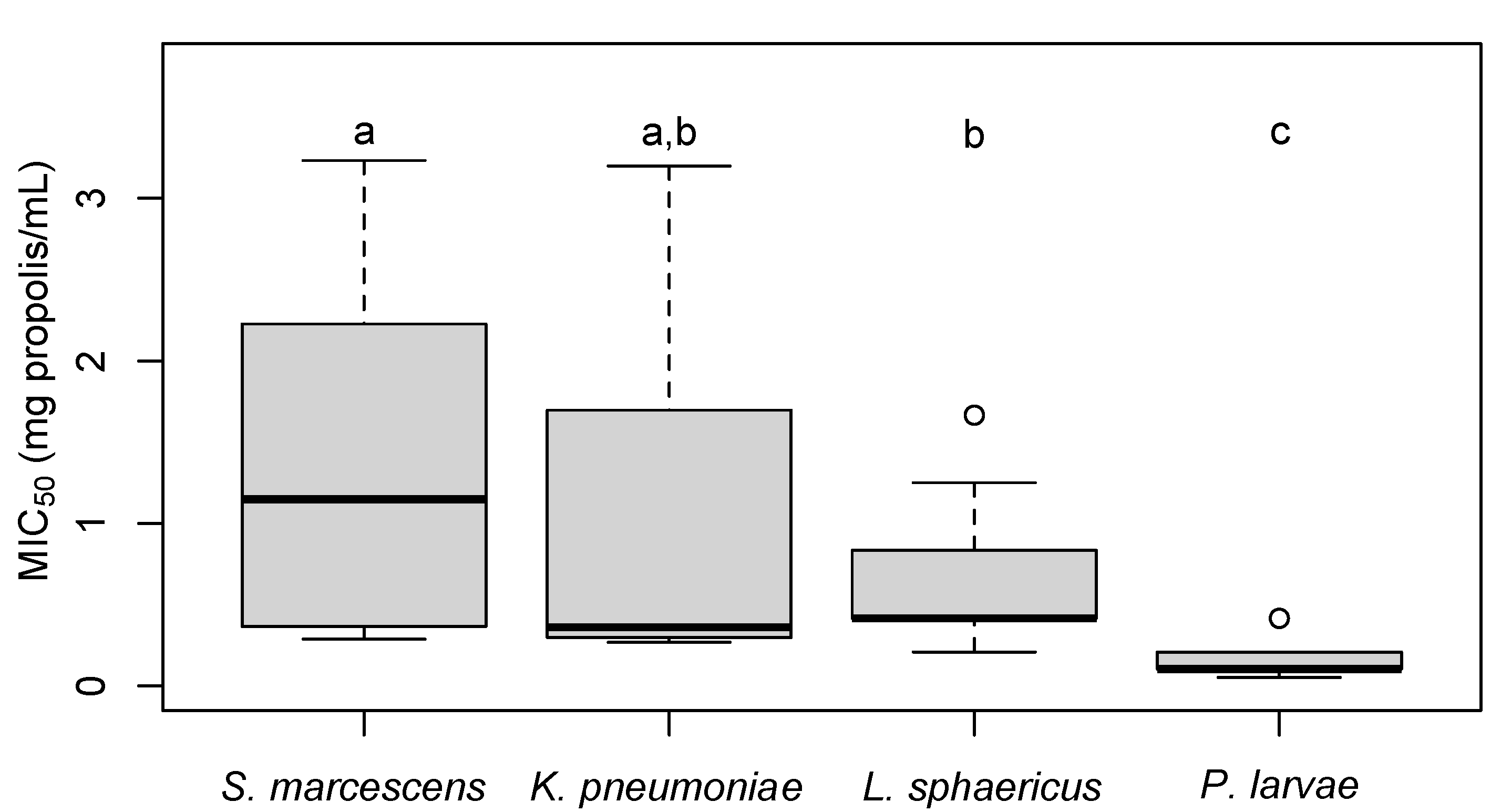
2.5.3. Effectiveness of Propolis against Pathogens
3. Results
3.1. Relationship between Land Use and Propolis Production
3.2. Comparison of Propolis Compounds among Apiaries
3.3. MIC50
3.3.1. Klebsiella pneumoniae
3.3.2. Serratia marcescens
3.3.3. Lysinibacillus sphaericus and Paenibaccillus larvae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simone-Finstrom, M.; Spivak, M. Propolis and bee health: The natural history and significance of resin use by honey bees. Apidologie 2010, 41, 295–311. [Google Scholar] [CrossRef] [Green Version]
- Simone-Finstrom, M.; Borba, R.S.; Wilson, M.; Spivak, M. Propolis counteracts some threats to honey bee health. Insects 2017, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cremer, S. Social immunity in insects. Curr. Biol. 2019, 29, R458–R463. [Google Scholar] [CrossRef]
- Simone, M.; Evans, J.D.; Spivak, M. Resin collection and social immunity in honey bees. Evolution 2009, 63, 3016–3022. [Google Scholar] [CrossRef]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect. Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartal, M.; Kaya, S.; Kurucu, S. GC-MS analysis of propolis samples from two different regions of Turkey. Z. Naturforsch. Sect. C J. Biosci. 2002, 57, 905–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AL-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Antimicrobial Activities of European Propolis Collected from Various Geographic Origins Alone and in Combination with Antibiotics. Medicines 2018, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.; Zhang, C.P.; Wang, K.; Li, G.Q.; Hu, F.L. Recent advances in the chemical composition of propolis. Molecules 2014, 19, 19610–19632. [Google Scholar] [CrossRef] [Green Version]
- Borba, R.S.; Klyczek, K.K.; Mogen, K.L.; Spivak, M. Seasonal benefits of a natural propolis envelope to honey bee immunity and colony health. J. Exp. Biol. 2015, 218, 3689–3699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Przybyłek, I.; Karpiński, T.M. Antibacterial properties of propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, J.; Chang, F.R.; Wang, H.K.; Park, Y.K.; Ikegaki, M.; Kilgore, N.; Lee, K.H. Anti-AIDS agents. 48. Anti-HIV activity of moronic acid derivatives and the new melliferone-related triterpenoid isolated from Brazilian propolis. J. Nat. Prod. 2001, 64, 1278–1281. [Google Scholar] [CrossRef] [PubMed]
- Vynograd, N.; Vynograd, I.; Sosnowski, Z. A comparative multi-centre study of the efficacy of propolis, acyclovir and placebo in the treatment of genital herpes (HSV). Phytomedicine 2000, 7, 1–6. [Google Scholar] [CrossRef]
- Kujumgiev, A.; Tsvetkova, I.; Serkedjieva, Y.; Bankova, V.; Christov, R.; Popov, S. Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. J. Ethnopharmacol. 1999, 64, 235–240. [Google Scholar] [CrossRef]
- Ghisalberti, E.L. Propolis: A Review. Bee World 1979, 60, 59–84. [Google Scholar] [CrossRef]
- Bankova, V.; Marcucci, M.C.; Simova, S.; Nikolova, N.; Kujumgiev, A.; Popov, S. Antibacterial diterpenic acids from Brazilian propolis. Z. Naturforsch. Sect. C J. Biosci. 1996, 51, 277–280. [Google Scholar] [CrossRef]
- Haydak, M.H. Propolis. Rep. Iowa State Apiarist 1953, 74–87. [Google Scholar]
- Veiga, F.F.; Gadelha, M.C.; da Silva, M.R.T.; Costa, M.I.; Kischkel, B.; de Castro-Hoshino, L.V.; Sato, F.; Baesso, M.L.; Voidaleski, M.F.; Vasconcellos-Pontello, V.; et al. Propolis extract for onychomycosis topical treatment: From bench to clinic. Front. Microbiol. 2018, 9, 779. [Google Scholar] [CrossRef] [Green Version]
- Galletti, J.; Tobaldini-Valerio, F.K.; Silva, S.; Kioshima, É.S.; Trierveiler-Pereira, L.; Bruschi, M.; Negri, M.; Estivalet Svidzinski, T.I. Antibiofilm activity of propolis extract on Fusarium species from onychomycosis. Future Microbiol. 2017, 12, 1311–1321. [Google Scholar] [CrossRef] [Green Version]
- Agüero, M.B.; Svetaz, L.; Baroni, V.; Lima, B.; Luna, L.; Zacchino, S.; Saavedra, P.; Wunderlin, D.; Feresin, G.E.; Tapia, A. Urban propolis from San Juan province (Argentina): Ethnopharmacological uses and antifungal activity against Candida and dermatophytes. Ind. Crops Prod. 2014, 57, 166–173. [Google Scholar] [CrossRef]
- Waller, S.B.; Peter, C.M.; Hoffmann, J.F.; Picoli, T.; da G. Osório, L.; Chaves, F.; Zani, J.L.; de Faria, R.O.; de Mello, J.R.B.; Meireles, M.C.A. Chemical and cytotoxic analyses of brown Brazilian propolis (Apis mellifera) and its in vitro activity against itraconazole-resistant Sporothrix brasiliensis. Microb. Pathog. 2017, 105, 117–121. [Google Scholar] [CrossRef]
- Tobaldini-Valerio, F.K.; Bonfim-Mendonça, P.S.; Rosseto, H.C.; Bruschi, M.L.; Henriques, M.; Negri, M.; Silva, S.; Svidzinski, T.I.E. Propolis: A potential natural product to fight Candida species infections. Future Microbiol. 2016, 11, 1035–1046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ota, C.; Unterkircher, C.; Fantinato, V.; Shimizu, M.T. Antifungal activity of propolis on different species of Candida. Mycoses 2001, 44, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Marcucci, M.C. Propolis: Chemical composition, biological properties and therapeutic activity. Apidologie 1995, 26, 83–99. [Google Scholar] [CrossRef]
- Grimont, P.A.; Grimont, F. The genus Serratia. Annu. Rev. Microbiol. 1978, 32, 221–248. [Google Scholar] [CrossRef]
- Fünfhaus, A.; Ebeling, J.; Genersch, E. Bacterial pathogens of bees. Curr. Opin. Insect. Sci. 2018, 26, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Disayathanoowat, T.; Yoshiyama, M.; Kimura, K.; Chantawannakul, P. Isolation and characterization of bacteria from the midgut of the Asian honey bee (Apis cerana indica). J. Apic. Res. 2012, 51, 312–319. [Google Scholar] [CrossRef]
- Erban, T.; Ledvinka, O.; Kamler, M.; Nesvorna, M.; Hortova, B.; Tyl, J.; Titera, D.; Markovic, M.; Hubert, J. Honeybee (Apis mellifera)-associated bacterial community affected by American foulbrood: Detection of Paenibacillus larvae via microbiome analysis. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef]
- Ebeling, J.; Knispel, H.; Hertlein, G.; Fünfhaus, A.; Genersch, E. Biology of Paenibacillus larvae, a deadly pathogen of honey bee larvae. Appl. Microbiol. Biotechnol. 2016, 100, 7387–7395. [Google Scholar] [CrossRef]
- Poppinga, L.; Genersch, E. Molecular pathogenesis of American Foulbrood: How Paenibacillus larvae kills honey bee larvae. Curr. Opin. Insect. Sci. 2015, 10, 29–36. [Google Scholar] [CrossRef]
- Shanks, J.L.; Haigh, A.M.; Riegler, M.; Spooner-Hart, R.N. First confirmed report of a bacterial brood disease in stingless bees. J. Invertebr. Pathol. 2017, 144, 7–10. [Google Scholar] [CrossRef]
- Stein, S.E.; Scott, D.R. Optimization and testing of mass spectral library search algorithms for compound identification. J. Am. Soc. Mass Spectrom. 1994, 5, 859–866. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Yang, Z.; Di, L.; Mueller, R. CropScape: A web service based application for exploring and disseminating US conterminous geospatial Cropland Data Products for decision support. Comput. Electron. Agric. 2012, 84, 111–123. [Google Scholar] [CrossRef]
- Eckert, J.E. The flight range of the honeybee. J. Agric. Res. 1933, 47, 257–285. [Google Scholar]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Grange, J.; Davey, R.W. Antibacterial properties of propolis (bee glue). J. R. Soc. Med. 1990, 83, 159–160. [Google Scholar] [CrossRef]
- Pamplona-Zomenhan, L.C.; Pamplona, B.C.; da Silva, C.B.; Marcucci, M.C.; Mimica, L.M.J. Evaluation of the in vitro antimicrobial activity of an ethanol extract of Brazilian classified propolis on strains of Staphylococcus aureus. Braz. J. Microbiol. 2011, 42, 1259–1264. [Google Scholar] [CrossRef] [Green Version]
- Collyer ML, A.D. RRPP: An R package for fitting linear models to high-dimensional data using residual randomization. Methods Ecol. Evol. 2018, 9, 1772–1779. [Google Scholar] [CrossRef] [Green Version]
- Collyer, M.; Adams, D. RRPP: Linear Model Evaluation with Randomized Residuals in a Permutation Procedure. R Package Version 0.4.0. 2019. Available online: https://CRAN.R-project.org/package=RRPP (accessed on 1 July 2021).
- Sforcin, J.M. Biological Properties and Therapeutic Applications of Propolis. Phyther. Res. 2016, 30, 894–905. [Google Scholar] [CrossRef]
- Lindenfelser, L.A. In vivo activity of propolis against Bacillus larvae. J. Invertebr. Pathol. 1968, 12, 129–131. [Google Scholar] [CrossRef]
- Wilson, M.B.; Brinkman, D.; Spivak, M.; Gardner, G.; Cohen, J.D. Regional variation in composition and antimicrobial activity of US propolis against Paenibacillus larvae and Ascosphaera apis. J. Invertebr. Pathol. 2015, 124, 44–50. [Google Scholar] [CrossRef]
- Leonhardt, S.D. Chemical Ecology of Stingless Bees. J. Chem. Ecol. 2017, 43, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Lambert, R.J.; Stratford, M. Weak-acid preservatives: Modelling microbial inhibition and response. J. Appl. Microbiol. 1999, 86, 157–164. [Google Scholar] [CrossRef]
- Salmond, C.V.; Kroll, R.G.; Booth, I.R. The effect of food preservatives on pH homeostasis in Escherichia coli. J. Gen. Microbiol. 1984, 130, 2845–2850. [Google Scholar] [CrossRef] [Green Version]
- Rubab, M.; Chelliah, R.; Saravanakumar, K.; Barathikannan, K.; Wei, S.; Kim, J.R.; Yoo, D.; Wang, M.H.; Oh, D.H. Bioactive potential of 2-methoxy-4-vinylphenol and benzofuran from Brassica oleracea L. var. capitate f, rubra (Red Cabbage) on oxidative and microbiological stability of beef meat. Foods 2020, 9, 568. [Google Scholar] [CrossRef]
- Renuka, J.; Reddy, K.I.; Srihari, K.; Jeankumar, V.U.; Shravan, M.; Sridevi, J.P.; Yogeeswari, P.; Babu, K.S.; Sriram, D. Design, synthesis, biological evaluation of substituted benzofurans as DNA gyraseB inhibitors of Mycobacterium tuberculosis. Bioorg. Med. Chem. 2014, 22, 4924–4934. [Google Scholar] [CrossRef]
- Tomás-Barberán, F.A.; García-Viguera, C.; Vit-Olivier, P.; Ferreres, F.; Tomás-Lorente, F. Phytochemical evidence for the botanical origin of tropical propolis from Venezuela. Phytochemistry 1993, 34, 191–196. [Google Scholar] [CrossRef]
- Greenaway, W.; Scaysbrook, T.; Whatley, F.R. The composition and plant origins of propolis: A report of work at oxford. Bee World 1990, 71, 107–118. [Google Scholar] [CrossRef]
- García-Viguera, C.; Ferreres, F.; Tomás-B arberán, F.A. Study of canadian propolis by gc-ms and hplc. Z. Naturforsch. Sect. C J. Biosci. 1993, 48, 731–735. [Google Scholar] [CrossRef]
- Bankova, V. Chemical diversity of propolis and the problem of standardization. J. Ethnopharmacol. 2005, 100, 114–117. [Google Scholar] [CrossRef]
- Drescher, N.; Wallace, H.M.; Katouli, M.; Massaro, C.F.; Leonhardt, S.D. Diversity matters: How bees benefit from different resin sources. Oecologia 2014, 176, 943–953. [Google Scholar] [CrossRef]
- Wilson, M.B.; Spivak, M.; Hegeman, A.D.; Rendahl, A.; Cohen, J.D. Metabolomics Reveals the Origins of Antimicrobial Plant Resins Collected by Honey Bees. PLoS ONE 2013, 8, e77512. [Google Scholar] [CrossRef] [Green Version]
- Dezmirean, D.S.; Paşca, C.; Moise, A.R.; Bobiş, O. Plant Sources Responsible for the Chemical Composition and Main Bioactive Properties of Poplar-Type Propolis. Plants 2020, 10, 22. [Google Scholar] [CrossRef]
- Millard, J.; Outhwaite, C.L.; Kinnersley, R.; Freeman, R.; Gregory, R.D.; Adedoja, O.; Gavini, S.; Kioko, E.; Kuhlmann, M.; Ollerton, J.; et al. Global effects of land-use intensity on local pollinator biodiversity. Nat. Commun. 2021, 12, 2902. [Google Scholar] [CrossRef]
- Dolezal, A.G.; Clair, A.L.S.; Zhang, G.; Toth, A.L.; O’Neal, M.E. Native habitat mitigates feast–famine conditions faced by honey bees in an agricultural landscape. Proc. Natl. Acad. Sci. USA 2019, 116, 25147–25155. [Google Scholar] [CrossRef] [Green Version]
- Smart, M.D.; Otto, C.R.V.; Carlson, B.L.; Roth, C.L. The influence of spatiotemporally decoupled land use on honey bee colony health and pollination service delivery. Environ. Res. Lett. 2018, 13, 084016. [Google Scholar] [CrossRef]
- Smart, M.; Pettis, J.; Rice, N.; Browning, Z.; Spivak, M. Linking measures of colony and individual honey bee health to survival among apiaries exposed to varying agricultural land use. PLoS ONE 2016, 11, e0152685. [Google Scholar] [CrossRef] [Green Version]
- Tsvetkov, N.; Samson-Robert, O.; Sood, K.; Patel, H.S.; Malena, D.A.; Gajiwala, P.H.; Maciukiewicz, P.; Fournier, V.; Zayed, A. Chronic exposure to neonicotinoids reduces honey bee health near corn crops. Science 2017, 356, 1395–1397. [Google Scholar] [CrossRef] [Green Version]
- Tomšič, R.; Heath, D.; Heath, E.; Markelj, J.; Kandolf Borovšak, A.; Prosen, H. Determination of Neonicotinoid Pesticides in Propolis with Liquid Chromatography Coupled to Tandem Mass Spectrometry. Molecules 2020, 25, 5870. [Google Scholar] [CrossRef]
- Smith, D.; Davis, A.; Hitaj, C.; Hellerstein, D.; Preslicka, A.; Kogge, E.; Mushet, D.; Lonsdorf, E. The contribution of land cover change to the decline of honey yields in the Northern Great Plains. Environ. Res. Lett. 2021, 16, 064050. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Apiary A | Apiary B | Apiary C | Apiary D | Apiary E |
|---|---|---|---|---|---|
| 1 | benzoic acid (63%) | benzoic acid (53%) | benzoic acid (60%) | benzoic acid (40%) | benzoic acid (42%) |
| 2 | coumaran (15%) | coumaran (17%) | coumaran (20%) | coumaran (29%) | coumaran (18%) |
| 3 | 2-methoxy-4-vinylphenol (8%) | 2-methoxy-4-vinylphenol (7%) | 2-methoxy-4-vinylphenol (7%) | 2-methoxy-4-vinylphenol (11%) | trans-cinnamic acid (10%) |
| 4 | 3-methoxyaceto-phenone (7%) | alpha bisabolol (5%) | benzyl alcohol (4%) | 3-methoxyaceto-phenone (9%) | 2-methoxy-4-vinylphenol (7%) |
| 5 | benzyl alcohol (6%) | benzyl benzoate (5%) | benzyl benzoate (2%) | benzyl alcohol (7%) | alpha bisabolol (5%) |
| 6 | benzyl benzoate (2%) | benzyl alcohol (3%) | alpha bisabolol (1%) | alpha bisabolol (3%) | curcumene (4%) |
| 7 | cedrane (1%) | trans-cinnamic acid (3%) | acetophenone (1%) | benzyl benzoate (2%) | benzyl benzoate (3%) |
| 8 | 2-benzoylaminoethanol (1%) | naphthalene (2%) | curcumene (1%) | trans-isoeugenol (2%) | vanillin (3%) |
| 9 | 4-benzyloxybenzoic acid (1%) | vanillin (1%) | trans-isoeugenol (1%) | trans-cinnamic acid (1%) | benzyl alcohol (3%) |
| 10 | 4-hydroxy-3-methylacetophenone (1%) | acetophenone (0.4%) | vanillin (1%) | acetophenone (1%) | alpha farnesene (1%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orth, A.J.; Curran, E.H.; Haas, E.J.; Kraemer, A.C.; Anderson, A.M.; Mason, N.J.; Fassbinder-Orth, C.A. Land Use Influences the Composition and Antimicrobial Effects of Propolis. Insects 2022, 13, 239. https://doi.org/10.3390/insects13030239
Orth AJ, Curran EH, Haas EJ, Kraemer AC, Anderson AM, Mason NJ, Fassbinder-Orth CA. Land Use Influences the Composition and Antimicrobial Effects of Propolis. Insects. 2022; 13(3):239. https://doi.org/10.3390/insects13030239
Chicago/Turabian StyleOrth, Amara J., Emma H. Curran, Eric J. Haas, Andrew C. Kraemer, Audrey M. Anderson, Nicholas J. Mason, and Carol A. Fassbinder-Orth. 2022. "Land Use Influences the Composition and Antimicrobial Effects of Propolis" Insects 13, no. 3: 239. https://doi.org/10.3390/insects13030239