
Insecticidal Activity of a Component, (-)-4-Terpineol, Isolated from the Essential Oil of Artemisia lavandulaefolia DC. against Plutella xylostella (L.)
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Plant Materials and Essential Oil Extraction
2.3. Authentic Compounds
2.4. Identification of Compounds
2.5. Toxicity Bioassays of Bioactive Compounds
2.5.1. Contact Bioassays
2.5.2. Fumigant Bioassays
2.6. Median Lethal Dose (LD50) and Concentration (LC50)
2.7. Median Knockdown Time (KT50) of (-)-4-Terpineol
2.8. Enzyme Assays
2.8.1. Enzyme Preparation Method
2.8.2. Protein Assay
2.8.3. GST Activity Assay
2.8.4. CAT Activity Assay
2.8.5. AChE Activity Assay
2.8.6. Na+/K+-ATPase
2.9. Statistical Analysis
3. Results
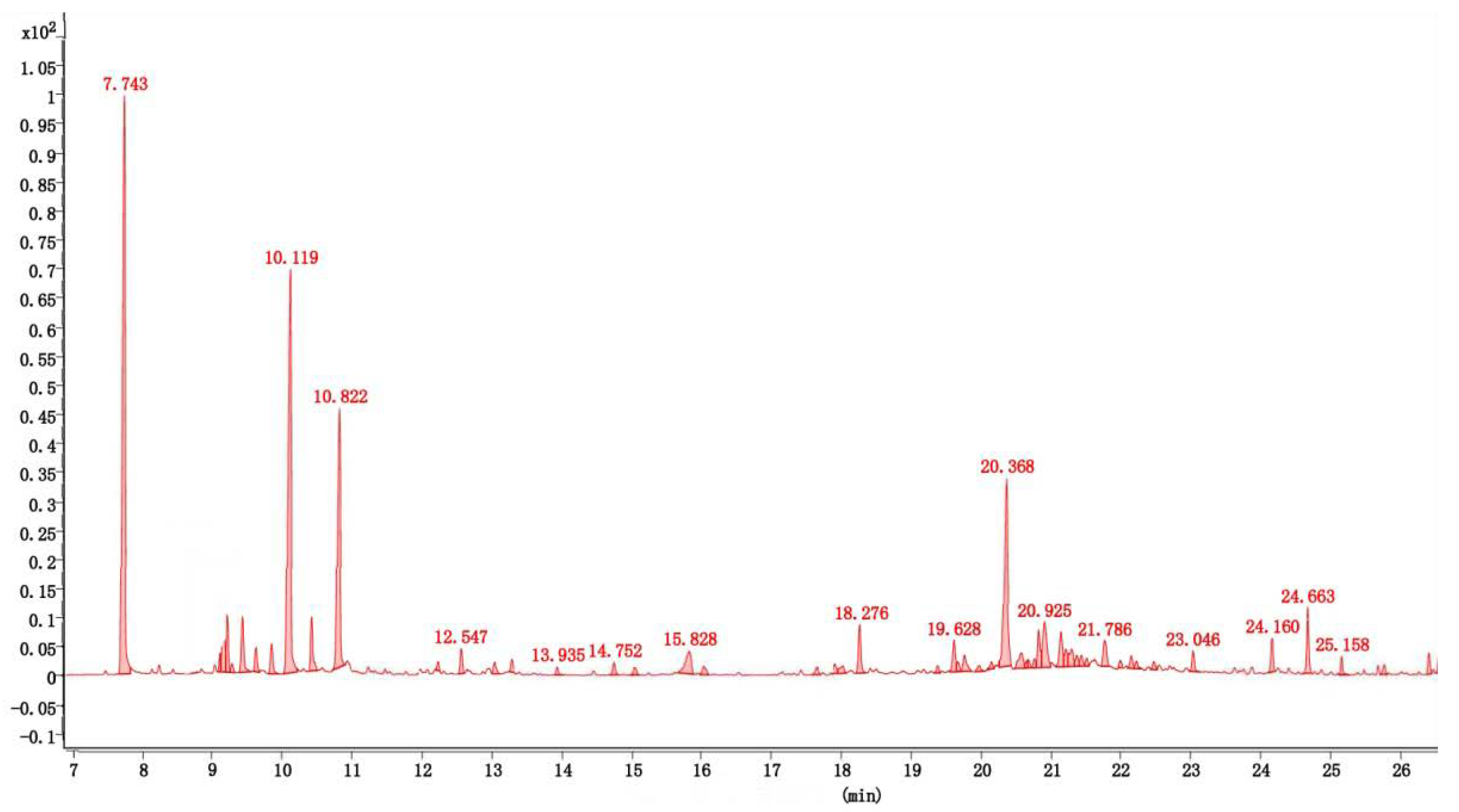
3.1. Composition of the Essential Oil
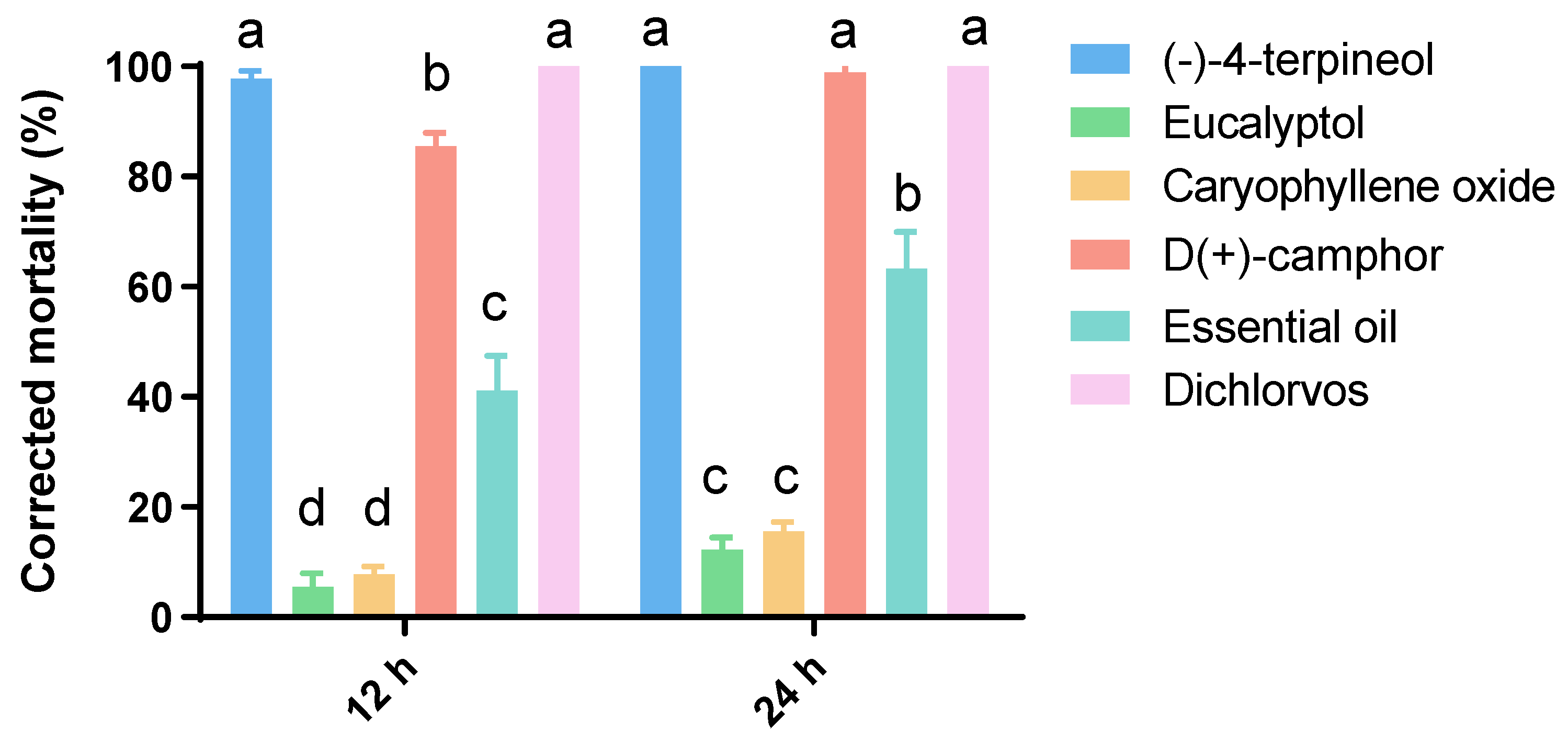
3.2. Insecticidal Assays
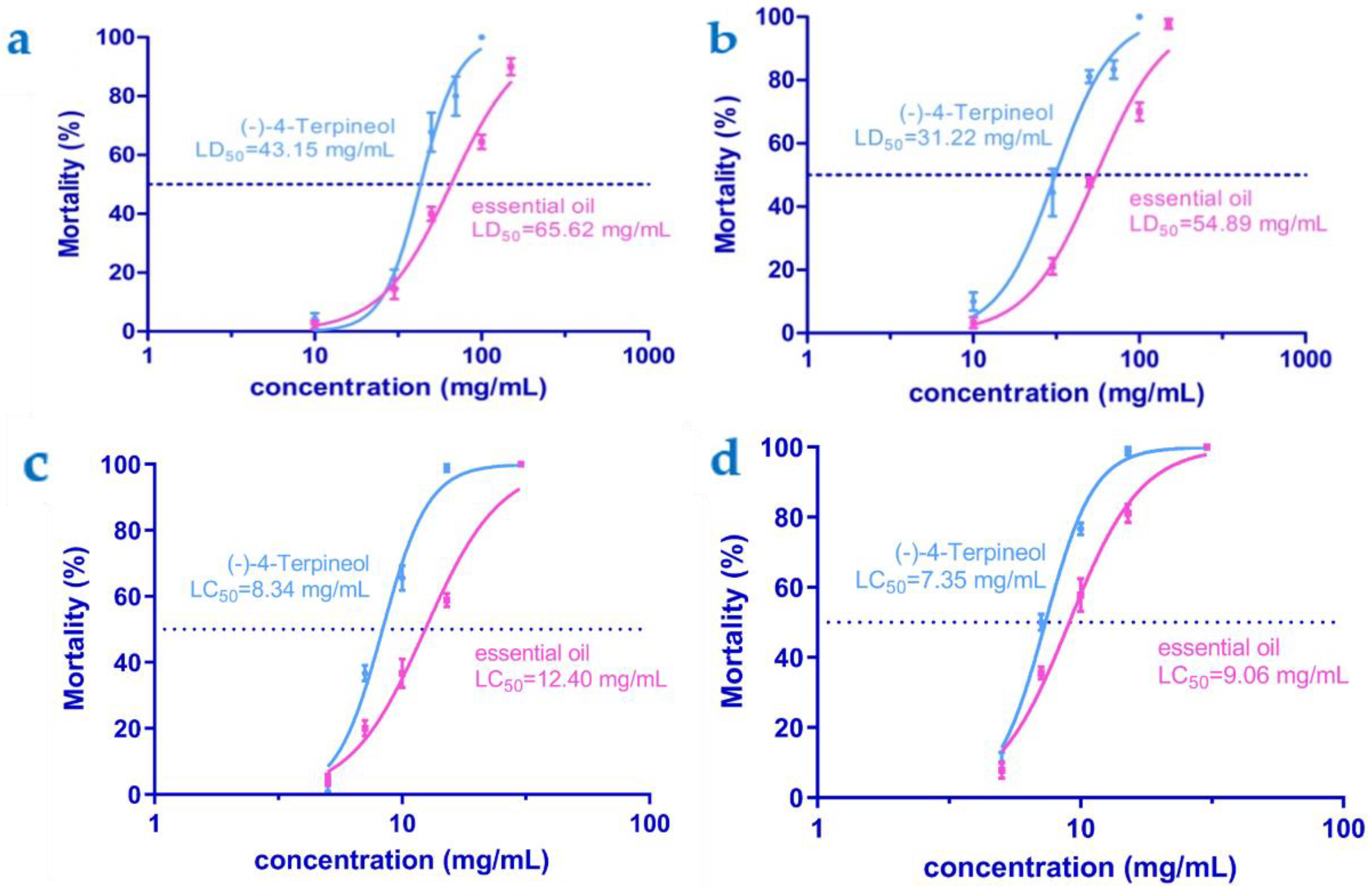
3.3. Toxicity Bioassays
3.4. Median Knockdown Time (KT50) of (-)-4-Terpineol
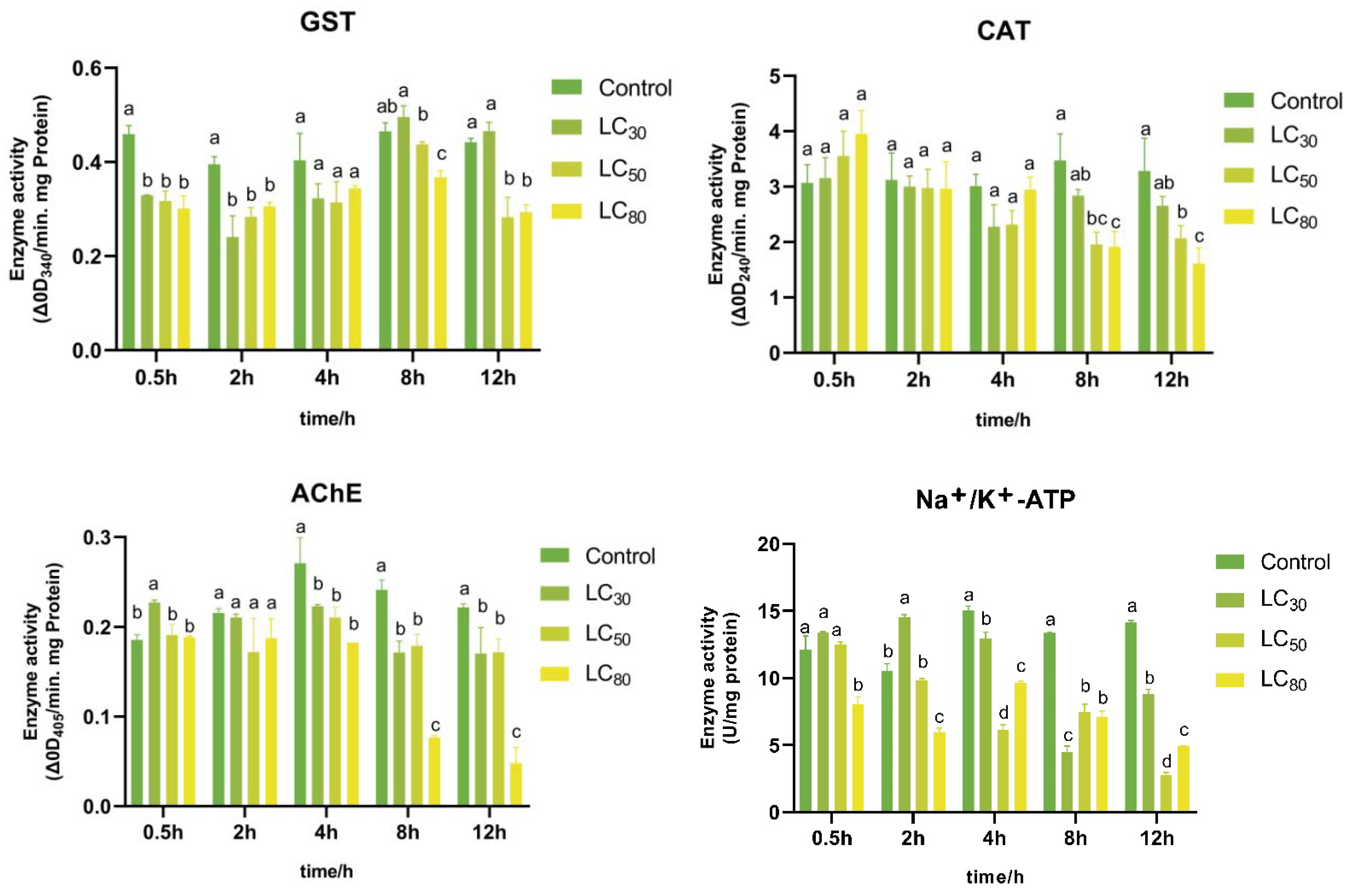
3.5. Enzyme Activities Assays
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Z.Y.; Xia, F.; Liu, S.S.; You, M.S.; Furlong, M.J. Biology, ecology, and management of the diamondback moth in China. Annu. Rev. Entomol. 2016, 61, 277–296. [Google Scholar] [CrossRef] [PubMed]
- Furlong, M.; Wright, D.; Dosdall, L. Diamondback moth ecology and management: Problems, progress, and prospects. Annu. Rev. Entomol. 2013, 58, 517–541. [Google Scholar] [CrossRef] [PubMed]
- Tharamak, S.; Yooboon, T.; Pengsook, A.; Ratwatthananon, A.; Pluempanupat, W. Synthesis of thymyl esters and their insecticidal activity against Spodoptera litura (Lepidoptera: Noctuidae). Pest Manag. Sci. 2019, 76, 928–935. [Google Scholar] [CrossRef] [PubMed]
- Charleston, D.S.; Kfir, R. The possibility of using Indian mustard, Brassica juncea, as a trap crop for the diamondback moth, Plutella xylostella, in South Africa. Crops Prot. 2000, 19, 455–460. [Google Scholar] [CrossRef]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef] [Green Version]
- Kumrungsee, N.; Pluempanupat, W.; Koul, O.; Bullangpoti, V. Toxicity of essential oil compounds against diamondback moth, Plutella xylostella, and their impact on detoxification enzyme activities. J. Pest Sci. 2014, 87, 721–729. [Google Scholar] [CrossRef]
- López, M.; Jordán, M.; Pascual-Villalobos, M.J. Toxic compounds in essential oils of coriander, caraway and basil active against stored rice pests. J. Stored Prod. Res. 2008, 44, 273–278. [Google Scholar] [CrossRef]
- Park, I.K.; Lee, S.G.; Choi, D.H.; Park, J.D.; Ahn, Y.J. Insecticidal activities of constituents identified in the essential oil from leaves of Chamaecyparis obtusa against Callosobruchus chinensis (L.) and Sitophilus oryzae (L.). J. Stored Prod. Res. 2003, 39, 375–384. [Google Scholar] [CrossRef]
- Radhika, S.; Kalhari, S.K.; Weerasinghe, S.I. Insecticidal activity of essential oils of Ceylon Cinnamomum and Cymbopogon species against Musca domestica. J. Essent. Oil Res. 2006, 18, 352–354. [Google Scholar] [CrossRef]
- Bossou, A.D.; Ahoussi, E.; Ruysbergh, E.; Adams, A.; Smagghe, G.; Kimpe, N.D.; Avlessi, F.; Sohounhloue, D.C.K.; Mangelinckx, S. Characterization of volatile compounds from three Cymbopogon species and Eucalyptus citriodora from Benin and their insecticidal activities against Tribolium castaneum. Ind. Crops Prod. 2015, 76, 306–317. [Google Scholar] [CrossRef]
- Abad, M.J.; Bedoya, L.M.; Apaza, L.; Bermejo, P. The Artemisia L. Genus: A review of bioactive essential oils. Molecules 2012, 17, 2542–2566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, R.X.; Zheng, W.F.; Tang, H.Q. Biologically active substances from the genus Artemisia. Planta Med. 1998, 64, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, J.W.; Wang, L.L.; He, C.N.; Zhao, J.; Si, L.J.; Huang, H. Artemisia scoparia: Traditional uses, active constituents and pharmacological effects. J. Ethnopharmacol. 2021, 273, 113960. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, I.S.; Cavaco, T.; Brodelius, M. Phenolic composition and antioxidant capacity of six Artemisia species. Ind. Crops Prod. 2011, 33, 382–388. [Google Scholar] [CrossRef]
- Jaume, P.; Haris, S.L.C.; Esperana, C.; Madeleine, E.; Teresa, G.; Grace, O.M.; Airy, G.; Màrius, M.; Joan, V.; Daniel, V. A phylogenetic road map to antimalarial Artemisia species. J. Ethnopharmacol. 2018, 225, 1–9. [Google Scholar] [CrossRef]
- Huang, X.; Ge, S.Y.; Liu, J.H.; Wang, Y.; Yuan, H.B. Chemical composition and bioactivity of the essential oil from Artemisia lavandulaefolia (Asteraceae) on Plutella xylostella (Lepidoptera: Plutellidae). Fla. Entomol. 2018, 101, 44–48. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.B.; Shang, L.N.; Wei, C.Y.; Ren, B.Z. Comparison of constituents and insecticidal activities of essential oil from Artemisia lavandulaefolia by steam distillation and supercritical-CO2 fluid extraction. Chem. Res. Chin. Univ. 2010, 26, 888–892. [Google Scholar] [CrossRef]
- Zhou, J.; Zou, K.X.; Zhang, W.J.; Guo, S.S.; Liu, H.; Sun, J.S.; Li, J.G.; Huang, D.Y.; Wu, Y.; Du, S.; et al. Efficacy of compounds isolated from the essential oil of Artemisia lavandulaefolia in control of the cigarette beetle, Lasioderma serricorne. Molecules 2018, 23, 343. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Huang, X.Y.; Geng, C.A.; Li, T.Z.; Chen, J.J. Cytotoxic sesquiterpenoids against hepatic stellate cell line LX2 from Artemisia lavandulaefolia. Bioorg. Chem. 2020, 103, 104107. [Google Scholar] [CrossRef]
- Cha, J.D.; Jeong, M.R.; Choi, H.J.; Jeong, S.I.; Song, Y.H. Chemical composition and antimicrobial activity of the essential oil of Artemisia lavandulaefolia. Planta Med. 2005, 71, 575–577. [Google Scholar] [CrossRef]
- Xiong, Z.W. Chemical Composition, Antioxidant and Antibacterial activity of Artemisia lavandulaefolia. Master’s Thesis, Nanchang University, Nanchang, China, 2011. [Google Scholar]
- Zhang, J.W. Composition, in vitro antioxidant and antimicrobial activity of the essential oils from Artemisa lavandulaefolia DC. J. Med. Plants Res. 2012, 6, 1284–1288. [Google Scholar] [CrossRef]
- Jordán, M.J.; Martínez, R.M.; Cases, M.A.; Sotomayor, J.A. Watering level effect on Thymus hyemalis Lange essential oil yield and composition. J. Agric. Food Chem. 2003, 51, 5420–5427. [Google Scholar] [CrossRef] [PubMed]
- Sellami, I.H.; Maamouri, E.; Chahed, T.; Wannes, W.A.; Kchouk, M.E.; Marzouk, B. Effect of growth stage on the content and composition of the essential oil and phenolic fraction of sweet marjoram (Origanum majorana L.). Ind. Crops Prod. 2009, 30, 395–402. [Google Scholar] [CrossRef]
- Kulkarni, S.A.; Nagarajan, S.K.; Ramesh, V.; Palaniyandi, V.; Mad Ha Van, T. Computational evaluation of major components from plant essential oils as potent inhibitors of SARS-CoV-2 spike protein. J. Mol. Struct. 2020, 1221, 128823. [Google Scholar] [CrossRef]
- Ren, G.L.; Zhong, Y.; Ke, G.; Liu, X.L.; Yang, M. The mechanism of compound anshen essential oil in the treatment of insomnia was examined by network pharmacology. Evid-Based Compl. Altern. 2019, 2019, 9241403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyazawa, M.; Haigou, R. Determination of cytochrome P450 enzymes involved in the metabolism of (-)-terpinen-4-ol by human liver microsomes. Xenobiotica 2011, 41, 1056–1062. [Google Scholar] [CrossRef]
- Gondim, A.N.S.; Lara, A.; Santos-Miranda, A.; Roman-Campos, D.; Lauton-Santos, S.; Menezes-Filho, J.E.R.; de Vasconcelos, C.M.L.; Conde-Garcia, E.A.; Guatimosim, S.; Cruz, J.S. (-)-Terpinen-4-ol changes intracellular Ca2+ handling and induces pacing disturbance in rat hearts. Eur. J. Pharmacol. 2017, 807, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.M.G.; Abou-Taleb, H.K.; Abdelgaleil, S.A.M. Insecticidal activities of monoterpenes and phenylpropenes against Sitophilus oryzae and their inhibitory effects on acetylcholinesterase and adenosine triphosphatases. Appl. Entomol. Zool. 2018, 53, 1–9. [Google Scholar] [CrossRef]
- Oviedo-Sarmiento, J.S.; Cortes, J.; Vila, W.; Suárez, L.; Prieto-Rodríguez, J. Fumigant toxicity and biochemical effects of selected essential oils toward the red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae). Pestic Biochem. Phys. 2021, 179, 104941. [Google Scholar] [CrossRef]
- Saad, M.; El-Deeb, D.A.; Abdelgaleil, S. Insecticidal potential and repellent and biochemical effects of phenylpropenes and monoterpenes on the red flour beetle, Tribolium castaneum Herbst. Environ. Sci. Pollut. Res. Int. 2019, 26, 6801–6810. [Google Scholar] [CrossRef]
- Yang, Y.C.; Lee, S.H.; Clark, J.M.; Ahn, Y.J. Ovicidal and adulticidal activities of Origanum majorana essential oil constituents against insecticide-susceptible and pyrethroid/malathion-resistant Pediculus humanus capitis (Anoplura: Pediculidae). J. Agric. Food Chem. 2009, 57, 2282–2287. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.R.; Perumalsamy, H.; Kwon, M.J.; Chae, S.U.; Ahn, Y.J. Toxicity of hiba oil constituents and spray formulations to American house dust mites and copra mites. Pest Manag. Sci. 2015, 71, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Sekine, N.; Shibutani, S. Chemical structures of p-menthane monoterpenes with special reference to their effect on seed germination and termite mortality. J. Wood Sci. 2013, 59, 229–237. [Google Scholar] [CrossRef]
- Piri, A.; Sahebzadeh, N.; Zibaee, A.; Sendi, J.J.; Shahriari, M. Toxicity and physiological effects of ajwain (Carum copticum, Apiaceae) essential oil and its major constituents against Tuta absoluta (Meyrick) (Lepidoptera: Gelechiidae). Chemosphere 2020, 256, 127103. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Song, L.; Sun, J.; Cao, H.Q.; Wang, L.K.; Lin, H.F.; Tang, F. Repellent action and contact toxicity mechanisms of the essential oil extracted from Chinese chive against Plutella xylostella larvae. Arch. Insect Biochem. 2019, 100, 21509. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.H.; Bai, Q.; Zheng, X.S.; Lu, Z.X. Expression and enzyme activity of catalase in Chilo suppressalis (Lepidoptera: Crambidae) is responsive to environmental stresses. J. Econ. Entomol. 2017, 110, 1803–1812. [Google Scholar] [CrossRef]
- Hematpoor, A.; Liew, S.Y.; Azirun, M.S.; Awang, K. Insecticidal activity and the mechanism of action of three phenylpropanoids isolated from the roots of Piper sarmentosum Roxb. Sci. Rep. 2017, 7, 12576. [Google Scholar] [CrossRef] [Green Version]
- Czerniewicz, P.; Chrzanowski, G.; Sprawka, I.; Sytykiewicz, H. Aphicidal activity of selected Asteraceae essential oils and their effect on enzyme activities of the green peach aphid, Myzus persicae (Sulzer). Pestic Biochem. Phys. 2018, 145, 84–92. [Google Scholar] [CrossRef]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Am. Mosq. Control Assoc. 1987, 3, 302–303. [Google Scholar] [CrossRef]
- Finney, D.J.; Tattersfield, F. Probit Analysis; University Press: Cambridge, UK, 1952. [Google Scholar]
- Tian, J.; Ban, X.Q.; Zeng, H.; He, J.S.; Huang, B.; Wang, Y.W. Chemical composition and antifungal activity of essential oil from Chemical L. var latisecta Celak. Int. J. Food Microbiol. 2011, 145, 464–470. [Google Scholar] [CrossRef]
- Zapata, N.; Smagghe, G. Repellency and toxicity of essential oils from the leaves and bark of Laurelia sempercirens and Drimys winteri against Tribolium castaneum. Ind. Crops Prod. 2010, 32, 405–410. [Google Scholar] [CrossRef]
- Chaubey, M.K. Insecticidal properties of Zingiber officinale and Piper cubebacu essential oils against Tribolium castaneum Herbst (Coleoptera: Tenebrionidae). J. Biol. Act. Prod. Nat. 2011, 1, 306–313. [Google Scholar] [CrossRef]
- Stefanazzi, N.; Stadler, T.; Ferrero, A. Composition and toxic, repellent and feeding deterrent activity of essential oils against the stored-grain pests Tribolium castaneum (Coleoptera: Tenebrionidae) and Sitophilus oryzae (Coleoptera: Curculionidae). Pest Manag. Sci. 2011, 67, 639–646. [Google Scholar] [CrossRef]
- Chaubey, M.K. Essential oils as green pesticides of stored grain insects. Eur. J. Biol. Res. 2019, 9, 202–244. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Prajapati, V.; Aggarwal, K.K.; Kumar, S. Toxicity, feeding deterrence, and effect of activity of 1,8-cineole from Artemisia annua on progeny production of Tribolium castanaeum (Coleoptera: Tenebrionidae). J. Econ. Entomol. 2001, 94, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Choi, W.S.; Lee, S.E.; Park, B.S. Fumigant toxicity of essential oils and their constituent compounds towards the rice weevil, Sitophilus oryzae (L.). Crops Prot. 2001, 20, 317–320. [Google Scholar] [CrossRef]
- Liu, J.; Hua, J.; Qu, B.; Guo, X.; Wang, Y.; Shao, M.; Luo, S. Insecticidal terpenes from the essential oils of Artemisia nakaii and their inhibitory effects on acetylcholinesterase. Front. Plant Sci. 2021, 12, 720816. [Google Scholar] [CrossRef]
- Li, B. Extraction, Chemical Constituents and Insecticidal Activity of Essential Oil from Chenopodium ambrosioides L. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2007. [Google Scholar]
- Liu, P.; Liu, X.C.; Dong, H.W.; Liu, Z.L.; Du, S.S.; Deng, Z. Chemical composition and insecticidal activity of the essential oil of Illicium pachyphyllum fruits against two grain storage insects. Molecules 2012, 17, 14870–14881. [Google Scholar] [CrossRef]
- Muturi, E.J.; Doll, K.; Berhow, M.; Flor-Weiler, L.B.; Rooney, A.P. Honeysuckle essential oil as a potential source of ecofriendly larvicides for mosquito control. Pest Manag. Sci. 2019, 75, 2043–2048. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.A.H.; Ling, S.; Tian, F.; Xie, F.; Zeng, X. Toxicity and enzyme inhibition activities of the essential oil and dominant constituents derived from Artemisia absinthium L. against adult Asian citrus psyllid Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Ind. Crops Prod. 2018, 121, 468–475. [Google Scholar] [CrossRef]
- Singh, R.; Koul, O.; Rup, P.J.; Jindal, J. Toxicity of some essential oil constituents and their binary mixtures against Chilo partellus (Lepidoptera: Pyralidae). Int. J. Trop. Insect Sci. 2009, 29, 93–101. [Google Scholar] [CrossRef]
- Kiran, S.; Prakash, B. Toxicity and biochemical efficacy of chemically characterized Rosmarinus officinalis essential oil against Sitophilus oryzae and Oryzaephilus surinamensis. Ind. Crops Prod. 2015, 74, 817–823. [Google Scholar] [CrossRef]
- Grant, D.F.; Matsumura, F. Glutathione S-transferase 1 and 2 in susceptible and insecticide resistant Aedes aegypti. Pestic Biochem. Phys. 1989, 33, 132–143. [Google Scholar] [CrossRef]
- Liao, M.; Xiao, J.J.; Zhou, L.J.; Yao, X.; Tang, F.; Hua, R.M.; Wu, X.W.; Cao, H.Q. Chemical composition, insecticidal and biochemical effects of Melaleuca alternifolia essential oil on the Helicoverpa armigera. J. Appl. Entomol. 2017, 141, 721–728. [Google Scholar] [CrossRef] [Green Version]
- Rajkumar, V.; Gunasekaran, C.; Christy, I.K.; Dharmaraj, J. Toxicity, antifeedant and biochemical efficacy of Mentha piperita L. essential oil and their major constituents against stored grain pest. Pestic Biochem. Phys. 2019, 156, 138–144. [Google Scholar] [CrossRef]
- Kiran, S.; Prakash, B. Assessment of toxicity, antifeedant activity, and biochemical responses in stored-grain insects exposed to lethal and sublethal doses of Gaultheria procumbens L. essential oil. J. Agric. Food Chem. 2015, 63, 10518–10524. [Google Scholar] [CrossRef]
- Magierowicz, K.; Górska-Drabik, E.; Sempruch, C. The insecticidal activity of Satureja hortensis essential oil and its active ingredient-carvacrol against Acrobasis advenella (Zinck.) (Lepidoptera, Pyralidae). Pestic Biochem. Phys. 2019, 153, 122–128. [Google Scholar] [CrossRef]
- Miyazawa, M.; Yamafuji, C. Inhibition of acetylcholinesterase activity by bicyclic monoterpenoids. J. Agric. Food Chem. 2005, 53, 1765–1768. [Google Scholar] [CrossRef]
- Rajashekar, Y.; Raghavendra, A.; Bakthavatsalam, N. Acetylcholinesterase inhibition by biofumigant (Coumaran) from leaves of Lantana camara in stored grain and household insect pests. Biomed. Res. Int. 2014, 2014, 187019. [Google Scholar] [CrossRef] [Green Version]
- Qin, W.Q.; Huang, S.C.; Li, C.X.; Chen, S.T.; Peng, Z.Q. Biological activity of the essential oil from the leaves of Piper sarmentosum Roxb. (Piperaceae) and its chemical constituents on Brontispa longissima (Gestro) (Coleoptera: Hispidae). Pestic Biochem. Phys. 2010, 96, 132–139. [Google Scholar] [CrossRef]
- Oyedeji, A.O.; Okunowo, W.O.; Osuntoki, A.A.; Olabode, T.B.; Ayo-Folorunso, F. Insecticidal and biochemical activity of essential oil from Citrus sinensis peel and constituents on Callosobrunchus maculatus and Sitophilus zeamais. Pestic Biochem. Phys. 2020, 168, 104643. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
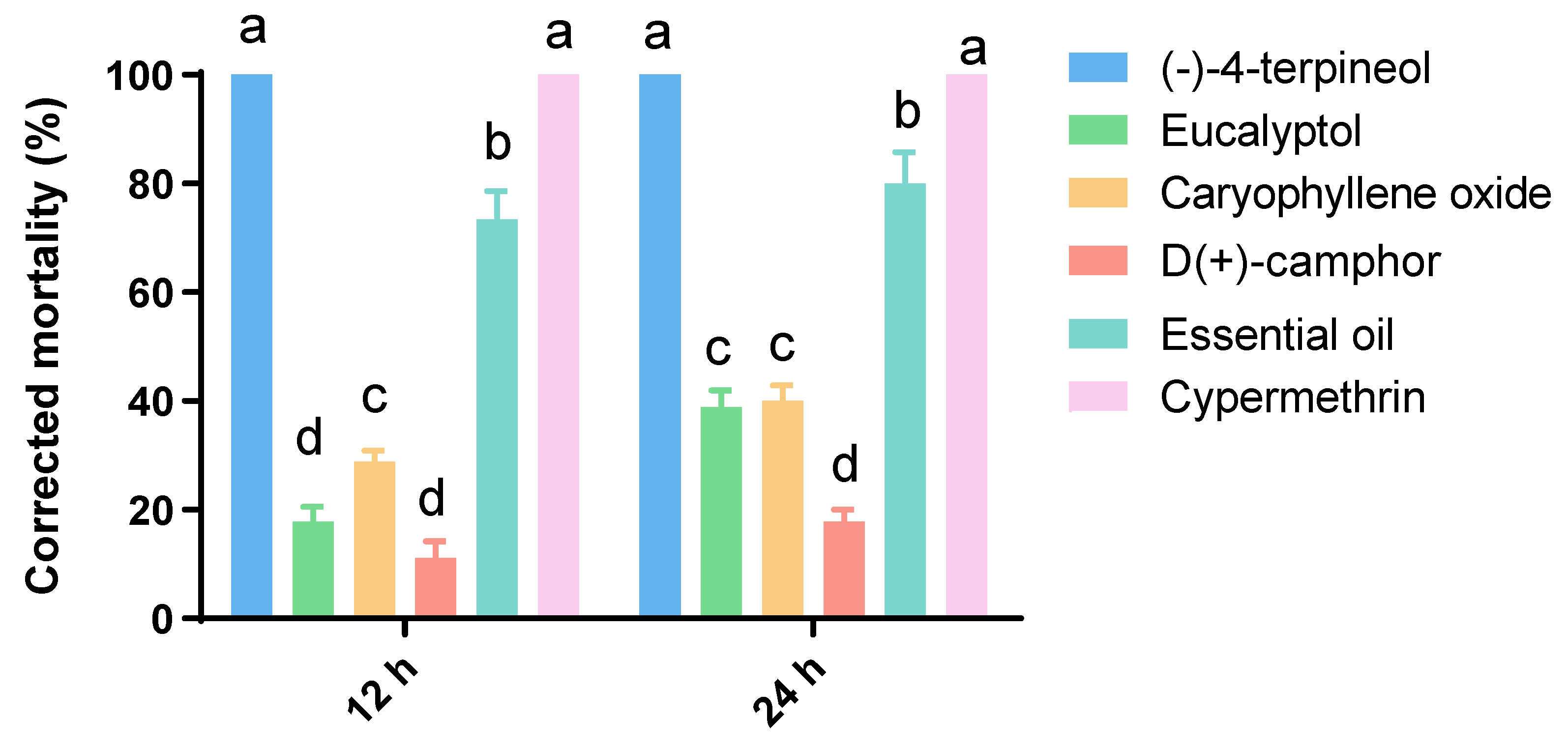{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time/min | RI 2 | Name of Constituent | Relative Content/% | Identifaicaion Methods | CAS 4 |
|---|---|---|---|---|---|
| 7.743 | 1022 | Eucalyptol | 21.57 | MS 1, RI | 470-82-6 |
| 8.7181 | 1072 | Artemisia alcohol | 3.90 | MS, RI | 27644-04-8 |
| 9.1215 | 1083 | Nonanal | 0.42 | MS, RI | 124-19-6 |
| 9.1685 | 1119 | Filifolone | 3.20 | MS, RI | 4613-37-0 |
| 9.2205 | 1089 | Thujone | 1.59 | MS, RI | 546-80-5 |
| 9.4407 | 1149 | Chrysanthenol | 1.95 | MS, RI | 18383-58-9 |
| 9.6387 | 1103 | 2-pinen-7-one | 0.80 | MS, RI | 473-06-3 |
| 9.8565 | 1163 | Isocyclocitral | 1.05 | MS, RI | 1335-66-6 |
| 10.1188 | 1121 | D(+)-camphor | 17.33 | MS, RI | 464-49-3 |
| 10.4331 | 1147 | cis-chrysanthenol | 1.77 | MS, RI | 55722-60-6 |
| 10.8217 | 1160 | (-)-4-terpineol | 9.96 | MS, RI, Co 3 | 20126-76-5 |
| 10.943 | 1171 | (-)-myrtenal | 0.27 | MS, RI | 564-94-3 |
| 12.2175 | 1230 | 3-Isopropylbenzaldehyde | 0.25 | MS, RI | 34246-57-6 |
| 12.9526 | 1190 | (+)-isopiperitenone | 0.29 | MS, RI | 16750-82-6 |
| 13.0268 | 1252 | perillyl aldehyde | 0.47 | MS, RI | 2111-75-3 |
| 13.2718 | 1273 | (-)-Bornyl acetate | 0.43 | MS, RI | 5655-61-8 |
| 13.9351 | 1279 | (E,Z)-2,4-decadienal | 0.26 | MS, RI | 25152-83-4 |
| 14.7519 | 1333 | Terpinyl acetate | 0.45 | MS, RI | 80-26-2 |
| 15.8285 | 1120 | 3-caren-5-one | 1.82 | MS, RI | 81800-50-2 |
| 16.0314 | 1372 | Methyl eugenol | 0.42 | MS, RI | 93-15-2 |
| 17.6526 | 1457 | 4-(2,2,6-trimethyl-bicyclo [4.1.0]hept-1-yl)-butan-2-one | 0.35 | MS, RI | 77143-20-5 |
| 17.9199 | 1586 | Zizanol | 0.35 | MS, RI | 28102-79-6 |
| 18.2764 | 1325 | Agropyrene | 1.69 | MS, RI | 520-74-1 |
| 19.3952 | 1521 | Cabreuva oxide | 0.25 | MS, RI | 107602-52-8 |
| 19.6278 | 1577 | β-Caryophyllene | 1.18 | MS, RI | 87-44-5 |
| 19.7813 | 1578 | β-caryophyllene alcohol | 0.83 | MS, RI | 58404-89-0 |
| 19.9868 | 1563 | Palustrol | 0.29 | MS, RI | 5986-49-2 |
| 20.1625 | 1419 | Caryophyllene | 0.28 | MS, RI | 69842-07-5 |
| 20.3679 | 1574 | Caryophyllene oxide | 10.96 | MS, RI | 1139-30-6 |
| 20.5858 | 1584 | Salvial-4(14)-en-1-one | 0.32 | MS, RI | 73809-82-2 |
| 20.8358 | 1534 | Copaborneol | 0.57 | MS, RI | 21966-93-8 |
| 21.1576 | 1613 | Junenol | 1.37 | MS, RI | 472-07-1 |
| 21.2269 | 1688 | 8-cedren-13-ol | 0.67 | MS, RI | 18319-35-2 |
| 21.3135 | 1540 | Selina-3,7(11)-diene | 0.93 | MS, RI | 6813-21-4 |
| 21.3803 | 1631 | Longifolenaldehyde | 0.40 | MS, RI | 19890-84-7 |
| 21.4521 | 1376 | Copaene | 0.40 | MS, RI | 3856-25-5 |
| 21.5313 | 1604 | Widdrenal | 0.26 | MS, RI | 470-41-7 |
| 21.6427 | 1873 | (2-Butyrylphenoxy) acetic acid | 0.27 | MS, RI | 65627-57-8 |
| 21.7863 | 1621 | Desmethoxyencecalin | 1.31 | MS, RI | 19013-07-1 |
| 22.1601 | 1324 | 10,10-Dimethyl-4-acetyl-tricyclo [5.2.1.0(1,5)]decane | 0.45 | MS, RI | 176171-97-4 |
| 22.4843 | 1668 | Bisabolol | 0.33 | MS, RI | 515-69-5 |
| 24.1602 | 1710 | 4,5-Dichloro-2-octyl-isothiazolone | 1.04 | MS, RI | 64373-81-5 |
| 24.6627 | 1810 | Methyl isocostate | 1.92 | MS, RI | 132342-55-3 |
| 25.1578 | 1842 | Fitone | 0.51 | MS, RI | 502-69-2 |
| 25.7718 | 1724 | Euparine | 0.36 | MS, RI | 532-48-9 |
| 26.4055 | 2513 | Cinerin II | 0.60 | MS, RI | 121-20-0 |
| 26.5491 | 1236 | β-homocyclocitral | 0.43 | MS, RI | 472-66-2 |
| Dose | KT50 4/95% FL 5 | Regression Equation | χ2 | p-Value |
|---|---|---|---|---|
| LC30 1 | 9.50 (9.16–9.87) | y = 7.586x − 7.418 | 3.664 | 0.300 |
| LC50 2 | 6.07 (9.16–9.87) | y = 11.307x − 8.852 | 6.193 | 0.103 |
| LC80 3 | 5.09 (4.65–5.50) | y = 10.365x − 7.323 | 7.847 | 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Du, L.; Liu, T.; Ma, R.; Liu, X.; Yuan, H.; Liu, S. Insecticidal Activity of a Component, (-)-4-Terpineol, Isolated from the Essential Oil of Artemisia lavandulaefolia DC. against Plutella xylostella (L.). Insects 2022, 13, 1126. https://doi.org/10.3390/insects13121126
Huang X, Du L, Liu T, Ma R, Liu X, Yuan H, Liu S. Insecticidal Activity of a Component, (-)-4-Terpineol, Isolated from the Essential Oil of Artemisia lavandulaefolia DC. against Plutella xylostella (L.). Insects. 2022; 13(12):1126. https://doi.org/10.3390/insects13121126
Chicago/Turabian StyleHuang, Xing, Linjie Du, Tiantian Liu, Rui Ma, Xing Liu, Haibin Yuan, and Shuai Liu. 2022. "Insecticidal Activity of a Component, (-)-4-Terpineol, Isolated from the Essential Oil of Artemisia lavandulaefolia DC. against Plutella xylostella (L.)" Insects 13, no. 12: 1126. https://doi.org/10.3390/insects13121126