
Lateralization of Courtship Traits Impacts Pentatomid Male Mating Success—Evidence from Field Observations



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Observations
2.2. Statistical Analysis
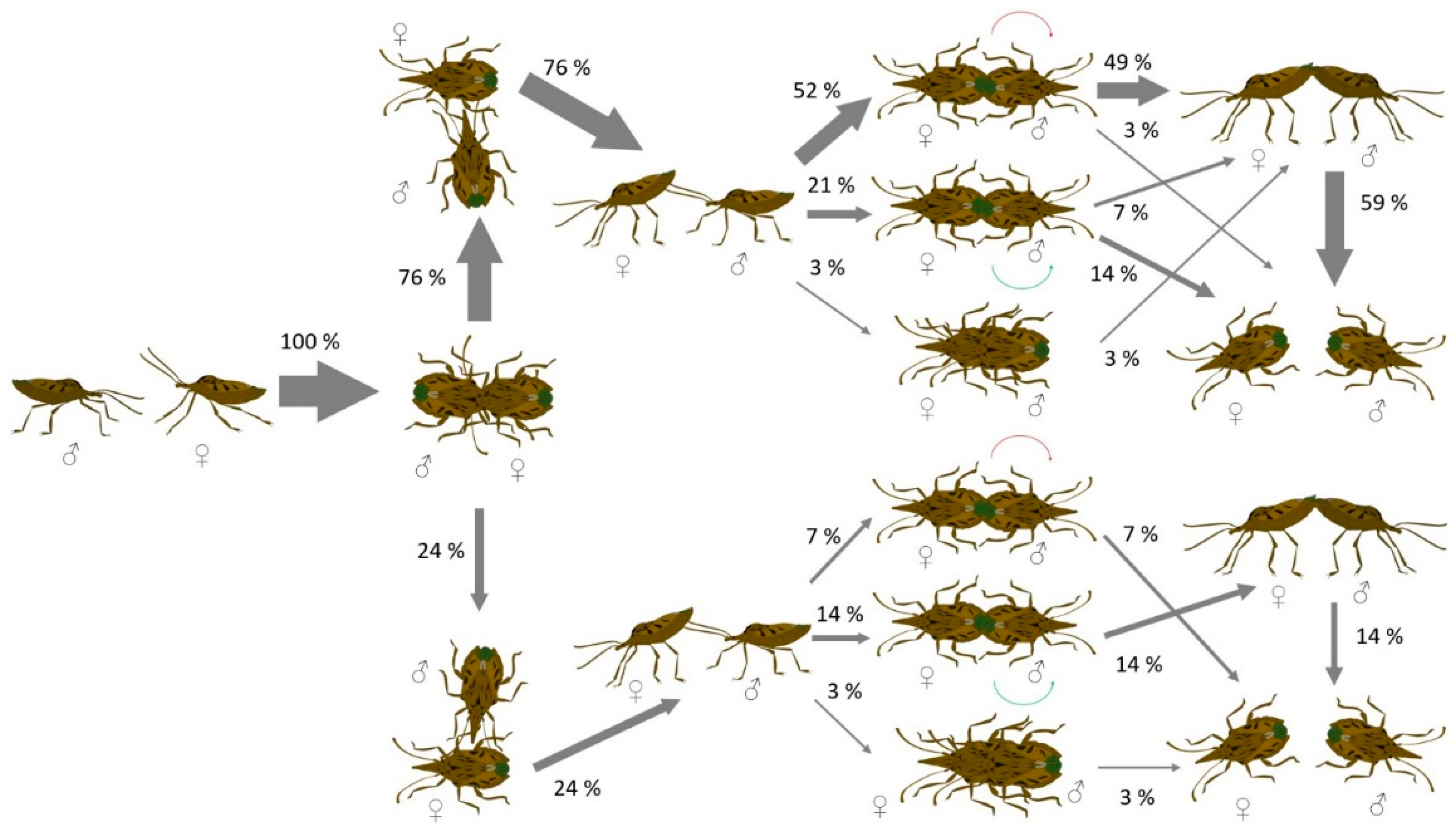
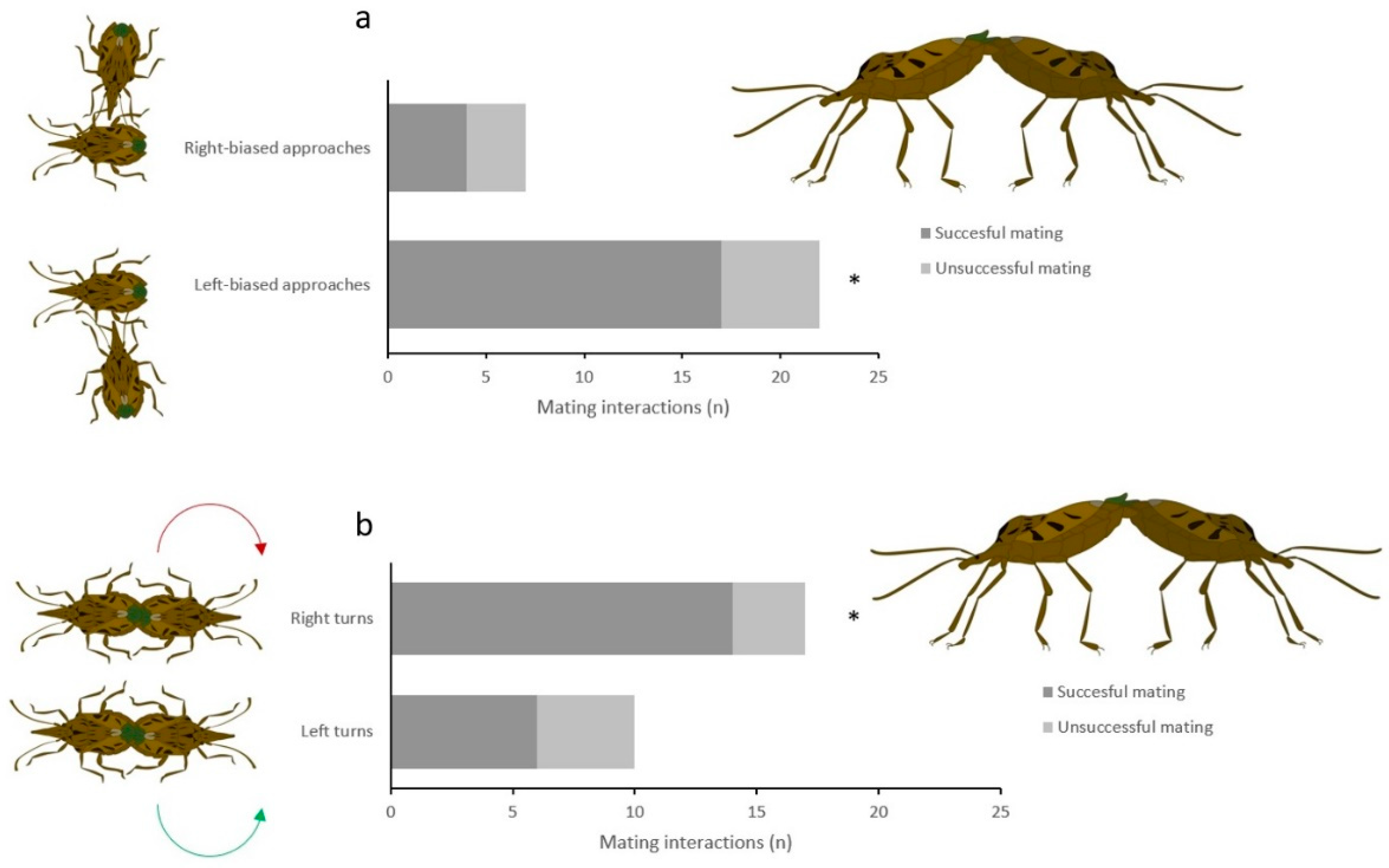
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vallortigara, G.; Rogers, L.J. Survival with an asymmetrical brain: Advantages and disadvantages of cerebral lateralization. Behav. Brain. Sci. 2005, 28, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Romano, D.; Benelli, G.; Stefanini, C. Escape and surveillance asymmetries in locusts exposed to a Guinea fowl-mimicking robot predator. Sci. Rep. 2017, 7, 12825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallortigara, G.; Andrew, R.J. Lateralization of response by chicks to change in a model partner. Anim. Behav. 1991, 41, 187–194. [Google Scholar] [CrossRef]
- Cantalupo, C.; Bisazza, A.; Vallortigara, G. Lateralization of predator-evasion response in a teleost fish (Girardinus falcatus). Neuropsychologia 1995, 33, 1637–1646. [Google Scholar] [CrossRef]
- Mascetti, G.G.; Rugger, M.; Vallortigara, G. Visual lateralization and monocular sleep in the domestic chick. Cogn. Brain Res. 1999, 7, 451–463. [Google Scholar] [CrossRef]
- Lippolis, G.; Bisazza, A.; Rogers, L.J.; Vallortigara, G. Lateralisation of predator avoidance responses in three species of toads. Laterality 2002, 7, 163–183. [Google Scholar] [CrossRef]
- George, I.; Hara, E.; Hessler, N.A. Behavioral and neural lateralization of vision in courtship singing of the zebra finch. J. Neurobiol. 2006, 66, 1164–1173. [Google Scholar] [CrossRef]
- Dadda, M.; Agrillo, C.; Bisazza, A.; Brown, C. Laterality enhances numerical skills in the guppy, Poecilia reticulata. Front. Behav. Neurosci. 2015, 9, 285. [Google Scholar] [CrossRef] [Green Version]
- Sovrano, V.A.; Quaresmini, C.; Stancher, G. Tortoises in front of mirrors: Brain asymmetries and lateralized behaviours in the tortoise (Testudo hermanni). Behav. Brain Res. 2018, 352, 183–186. [Google Scholar] [CrossRef]
- Kight, S.L.; Steelman, L.; Coffey, G.; Lucente, J.; Castillo, M. Evidence of population-level lateralized behaviour in giant water bugs, Belostoma flumineum Say (Heteroptera: Belostomatidae): T-maze turning is left biased. Behav. Process. 2008, 79, 66–69. [Google Scholar] [CrossRef]
- Letzkus, P.; Boeddeker, N.; Wood, J.T.; Zhang, S.W.; Srinivasan, M.V. Lateralization of visual learning in the honeybee. Biol. Lett. 2008, 4, 16–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frasnelli, E.; Vallortigara, G.; Rogers, L.J. Left—Right asymmetries of behaviour and nervous system in invertebrates. Neurosci. Biobehav. Rev. 2012, 36, 1273–1291. [Google Scholar] [CrossRef] [PubMed]
- Hunt, E.R.; O’Shea-Wheller, T.; Albery, G.F.; Bridger, T.H.; Gumn, M.; Franks, N.R. Ants show a leftward turning bias when exploring unknown nest sites. Biol. Lett. 2014, 10, 20140945. [Google Scholar] [CrossRef] [Green Version]
- Benelli, G.; Romano, D.; Messing, R.H.; Canale, A. First report of behavioural lateralisation in mosquitoes: Right-biased kicking behaviour against males in females of the Asian tiger mosquito, Aedes albopictus. Parasitol. Res. 2015, 114, 1613–1617. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Donati, E.; Romano, D.; Stefanini, C.; Messing, R.H.; Canale, A. Lateralisation of aggressive displays in a tephritid fly. Sci. Nat. 2015, 102, 1. [Google Scholar] [CrossRef]
- Romano, D.; Canale, A.; Benelli, G. Do right-biased boxers do it better? Population-level asymmetry of aggressive displays enhances fighting success in blowflies. Behav. Process. 2015, 113, 159–162. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D.; Messing, R.H.; Canale, A. Population-level lateralized aggressive and courtship displays make better fighters not lovers: Evidence from a fly. Behav. Process. 2015, 115, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Romano, D.; Donati, E.; Canale, A.; Messing, R.H.; Benelli, G.; Stefanini, C. Lateralized courtship in a parasitic wasp. Laterality 2016, 21, 243–254. [Google Scholar] [CrossRef]
- Romano, D.; Benelli, G.; Stefanini, C.; Desneux, N.; Ramirez-Romero, R.; Canale, A.; Lucchi, A. Behavioral asymmetries in the mealybug parasitoid Anagyrus sp. near pseudococci: Does lateralized antennal tapping predict male mating success? J. Pest Sci. 2018, 91, 341–349. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D.; Stefanini, C.; Kavallieratos, N.G.; Athanassiou, C.G.; Canale, A. Asymmetry of mating behaviour affects copulation success in two stored-product beetles. J. Pest Sci. 2017, 90, 547–556. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D.; Kavallieratos, N.; Conte, G.; Stefanini, C.; Mele, M.; Canale, A. Multiple behavioural asymmetries impact male mating success in the khapra beetle, Trogoderma granarium. J. Pest Sci. 2017, 90, 901–909. [Google Scholar] [CrossRef]
- Romano, D.; Kavallieratos, N.G.; Athanassiou, C.G.; Stefanini, C.; Canale, A.; Benelli, G. Impact of geographical origin and rearing medium on mating success and lateralization in the rice weevil, Sitophilus oryzae (L.) (Coleoptera: Curculionidae). J. Stored Prod. Res. 2016, 69, 106–112. [Google Scholar] [CrossRef]
- Boukouvala, M.; Romano, D.; Kavallieratos, N.G.; Stefanini, C.; Canale, A.; Benelli, G. Asymmetric courtship boosts male mating success in the red flour beetle, Tribolium castaneum (Herbst) (Coleoptera: Tenebrionidae). J. Stored Prod. Res. 2019, 81, 1–6. [Google Scholar] [CrossRef]
- Benelli, G. Mating behavior of the West Nile virus vector Culex pipiens—Role of behavioral asymmetries. Acta Trop. 2018, 179, 88–95. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D. Does indirect mating trophallaxis boost male mating success and female egg load in Mediterranean fruit flies? J. Pest Sci. 2018, 91, 181–188. [Google Scholar] [CrossRef]
- Chopra, N.P. Studies on the genus Halys Hemiptera: Pentatomidae. Orient. Insects 1974, 8, 473–479. [Google Scholar] [CrossRef]
- Schmutterer, H. Properties and potential of natural pesticides from the neem tree, Azadirachta indica. Annu. Rev. Entomol. 1990, 35, 271–297. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V. A review on medicinal properties of neem (Azadirachta indica). J. Pharm. Innov. 2018, 7, 648–650. [Google Scholar]
- Benelli, G.; Murugan, K.; Panneerselvam, C.; Madhiyazhagan, P.; Conti, B.; Nicoletti, M. Old ingredients for a new recipe? Neem cake, a low-cost botanical by-product in the fight against mosquito-borne diseases. Parasitol. Res. 2015, 114, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Canale, A.; Toniolo, C.; Higuchi, A.; Murugan, K.; Pavela, R.; Nicoletti, M. Neem (Azadirachta indica): Towards the ideal insecticide? Nat. Prod. Res. 2017, 31, 369–386. [Google Scholar] [CrossRef]
- Panizzi, A.R.; McPherson, J.E.; James, D.G.; Javahery, M.; McPherson, R.M. Stink bugs (Pentatomidae). Heteroptera of Econ. Import. 2000, 828, 421–474. [Google Scholar]
- Cokl, A.; Borges, M. Stink bug classification, phylogeny, biology and reproductive behavior. In Stinkbugs; CRC Press: Boca Raton, FL, USA, 2017; pp. 21–50. [Google Scholar]
- Pezzini, D.T.; Nystrom Santacruz, E.C.; Koch, R.L. Predation and Parasitism of Halyomorpha halys (Hemiptera: Pentatomidae) Eggs in Minnesota. Environ. Entomol. 2018, 47, 812–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simberloff, D. Everything you want to know about stink bugs. Biol. Invasions 2018, 20, 2691–2693. [Google Scholar] [CrossRef]
- Carvajal, M.A.; Alaniz, A.J.; Núñez-Hidalgo, I.; González-Césped, C. Spatial global assessment of the pest Bagrada hilaris (Burmeister) (Heteroptera: Pentatomidae): Current and future scenarios. Pest Manag. Sci. 2018, 75, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Havaskary, M.; Farshbaf Pour-Abad, R.; Kazemi, M.H.; Rafeii, A. A contribution to the stink bugs from Khodafarin, NW Iran (Heteroptera: Pentatomidae: Pentatominae). Munis Entomol. Zool. 2012, 7, 607–616. [Google Scholar]
- Dhiman, S.C.; Yadav, Y.K. Studies on parasitoids of Halys dentatus Fabr. (Heteroptera: Pentatomidae) and their bio-control efficacy. Ann. Plant Prot. Sci. 2004, 12, 263–266. [Google Scholar]
- Gamboa, G.; Alcock, J. The Mating Behavior of Brochymen quadrapustulata (Fabricius). Psyche J. Entomol. 1973, 80, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Kawada, H.; Kitamura, C. The reproductive behavior of the brown marmorated stink bug, Halyomorpha mista Uhler (Heteroptera: Pentatomidae) I. Observation of mating behavior and multiple copulation. Appl. Entomol. Zool. 1983, 18, 234–242. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Millar, J.G. Reproductive behavior of Thyanta pallidovirens (Heteroptera: Pentatomidae). Ann. Entomol. Soc. 1997, 90, 380–388. [Google Scholar] [CrossRef]
- Benelli, G.; Romano, D. Looking for the right mate—What do we really know on the courtship and mating of Lucilia sericata (Meigen)? Acta Trop. 2019, 189, 145–153. [Google Scholar] [CrossRef]
- Fahrbach, S.E.; Mesce, K.A. “Neuroethoendocrinology”: Integration of field and laboratory studies in insect neuroendocrinology. Horm. Behav. 2005, 48, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Meregalli, M.; Canale, A. Field observations on the mating behavior of Aclees sp. cf. foveatus Voss (Coleoptera: Curculionidae), an exotic pest noxious to fig orchards. J. Insect Behav. 2014, 27, 419–427. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry; WH Freeman and Company: San Francisco, CA, USA, 1981; p. 859. [Google Scholar]
- Ajelli, M. Modeling mosquito-borne diseases in complex urban environments. Acta Trop. 2017, 176, 332–334. [Google Scholar] [CrossRef] [PubMed]
- Scavuzzo, J.M.; Trucco, F.; Espinosa, M.; Tauro, C.B.; Abril, M.; Scavuzzo, C.M.; Frery, A.C. Modeling Dengue vector population using remotely sensed data and machine learning. Acta Trop. 2018, 185, 167–175. [Google Scholar] [CrossRef] [Green Version]
- Pavlovčič, P.; Čokl, A. Songs of Holcostethus strictus (Fabricius): A different repertoire among landbugs (Heteroptera: Pentatomidae). Behav. Process. 2001, 53, 65–73. [Google Scholar] [CrossRef]
- Cousillas, H.; Henry, L.; George, I.; Marchesseau, S.; Hausberger, M. Lateralization of social signal brain processing correlates with the degree of social integration in a songbird. Sci. Rep. 2020, 10, 14093. [Google Scholar] [CrossRef]
- Rogers, L.J.; Rigosi, E.; Frasnelli, E.; Vallortigara, G. A right antenna for social behaviour in honeybees. Sci. Rep. 2013, 3, 2045. [Google Scholar] [CrossRef] [Green Version]
- Waite, L.; Frasnelli, E. Visuo-motor biases in buff-tailed bumblebees (Bombus terrestris). Laterality 2021, 26, 55–70. [Google Scholar] [CrossRef]
- Kamimura, Y. Right-handed penises of the earwig Labidura riparia (Insecta, Dermaptera, Labiduridae): Evolutionary relationships between structural and behavioral asymmetries. J. Morphol. 2006, 267, 1381–1389. [Google Scholar] [CrossRef]
- Kamimura, Y.; Iwase, R. Evolutionary genetics of genital size and lateral asymmetry in the earwig Euborellia plebeja (Dermaptera: Anisolabididae). Biol. J. Linn. Soc. 2010, 101, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Kamimura, Y.; Matsumura, Y.; Yang, C.C.S.; Gorb, S.N. Random or handedness? Use of laterally paired penises in Nala earwigs (Insecta: Dermaptera: Labiduridae). Biol. J. Linn. Soc. 2021, 134, 716–731. [Google Scholar] [CrossRef]
- Romano, D.; Benelli, G.; Kavallieratos, N.G.; Athanassiou, C.G.; Canale, A.; Stefanini, C. Beetle-robot hybrid interaction: Sex, lateralization and mating experience modulate behavioural responses to robotic cues in the larger grain borer Prostephanus truncatus (Horn). Biol. Cybern. 2020, 114, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Ghirlanda, S.; Vallortigara, G. The evolution of brain lateralization: A game-theoretical analysis of population structure. Proc. R. Soc. B 2004, 271, 853. [Google Scholar] [CrossRef] [Green Version]
- Ghirlanda, S.; Frasnelli, E.; Vallortigara, G. Intraspecific competition and coordination in the evolution of lateralization. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2009, 364, 861–866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benelli, G.; Romano, D.; Rocchigiani, G.; Caselli, A.; Mancianti, F.; Canale, A.; Stefanini, C. Behavioral asymmetries in ticks–Lateralized questing of Ixodes ricinus to a mechatronic apparatus delivering host-borne cues. Acta Trop. 2018, 178, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Frasnelli, E.; Vallortigara, G. Individual-level and population-level lateralization: Two sides of the same coin. Symmetry 2018, 10, 739. [Google Scholar] [CrossRef]
- Manns, M. Laterality for the next decade: Costs and benefits of neuronal asymmetries—Putting lateralization in an evolutionary context. Laterality 2021, 1–4. [Google Scholar] [CrossRef]
- Ventolini, N.; Ferrero, E.A.; Sponza, S.; Della Chiesa, A.; Zucca, P.; Vallortigara, G. Laterality in the wild: Preferential hemifield use during predatory and sexual behaviour in the black-winged stilt. Anim. Behav. 2005, 69, 1077–1084. [Google Scholar] [CrossRef]
- Gross, M.R.; Suk, H.Y.; Robertson, C.T. Courtship and genetic quality: Asymmetric males show their best side. Proc. R. Soc. B Biol. Sci. 2007, 274, 2115–2122. [Google Scholar] [CrossRef] [Green Version]
- Hews, D.K.; Castellano, M.; Hara, E. Aggression in females is also lateralized: Left-eye bias during aggressive courtship rejection in lizards. Anim. Behav. 2004, 68, 1201–1207. [Google Scholar] [CrossRef]
- Sion, G. Foot-preference underlies bite-scar asymmetry in the gecko Ptyodactylus guttatus. Laterality 2018, 23, 129–151. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Behavioral Display | Left-Biased Approaches | Right-Biased Approaches | Tested Bugs (n., Left + Right-Biased Bugs) |
|---|---|---|---|
| antennal tapping duration (s) | 20.72 ± 1.01 a | 21.42 ± 1.58 a | 22 + 7 = 29 |
| foreleg palpation duration (s) | 15.90 ± 1.05 a | 16 ± 2.60 a | 22 + 7 = 29 |
| duration of male’s antennal contact with the backside of the female (s) | 22.77 ± 0.96 a | 21 ± 1.92 a | 22 + 7 = 29 |
| copulation attempts (n) | 9.77 ± 1.46 a | 13.85 ± 2.12 a | 22 + 7 = 29 |
| copula duration (min) | 76.63 ± 9.65 a | 56 ± 20.03 a | 17 + 4 = 21 |
| Behavioral Display | Turning 180° Left | Turning 180° Right | Backside Mounting | Tested Bugs (n., Left + Right-Biased + Back Mouting Bugs) |
|---|---|---|---|---|
| antennal tapping duration (s) | 19.9 ± 1.38 a | 21.70 ± 1.15 a | 19 + 3 a | 17 + 10 + 2 = 29 |
| foreleg palpation duration (s) | 15.3 ± 2.02 a | 16.05 ± 1.11 a | 18 ± 6 a | 17 + 10 + 2 = 29 |
| duration of male’s antennal contact with the backside of the female (s) | 20.1 ± 0.75 a | 23.88 ± 1.10 a | 20.5 ± 4.5 a | 17 + 10 + 2 = 29 |
| copulation attempts (n) | 16.8 ± 2.06 a | 6.64 ± 0.77 b | 15.5 ± 3.5 a | 17 + 10 + 2 = 29 |
| copula duration (min) | 55.3 ± 15.34 a | 84.58 ± 10.52 a | 43.5 ± 43.5 a | 14 + 6 + 1 = 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romano, D.; Benelli, G.; Stefanini, C. Lateralization of Courtship Traits Impacts Pentatomid Male Mating Success—Evidence from Field Observations. Insects 2022, 13, 172. https://doi.org/10.3390/insects13020172
Romano D, Benelli G, Stefanini C. Lateralization of Courtship Traits Impacts Pentatomid Male Mating Success—Evidence from Field Observations. Insects. 2022; 13(2):172. https://doi.org/10.3390/insects13020172
Chicago/Turabian StyleRomano, Donato, Giovanni Benelli, and Cesare Stefanini. 2022. "Lateralization of Courtship Traits Impacts Pentatomid Male Mating Success—Evidence from Field Observations" Insects 13, no. 2: 172. https://doi.org/10.3390/insects13020172