
Performance of Fall Armyworm Preimaginal Development on Cultivars of Tropical Grass Forages
 , ,
, ,  , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Forage Grasses
2.2. Bioassays
2.3. Statistical Analyses
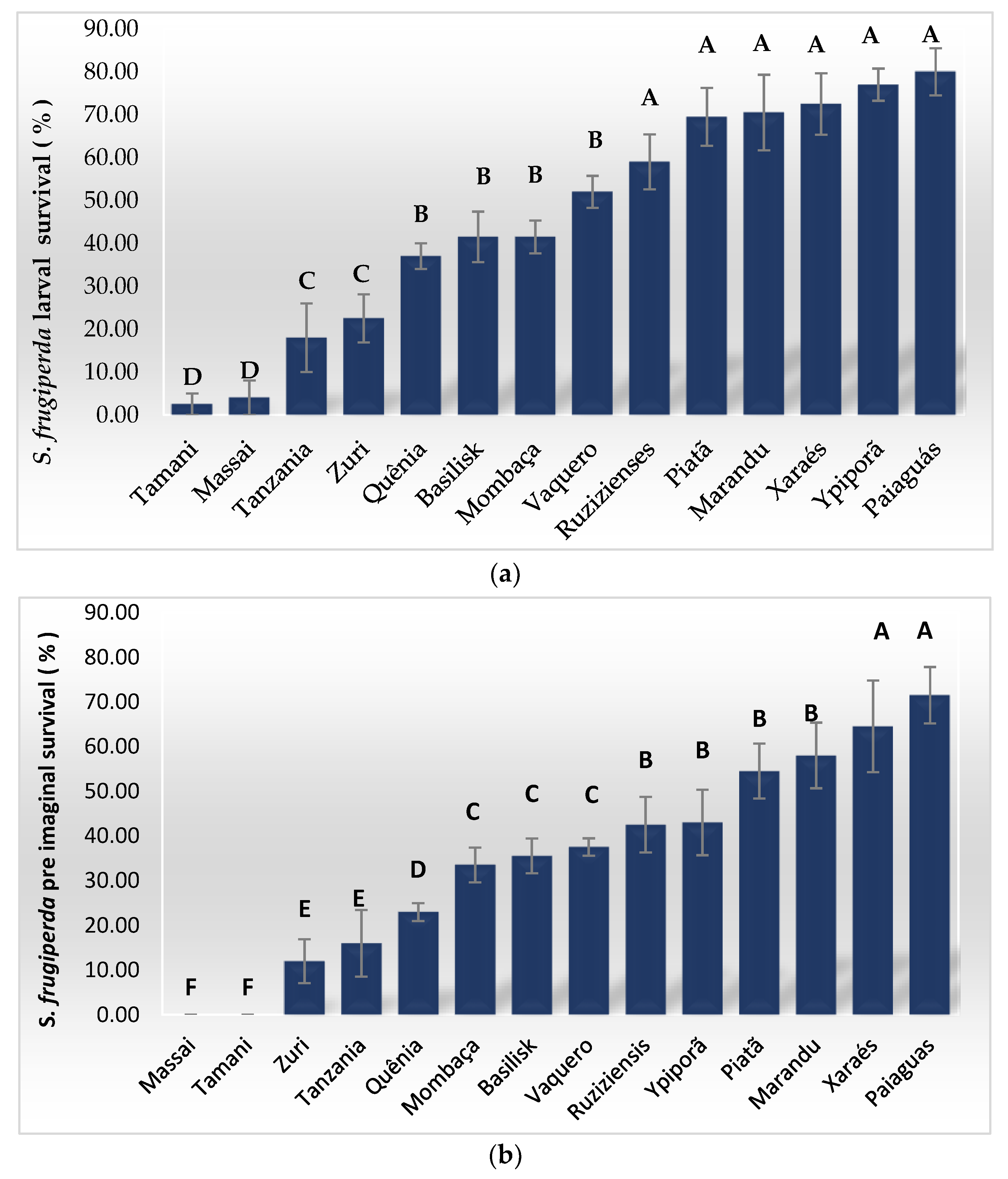
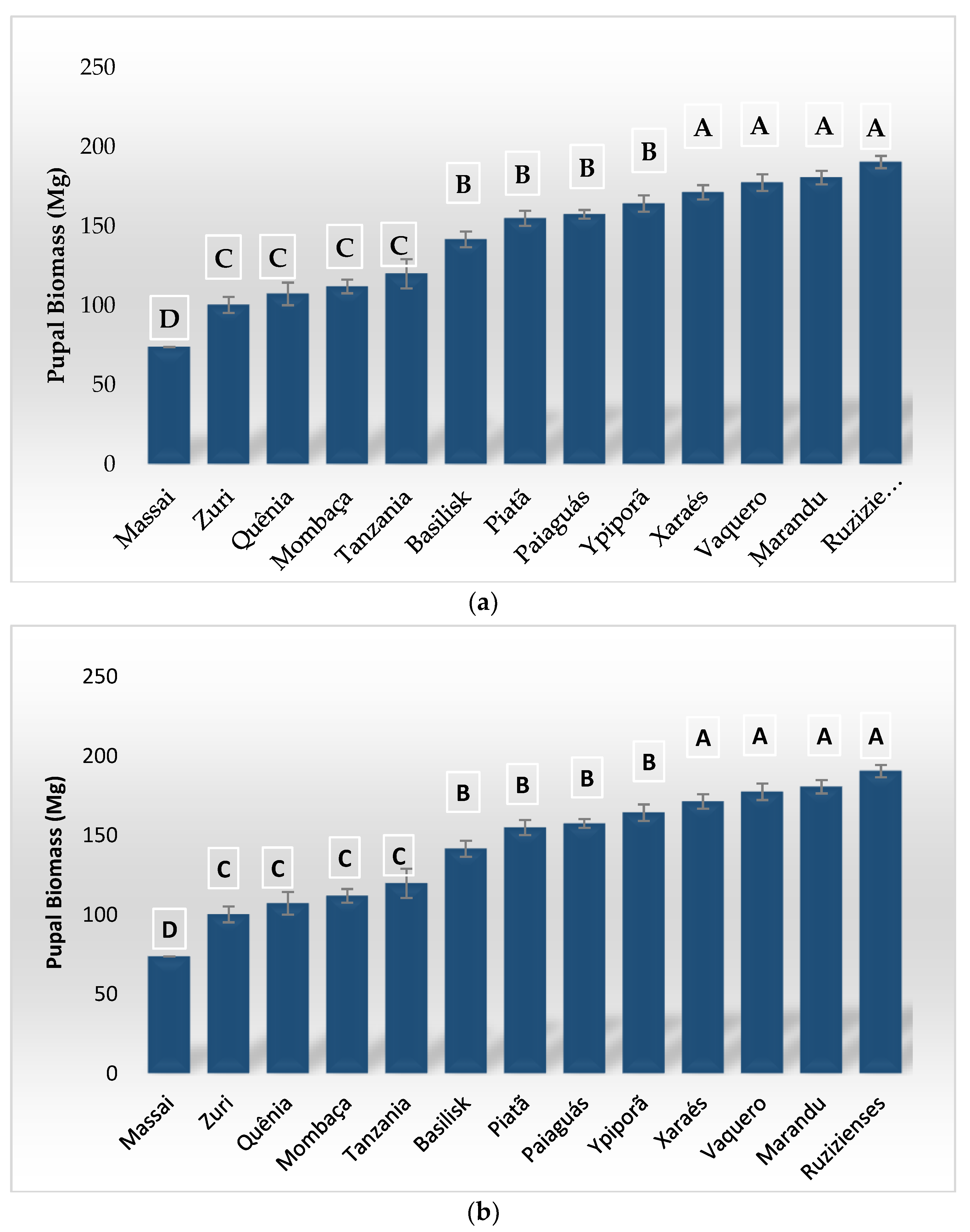
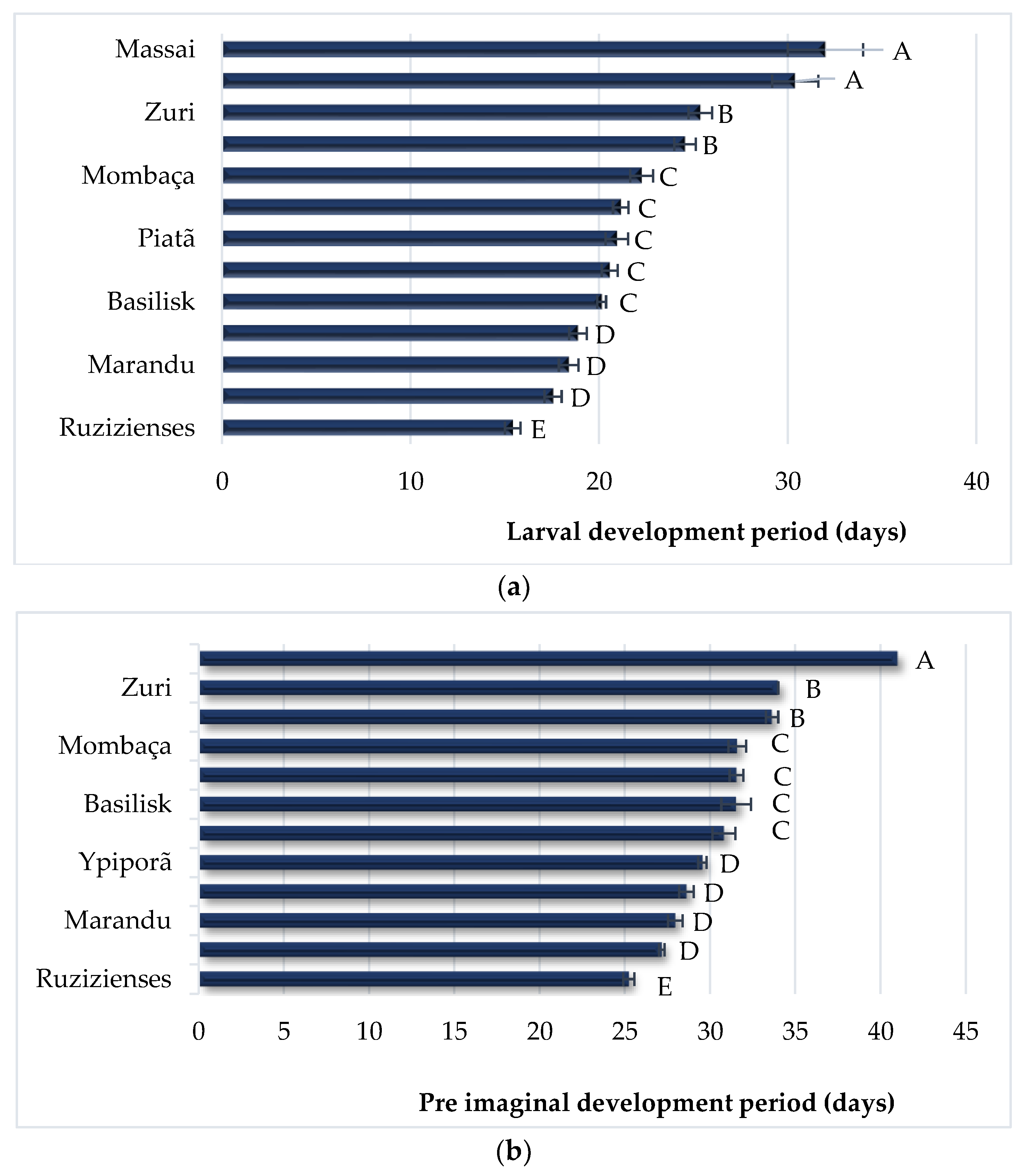
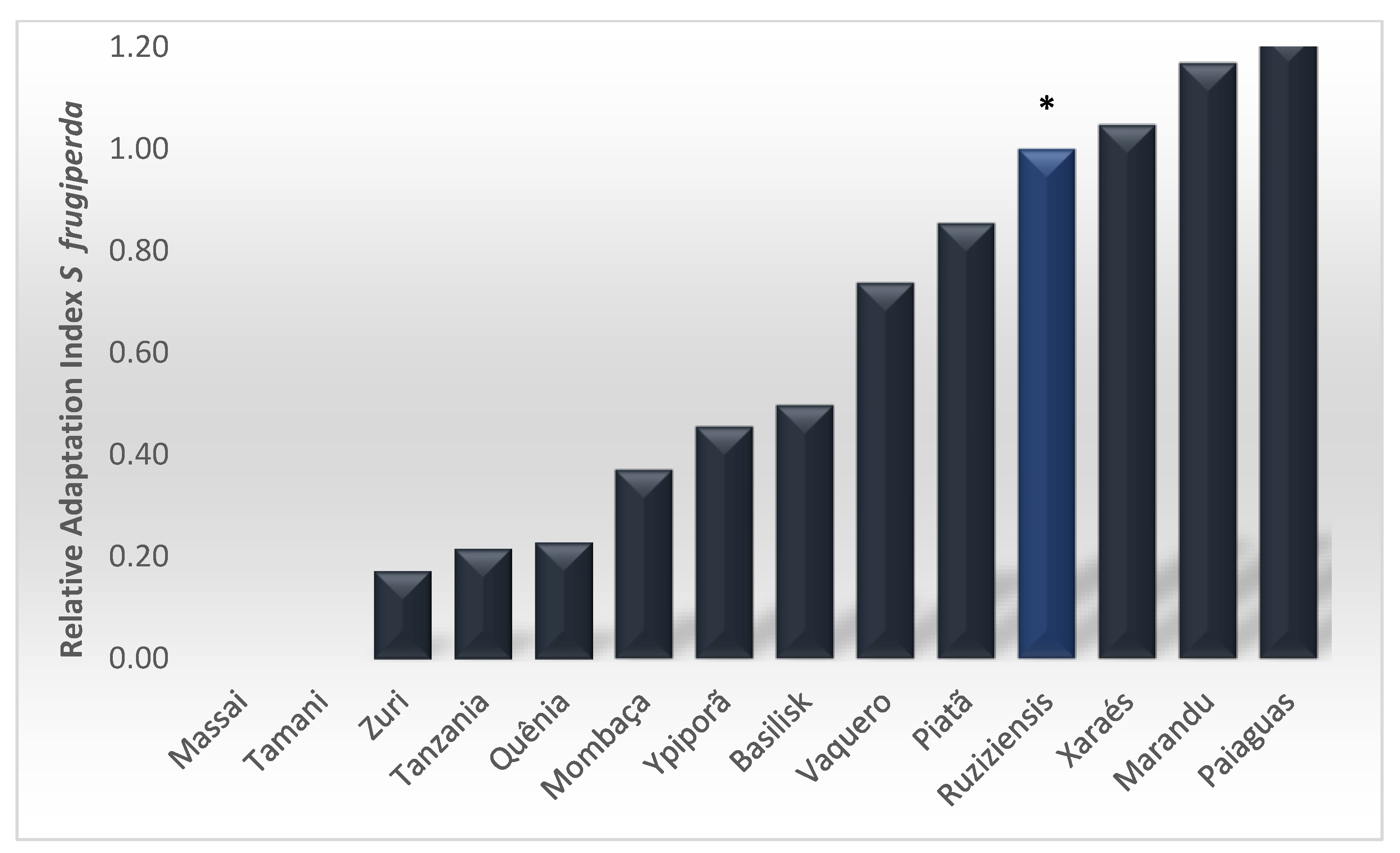
3. Results
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Altaf, N.; Idrees, A.; Ullah, M.I.; Arshad, M.; Afzal, A.; Afzal, M.; Rizwan, M.; Li, J. Biotic Potential Induced by Different Host Plants in the Fall Armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 921. [Google Scholar] [CrossRef] [PubMed]
- Boregas, K.G.B.; Mendes, S.M.; Waquil, J.M.; Fernandes, G.W. Estádio de adaptação de Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) em hospedeiros alternativos. Bragantia 2013, 72, 61–70. (In Portuguese) [Google Scholar] [CrossRef] [Green Version]
- Sá, V.G.M.; Fonseca, B.V.C.; Boregas, K.G.B.; Waquil, J.M. Sobrevivência e desenvolvimento larval de Spodoptera frugiperda (J. E. Smith, 1797) (Lepidoptera: Noctuidae) em hospedeiros alternativos. Neotrop. Entomol. 2009, 38, 108–115. (In Portuguese) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Moraes, T.; Da Silva, A.F.; Leite, N.A.; Karam, D.; Mendes, S.M. Survival and development of fall armyworm (Lepidoptera: Noctuidae) in weeds during the off-season. Fla Entomol. 2020, 103, 288–292. [Google Scholar] [CrossRef]
- Li, D.-D.; Li, J.-Y.; Hu, Z.-Q.; Liu, T.-X.; Zhang, S.-Z. Fall Armyworm Gut Bacterial Diversity Associated with Different Developmental Stages, Environmental Habitats, and Diets. Insects 2022, 13, 762. [Google Scholar] [CrossRef] [PubMed]
- Gontijo, N.M.M.; Silva, R.V.; Borghi, E.; De Resende, A.V.; Melo, C.J. Alternativas de Integração Lavoura-Pecuária Para Produção de Forragens e Recuperação de Pastagens; Estudo de caso da Fazenda São Pedro: Unaí, Brazil; Embrapa Milho e Sorgo: Sete Lagoas, Brazil, 2018. (In Portuguese) [Google Scholar]
- Ribeiro, L.P.; Klock, A.L.S.; Nesi, C.N.; Luczkievicz, F.R.G.; Travi, M.R.L.; Rech, A.F. Adaptability and comparative biology of fall armyworm on maize and perennial forage species and relation with chemical-bromatological composition. Neotrop. Entomol. 2020, 49, 758–767. [Google Scholar] [CrossRef] [PubMed]
- Dias, A.S.; Marucci, R.C.; Mendes, S.M.; Moreira, S.G.; Araújo, O.G.; Dos Santos, C.A.; Barbosa, T.A. Bioecology of Spodoptera frugiperda (JE Smith, 1757) in different cover crops. Biosci. J. 2016, 32, 2. [Google Scholar] [CrossRef] [Green Version]
- Lv, D.; Liu, X.; Dong, Y.; Yan, Z.; Zhang, X.; Wang, P.; Yuan, X.; Li, Y. Comparison of Gut Bacterial Communities of Fall Armyworm (Spodoptera frugiperda) Reared on Different Host Plants. Int. J. Mol. Sci. 2021, 22, 11266. [Google Scholar] [CrossRef] [PubMed]
- Parra, J.R.P. Técnicas de Criação de Insetos Para Programas de Controle Biológico, 3rd ed.; FEALQ: Piracicaba, Brazil, 1996; 137p. [Google Scholar]
- Barrios, S.C.L.; Carromeu, C.; Matsubara, E.T.; Crivellaro, L.L.; Silva, C.B.; Santos, M.F.; Jank, L. Pasto Certo—Versão 2.0®: Aplicativo Para Dispositivos Móveis e Desktop Sobre Forrageiras Tropicais; Gado de Corte: Campo Grande, Brazil, 2014. (In Portuguese) [Google Scholar]
- Jank, L.; Barrios, S.C.; Do Valle, C.B.; Simeão, R.M.; Alves, G.F. The value of improved pastures to Brazilian beef production. Crop. Pasture Sci. 2014, 65, 1132–1137. [Google Scholar] [CrossRef]
- Brunetti, H.B.; Cavalcanti, P.P.; Dias, C.T.S.; Pezzopane, J.R.M.; Santos, P.M. Climate risk and seasonal forage production of Marandu palisadegrass in Brazil. Anais da Academia Brasileira de Ciências 2020, 92, e20190046. [Google Scholar] [CrossRef] [PubMed]
- Landau, E.C.; Simeão, R.M.; Matos-Neto, F.C. Evolução da área ocupada por pastagens. In Dinâmica Da Produção Agropecuária e da Paisagem Natural no Brasil nas Últimas dÉcadas: Produtos de Origem Animal e da Silvicultura, 3rd ed.; Landau, E., da Silva, G.A., Moura, L., Hirsch, A., Guimaraes, D., Eds.; Embrapa: Brasília, Brazil, 2020; pp. 1555–1578. (In Portuguese) [Google Scholar]
- Pencoe, N.L.; Martin, P.B. Fall armyworm (Lepidoptera: Noctuidae) larval development and adult fecundity on five grass hosts. Environ. Entomol. 1982, 11, 720–724. [Google Scholar] [CrossRef]
- Jacques, F.L.; Degrande, P.E.; Gauer, E.; Malaquias, J.B.; Scoton, A.M.N. Intercropped Bt and non-Bt corn with ruzigrass (Urochloa ruziziensis) as a tool to resistance management of Spodoptera frugiperda (JE Smith, 1797) (Lepidoptera: Noctuidae). Pest Manag. Sci. 2021, 77, 3372–3381. [Google Scholar] [CrossRef] [PubMed]
- Pannuti, L.E.R.; Paula-Moraes, S.V.; Hunt, T.E.; Baldin, E.L.L.; Dana, L.; Malaquias, J.V. Plant-to-plant movement of Striacosta albicosta (Lepidoptera: Noctuidae) and Spodoptera frugiperda (Lepidoptera: Noctuidae) in maize (Zea mays). J. Econ. Entomol. 2016, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flausino, B.F.; Machado, C.F.; Silva, J.H.C.; Ronchi, C.P.; Pimentel, M.A.G.; Gontijo, L.M. Intercropping maize with brachiaria can be a double-edged sword strategy. Pest Manag. Sci. 2022. [Google Scholar] [CrossRef] [PubMed]
- Braman, S.K.; Duncan, R.R. Evaluation of turfgrass selections for resistance to fall armyworms (Lepidoptera: Noctuidae). Hort. Sci. 2000, 35, 1268–1270. [Google Scholar] [CrossRef] [Green Version]
- Galdos, M.; Brown, E.; Rosolem, C.; Pires, L.; Hallett, P.; Mooney, S.J.S.R. Brachiaria species influence nitrate transport in soil by modifying soil structure with their root system. Sci. Rep. 2020, 10, 5072. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scientific Name | Cultivar | Common Name |
|---|---|---|
| Brachiaria brizantha (Hochst) Stapf | Marandu | Signal grass |
| Panicum maximum Jacq. | Tanzânia | Guinea grass |
| Brachiaria decumbens Stapf | Basilisk | Signal grass |
| Brachiaria brizantha | BRS Paiaguás | Signal grass |
| Brachiaria brizantha | BRS Xaraés | Signal grass |
| Brachiaria spp. | BRS Ipyporã | hybrid |
| Brachiaria brizantha | BRS Piatã | Signal grass |
| Brachiaria ruziziensis Germain et Evrard | Ruziziensis | Congo signal grass |
| Panicum maximum | Mombaça | Guinea grass |
| Panicum maximum | Massai | Guinea grass |
| Panicum maximum | BRS Zuri | Guinea grass |
| Panicum maximum | BRS Tamani | Guinea grass |
| Panicum maximum | BRS Quênia | Guinea grass |
| Cynodon dactylon | Vaquero | Bermudagrass |
| Source: Embrapa |
| Host Plant | Initial Survival (%) | Larval Survival (%) | Biomassa Larval (mg) | Pupa Biomass (mg) | Larval Period (days) | Youth Development Period (days) |
|---|---|---|---|---|---|---|
| Basilisk | 4.5 ± 5.9 c | 35.5 ± 3.9 c | 66.2 ± 4.6 c | 141.6 ± 4.9 b | 20.1 ± 0.2 c | 31.5 ± 0.9 c |
| Brizantha | 70.5 ± 8.8 a | 58.0 ± 7.4 b | 84.5 ± 5.1 b | 180.6 ± 4.3 a | 18.4 ± 0.5 d | 28.0 ± 0.4 d |
| Massai | 4.0 ± 4.0 e | 0.0 ± 0.0 f | 10.4 ± 0.9 e | 73.8 ± 0.0 d | 32.0 ± 2.0 a | 0.0 ± 0.0 f |
| Mombaça | 41.5 ± 3.8 c | 33.5 ± 3.8 c | 39.2 ± 5.7 d | 112.0 ± 4.3 c | 22.3 ± 0.6 c | 31.6 ± 0.5 c |
| BRS Paiaguás | 80.0 ± 5.5 a | 71.5 ± 6.3 a | 55.5 ± 5.1 c | 157.5 ± 2.8 b | 18.9 ± 0.5 d | 28.6 ± 0.4 d |
| BRS Piatã | 69.5 ± 6.7 a | 54.5 ± 6.1 b | 48.9 ± 6.3 c | 154.9 ± 4.7 b | 20.9 ± 0.6 c | 30.8 ± 0.7 c |
| BRS Quênia | 37.0 ± 3.0 c | 23.0 ± 2.0 d | 41.1 ± 2.7 d | 107.3 ± 7.1 c | 24.6 ± 0.6 b | 33.6 ± 0.4 b |
| Ruziziensis | 59.0 ± 6.4 a | 42.5 ± 6.2 b | 204.9 ± 11.0 a | 190.5 ± 3.9 a | 15.4 ± 0.4 e | 25.2 ± 0.3 e |
| BRS Tamani | 2.5 ± 2.5 e | 0.0 ± 0.0 f | 24.9 ± 3.1 e | 0.0 ± 0.0 f | 0.0 ± 0.0 f | 0.0 ± 0.0 f |
| Tanzânia | 18.0 ± 8.0 d | 16.0 ± 7.5 e | 17.3 ± 2.3 e | 120.0 ± 9.3 c | 30.4 ± 1.2 a | 41.0 ± 1.5 a |
| Vaquero | 52.0 ± 3.7 b | 37.5 ± 1.9 c | 79.2 ± 4.5 b | 177.5 ± 5.2 a | 17.6 ± 0.5 d | 27.2 ± 0.2 d |
| BRS Xaraés | 72.5 ± 7.2 a | 64.5 ± 10.3 a | 26.6 ± 2.7 e | 171.3 ± 4.5 a | 21.1 ± 0.4 c | 31.6 ± 0.4 c |
| BRS Ipyporã | 77.0 ± 3.7 a | 43.0 ± 7.4 b | 52.2 ± 3.0 c | 164.3 ± 5.2 b | 20.6 ± 0.4 c | 29.6 ± 0.3 d |
| BRS Zuri | 22.5 ± 5.6 d | 12.0 ± 4.9 e | 13.2 ± 1.6 e | 100.4 ± 5.0 c | 25.4 ± 0.6 b | 34.0 ± 0.0 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
dos Santos, M.V.C.; Nascimento, P.T.; Simeone, M.L.; Lima, P.F.; Simeão, R.M.; Auad, A.; Oliveira, I.; Mendes, S. Performance of Fall Armyworm Preimaginal Development on Cultivars of Tropical Grass Forages. Insects 2022, 13, 1139. https://doi.org/10.3390/insects13121139
dos Santos MVC, Nascimento PT, Simeone ML, Lima PF, Simeão RM, Auad A, Oliveira I, Mendes S. Performance of Fall Armyworm Preimaginal Development on Cultivars of Tropical Grass Forages. Insects. 2022; 13(12):1139. https://doi.org/10.3390/insects13121139
Chicago/Turabian Styledos Santos, Marcos V. C., Priscilla T. Nascimento, Maria L. Simeone, Patrick F. Lima, Rosangela M. Simeão, Alexander Auad, Ivênio Oliveira, and Simone Mendes. 2022. "Performance of Fall Armyworm Preimaginal Development on Cultivars of Tropical Grass Forages" Insects 13, no. 12: 1139. https://doi.org/10.3390/insects13121139