
Molecular Phylogeny and Taxonomy of the Butterfly Subtribe Scolitantidina with Special Focus on the Genera Pseudophilotes, Glaucopsyche and Iolana (Lepidoptera, Lycaenidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Studies
2.3. Genus and Subgenus Concepts
2.4. Methodology of Molecular Taxonomy: Genomics, Phylogenomics, DNA-barcoding, and Mixed (Phylogenomics + Barcoding) Approaches
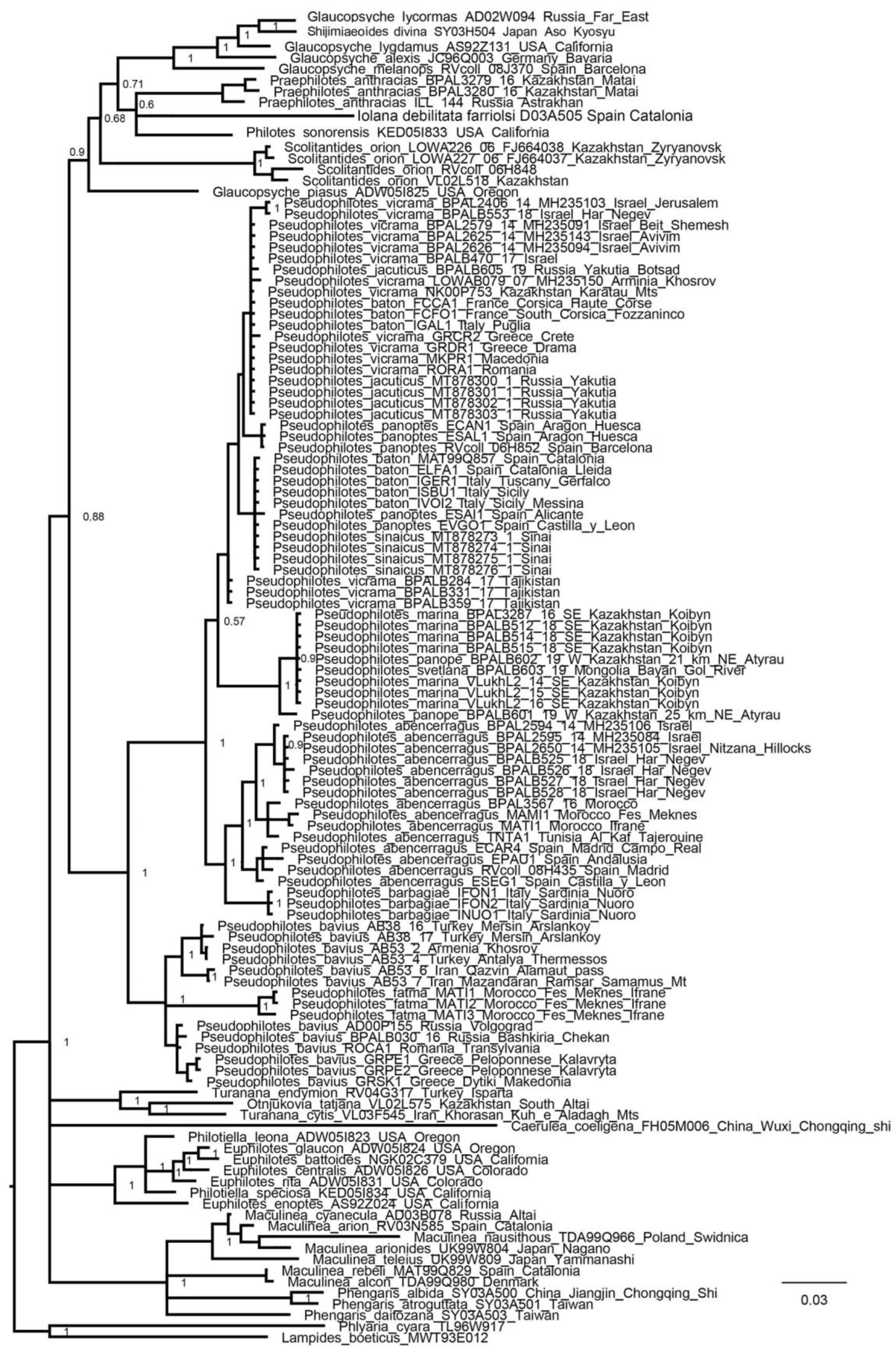
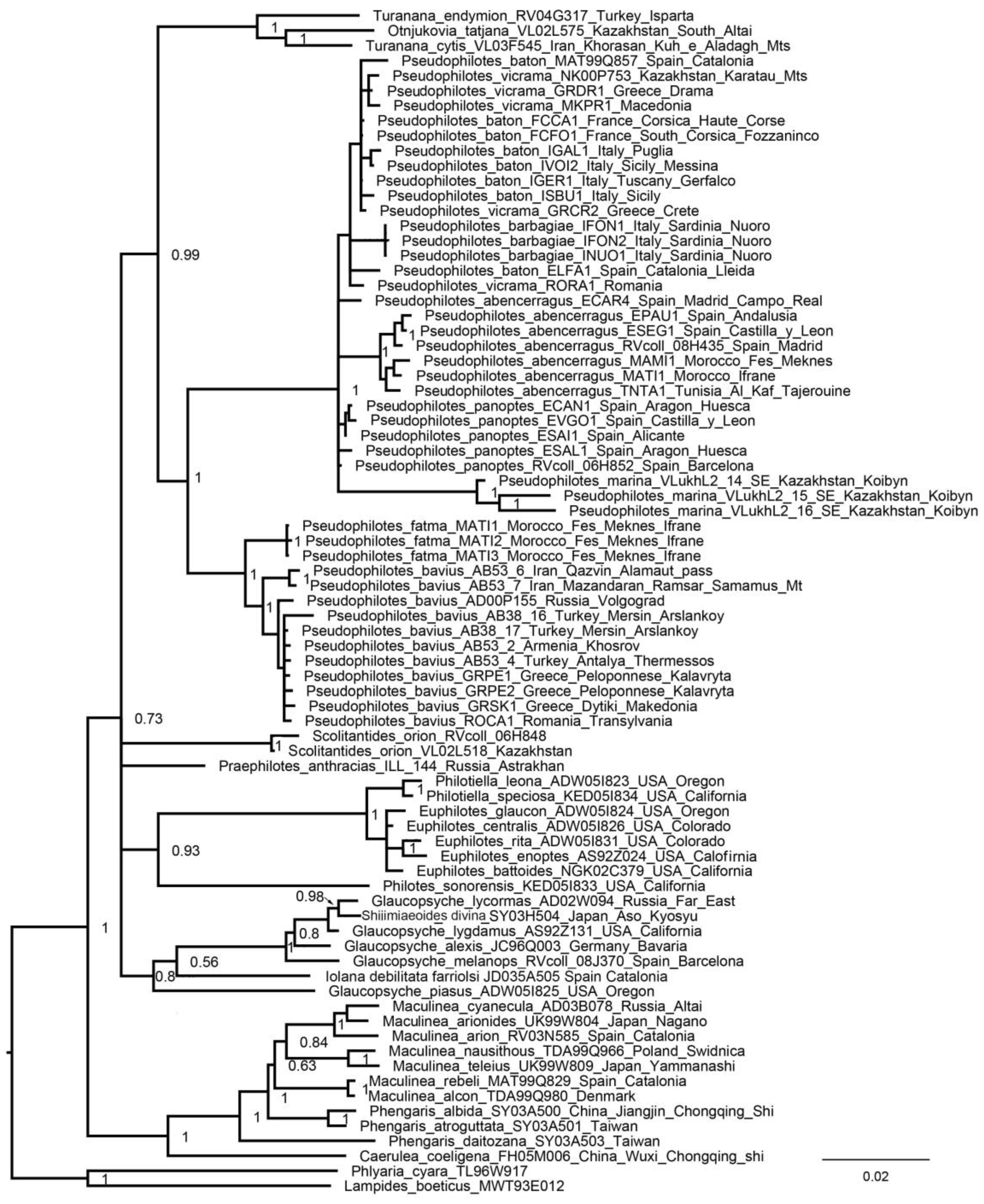
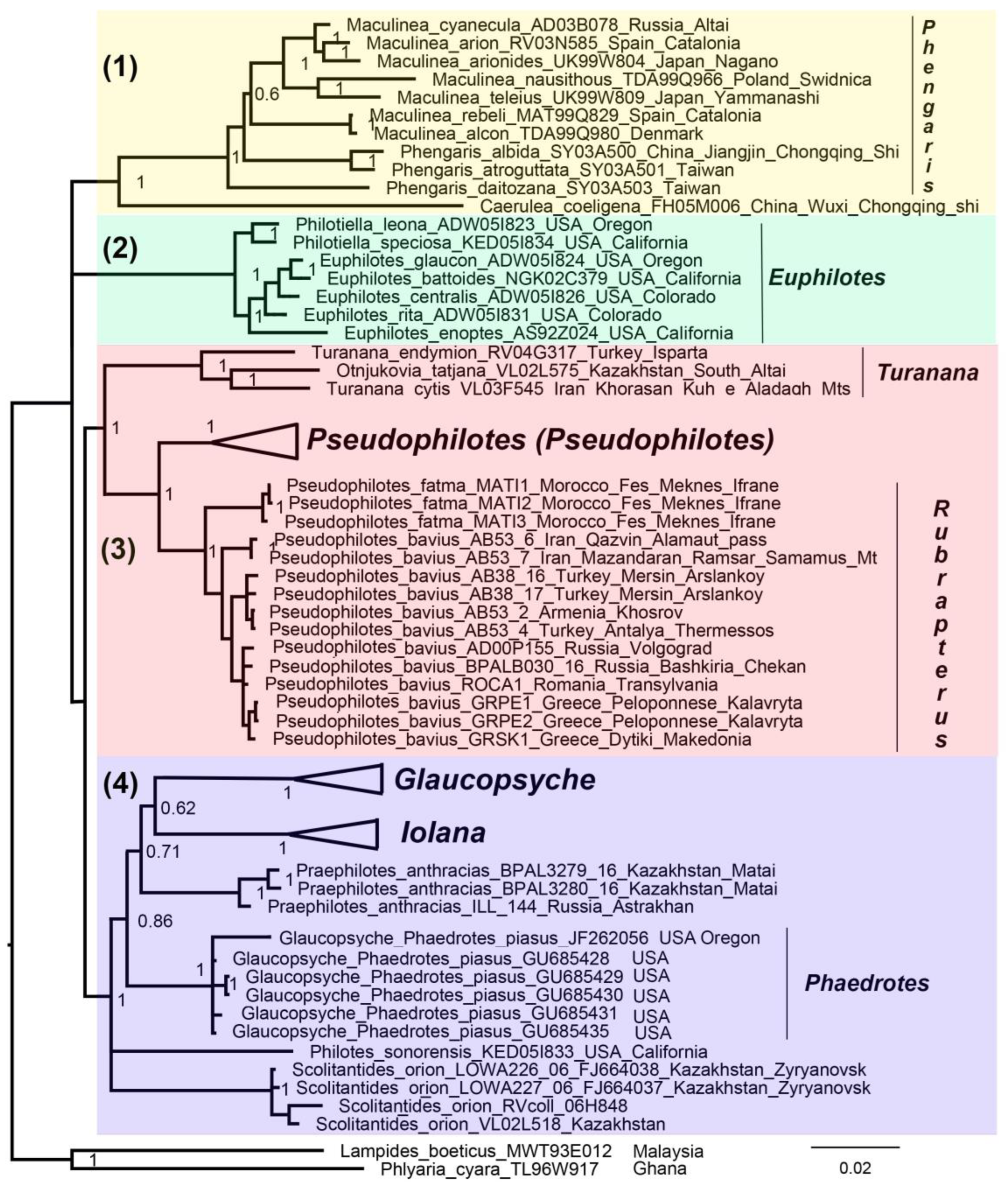
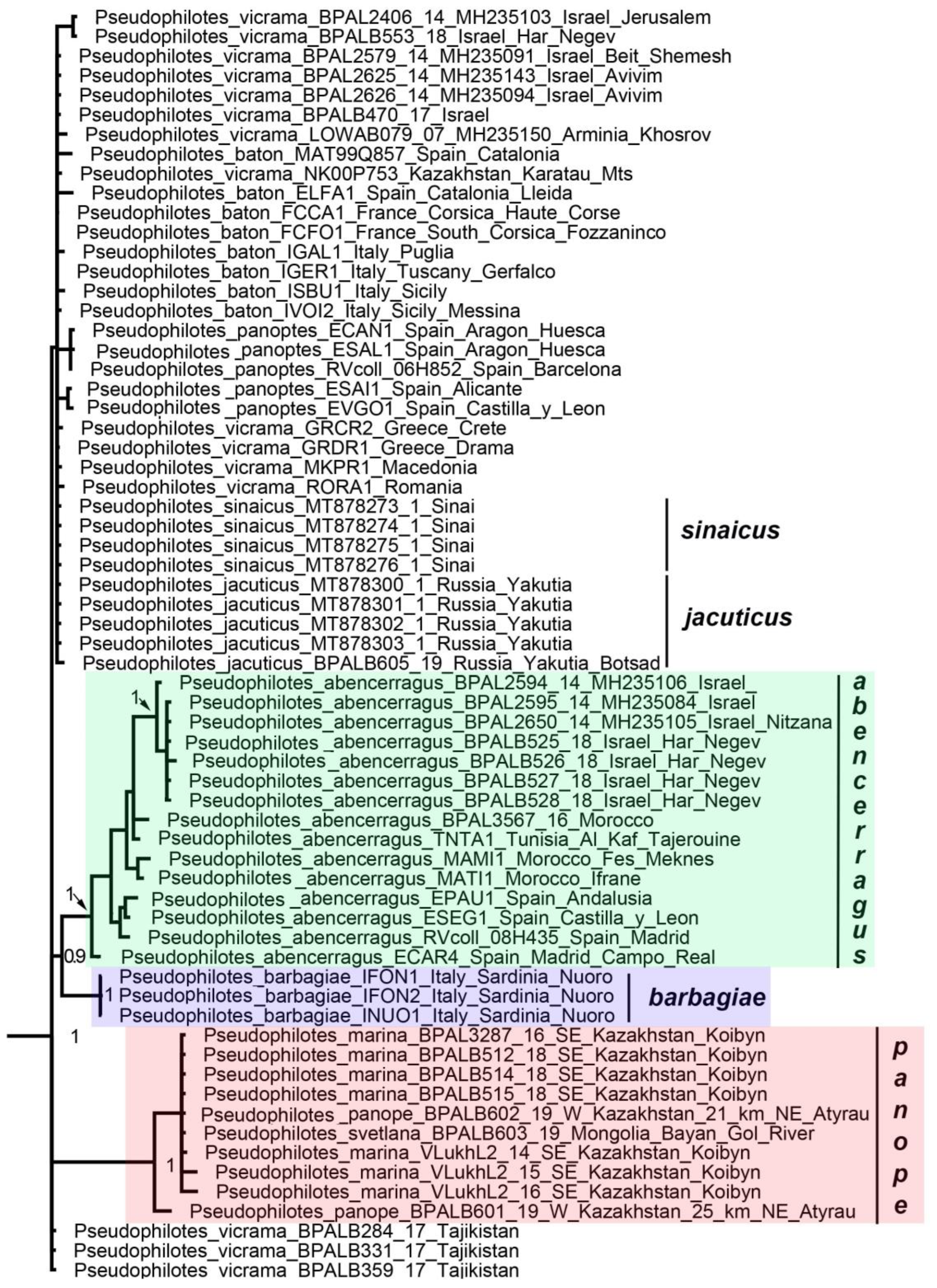
3. Results
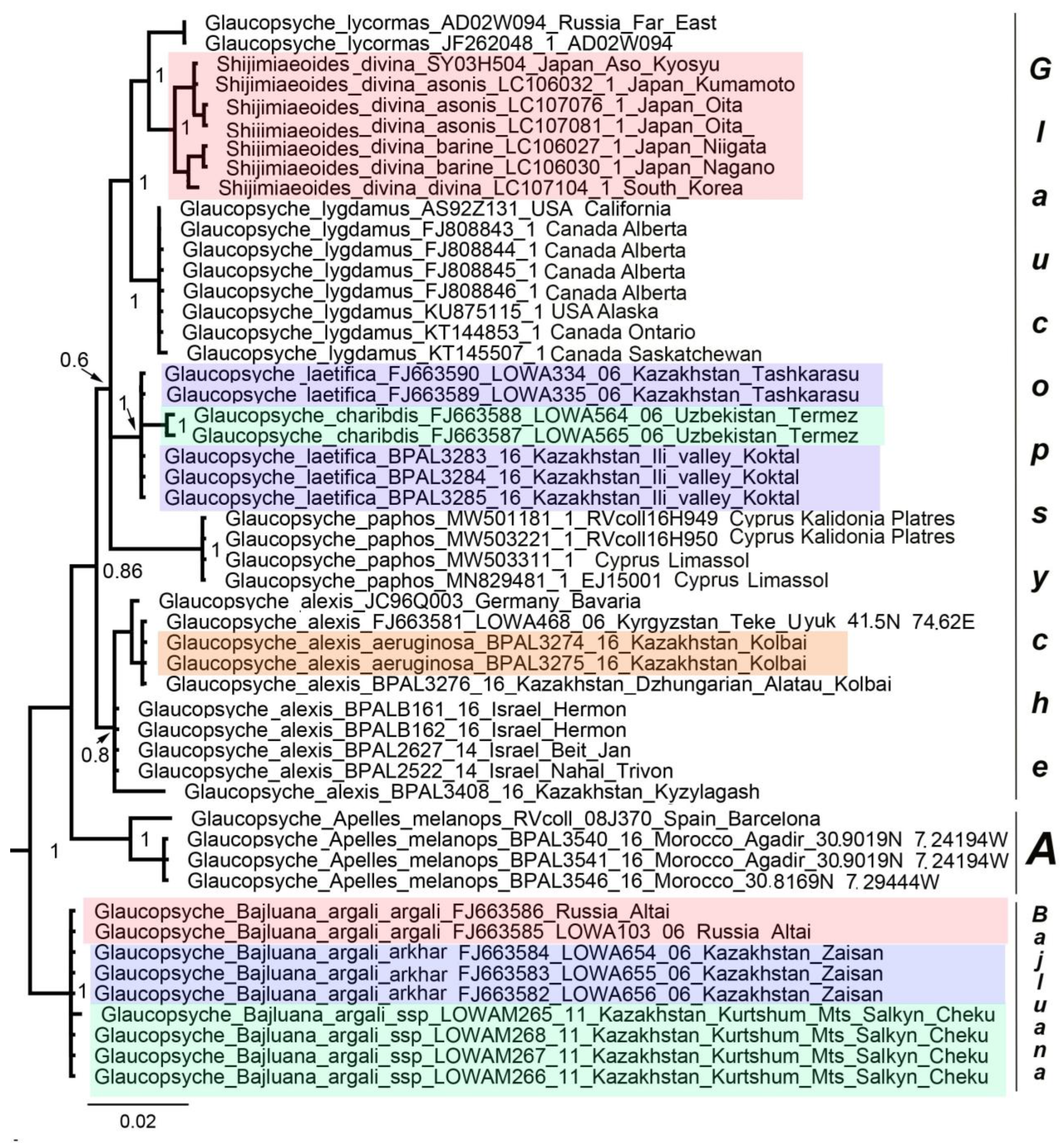
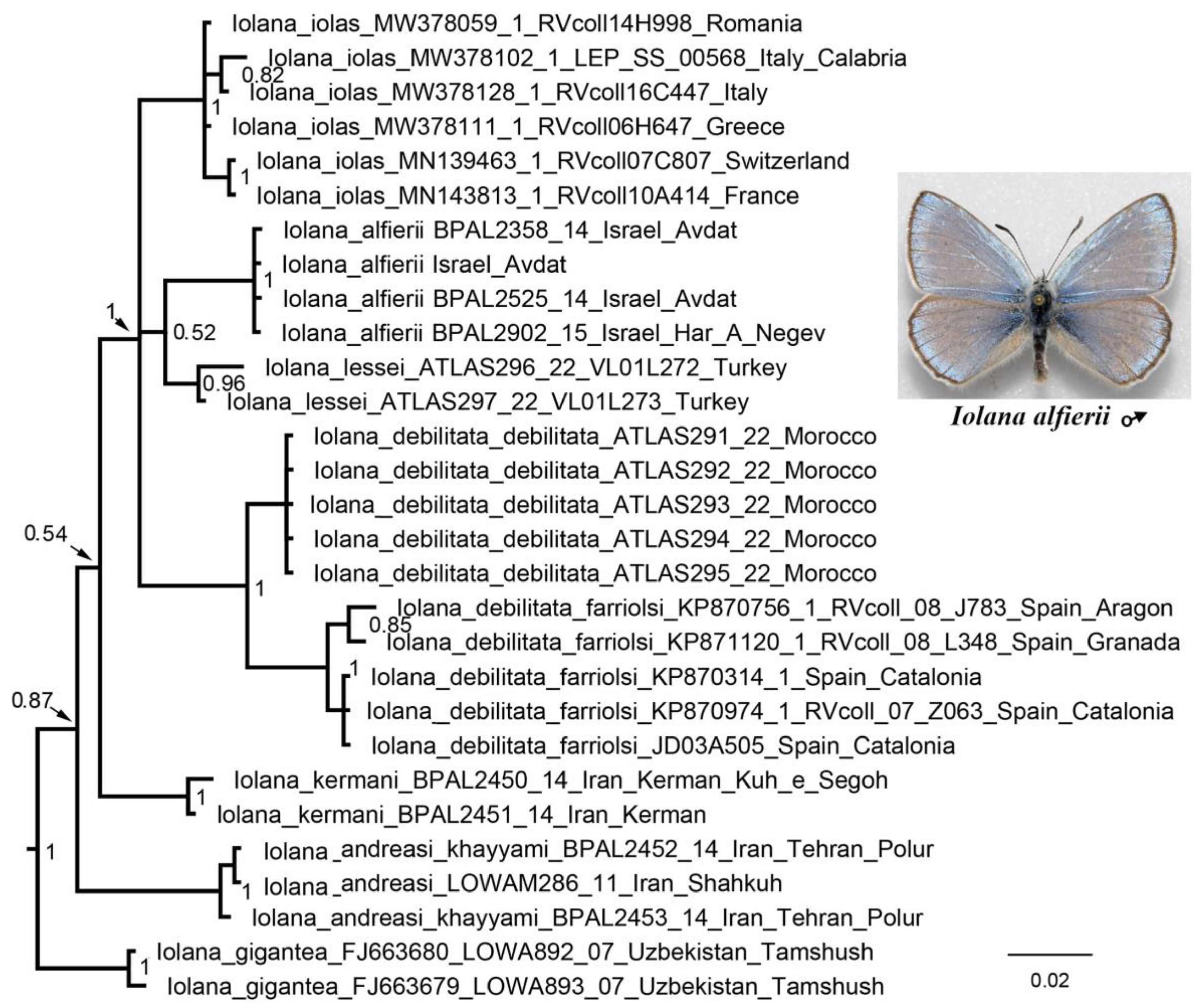
3.1. Mitochondrial Tree
3.2. Nuclear Tree
3.3. Concatenated Tree
4. Discussion
5. Taxonomic Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polyommatinae. Available online: https://ftp.funet.fi/index/Tree_of_life/insecta/lepidoptera/ditrysia/papilionoidea/lycaenidae/polyommatinae/ (accessed on 28 November 2022).
- Eliot, J.N. The higher classification of the Lycaenidae (Lepidoptera): A tentative arrangement. Bull. Nat. Hist. Mus. Entomol. Ser. 1973, 28, 371–505. [Google Scholar] [CrossRef]
- Mattoni, R.H.T. The Scolitantidini. I: Two new genera and a generic rearrangement (Lycaenidae). J. Res. Lepid. 1977, 16, 223–242. [Google Scholar] [CrossRef]
- Hesselbarth, G.; Van Oorschot, H.; Wagener, S. Die Tagfalter der Türkei; Selbstverlag Sigbert Wagener: Bocholt, Germany, 1995; Volume 1, 754p. [Google Scholar]
- Als, T.; Vila, R.; Kandul, N.; Nash, D.R.; Yen, S.H.; Hsu, Y.; Mignault, A.; Boomsma, J.J.; Pierce, N.E. The evolution of alternative parasitic life histories in large blue butterflies. Nature 2004, 432, 386–390. [Google Scholar] [CrossRef] [PubMed]
- Ugelvig, L.V.; Vila, R.; Pierce, N.E.; Nash, D.R. A phylogenetic revision of the Glaucopsyche section (Lepidoptera: Lycaenidae), with special focus on the Phengaris–Maculinea clade. Mol. Phylogenet. Evol. 2011, 61, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Fric, Z.; Wahlberg, N.; Pech, P.; Zrzavy, J.A.N. Phylogeny and classification of the Phengaris–Maculinea clade (Lepidoptera: Lycaenidae): Total evidence and phylogenetic species concepts. Syst. Entomol. 2007, 32, 558–567. [Google Scholar] [CrossRef]
- Zhang, J.; Cong, Q.; Shen, J.; Opler, P.A.; Grishin, N.V. Changes to North American butterfly names. Taxon. Rep. Int. Lepid. Surv. 2019, 8, 1–11. [Google Scholar]
- Zhang, J.; Cong, Q.; Shen, J.; Song, L.; Gott, R.J.; Boyer, P.; Guppy, S.; Kohler, S.; Lamas, G.; Opler, P.A.; et al. Taxonomic discoveries enabled by genomic analysis of butterflies. Taxon. Rep. Int. Lepid. Surv. 2022, 10, 1–59. [Google Scholar]
- Korshunov, Y.P. New Taxones [sic] of Lepidopterous [sic] (Lepidoptera, Rhopalocera) in Eurasia. In Nasekomye, Klestchi i Gelminty, Novye i Maloizvestnye Vidy Fauny Sibiri; Cherepanov, A.I., Ed.; Nauka, Sibirian Branch: Novosibirsk, Russia, 1987; pp. 9–13. [Google Scholar]
- Kolev, Z. Rubrapterus bavius (Eversmann, 1832), a butterfly genus and species new to Bulgaria (Insecta, Lepidoptera, Lycaenidae). ZooNotes 2017, 114, 1–4. [Google Scholar]
- Korshunov, Y.P. Butterflies of Ural, Siberia and Far East (Keys and Details); Vitel: Novosibirsk, Russia, 2000; 218p. (In Russian) [Google Scholar]
- Lukhtanov, V.A.; Lukhtanov, A.G. Die Tagfalter Nordwestasiens (Lepidoptera, Diurna); U. Eitschberger: Marktleuthen, Germany, 1994; 440p, ISBN 3-923807-02-3. [Google Scholar]
- Zhdanko, A.B. Notes on the systematic position of Inderskia Korshunov, 2000, with the description of Paleophilotes (Inderskia) marina sp. n. from East Kazakhstan (Lepidoptera, Lycaenidae). In Helios. Collection of Lepidopterological Articles, Vol. V; Churkin, S., Ed.; Chronos-Press: Moscow, Russia, 2004; pp. 68–74. [Google Scholar]
- Morgun, D.V. Notes on the Paleophilotes (Inderskia) panope (Eversmann, 1851) (Lepidoptera: Lycaenidae), a rare and little known lycaenid taxon for Europe. Zootaxa 2013, 3619, 595–600. [Google Scholar] [CrossRef] [Green Version]
- Todisco, V.; Grill, A.; Fiedler, K.; Gottsberger, B.; Dincă, V.; Vodă, R.; Lukhtanov, V.; Letsch, H. Molecular phylogeny of the Palaearctic butterfly genus Pseudophilotes (Lepidoptera: Lycaenidae) with focus on the Sardinian endemic P. barbagiae. BMC Zool. 2018, 3, 4. [Google Scholar] [CrossRef]
- Leigheb, G.; Cameron-Curry, V. Observations on the biology and distribution of Pseudophilotes barbagiae (Lycaenidae: Polyommatini). Nota Lepid. 1998, 21 (Suppl. 1), 66–73. [Google Scholar]
- James, M.; Gilbert, F.; Zalat, S. Thyme and isolation for the Sinai baton blue butterfly (Pseudophilotes sinaicus). Oecologia 2003, 134, 445–453. [Google Scholar] [CrossRef] [PubMed]
- James, M. The natural history of the Sinai Baton Blue: The smallest butterfly in the world. Egypt. J. Biol. 2006, 8, 67–83. [Google Scholar] [CrossRef]
- Bartoňová, A.S.; Konvička, M.; Marešová, J.; Kolev, Z.; Wahlberg, N.; Fric, Z.F. Recently lost connectivity in the Western Palaearctic steppes: The case of a scarce specialist butterfy. Conserv. Genet. 2020, 21, 561–575. [Google Scholar] [CrossRef]
- Bartoňová, A.S.; Konvička, M.; Marešová, J.; WIemers, M.; Ignatev, N.; Wahlberg, N.; Schmitt, T.; Fric, Z.F. Wolbachia afects mitochondrial population structure in two systems of closely related Palaearctic blue butterfies. Sci. Rep. 2021, 11, 3019. [Google Scholar] [CrossRef]
- Fiedler, K. Systematic, evolutionary, and ecological implications of myrmecophily within the Lycaenidae (Insecta: Lepidoptera: Papilionoidea). Bonn. Zool. Monogr. 1991, 31, 1–211. [Google Scholar]
- Német, E.; Czekes, Z.; Markó, B.; Rákosy, L. Host plant preference in the protected myrmecophilous Transylvanian Blue (Pseudophilotes bavius hungarica) butterfly (Lepidoptera: Lycaenidae) and its relationship with potential ant partners. J. Insect. Conserv. 2016, 20, 765–772. [Google Scholar] [CrossRef]
- Dincă, V.; Cuvelier, S.; Mølgaard, M.S. Distribution and conservation status of (Lepidoptera: Lycaenidae) in Dobrogea (south-eastern Romania). Phegea 2011, 39, 59–67. [Google Scholar]
- van Swaay, C.; Cuttelod, A.; Collins, S.; Maes, D.; López Munguira, M.; Šašić, M.; Settele, J.; Verovnik, R.; Verstrael, T.; Warren, M.; et al. European Red List of Butterfies; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- van Swaay, C.; Wynhoff, I.; Verovnik, R.; Wiemers, M.; López Munguira, M.; Maes, D.; Sasic, M.; Verstrael, T.; Warren, M.; Settele, J. Pseudophilotes Baton. The IUCN Red List of Threatened Species; Red List: London, UK, 2010; e.T173277A6983969. [Google Scholar] [CrossRef]
- van Swaay, C.; Wynhoff, I.; Verovnik, R.; Wiemers, M.; López Munguira, M.; Maes, D.; Sasic, M.; Verstrael, T.; Warren, M.; Settele, J. Pseudophilotes Panoptes. The IUCN Red List of Threatened Species; Red List: London, UK, 2010; e.T173247A6979667. [Google Scholar] [CrossRef]
- van Swaay, C.; Wynhoff, I.; Wiemers, M.; Katbeh-Bader, A.; Power, A.; Benyamini, D.; Tzirkalli, E.; Balletto, E.; Monteiro, E.; Karaçetin, E.; et al. Pseudophilotes Barbagiae. The IUCN Red List of Threatened Species; Red List: London, UK, 2015; e.T173207A64553436. [Google Scholar] [CrossRef]
- van Swaay, C.; Wynhoff, I.; Wiemers, M.; Katbeh-Bader, A.; Power, A.; Benyamini, D.; Tzirkalli, E.; Balletto, E.; Monteiro, E.; Karaçetin, E.; et al. Pseudophilotes Fatma. The IUCN Red List of Threatened Species; Red List: London, UK, 2015; e.T62148778A62151921. [Google Scholar] [CrossRef]
- van Swaay, C.; Wynhoff, I.; Wiemers, M.; Katbeh-Bader, A.; Power, A.; Benyamini, D.; Tzirkalli, E.; Balletto, E.; Monteiro, E.; Karaçetin, E.; et al. Pseudophilotes Jordanicus. The IUCN Red List of Threatened Species; Red List: London, UK, 2015; e.T62148766A62151947. [Google Scholar] [CrossRef]
- Shchurov, V. Pseudophilotes vicrama schiffermulleri Hemming, 1929. In Red Data Book of Krasnodar Territory (Animals), 2nd ed.; Zamotajlov, A.S., Shchurov, V.I., Eds.; Tsentr Razvitiya pechati: Krasnodar, Russia, 2007; pp. 267–268. ISBN 5-98630-015-3. [Google Scholar]
- Shchurov, V.I. Pseudophilotes bavius (Eversmann, 1832). In Red Data Book of Krasnodar Territory (Animals), 2nd ed.; Zamotajlov, A.S., Shchurov, V.I., Eds.; Tsentr Razvitiya pechati: Krasnodar, Russia, 2007; pp. 265–267. ISBN 5-98630-015-3. [Google Scholar]
- Thompson, K.; Gilbert, F. Pseudophilotes Sinaicus. The IUCN Red List of Threatened Species; Red List: London, UK, 2012. [Google Scholar] [CrossRef]
- De Prins, W.; Van der Poorten, D. Overzicht van het genus Pseudophilotes in Europa en Noord Afrika met beschrijving van een soort uit Sardinie, nieuw voor de wetenschap. Phegea 1982, 10 (Suppl. S2), 61–76. [Google Scholar]
- Yakovlev, R.V. Pseudophilotes (Inderskia) Svetlana sp. n., A New Blue from Mongolian Altai (Lepidoptera, Lycaenidae). Helios. Collection of Lepidopterological Articles, IV.; Chronos-Press: Moscow, Russia, 2003; pp. 153–159. [Google Scholar]
- Korshunov, Y.P.; Ivonin, V.V. To the systematics of the blue butterfly genera Glaucopsyche and Celastrina. In Taxonomiya Nasekomykh i Gelmintov; Zolotarenko, G.S., Ed.; Nauka: Novosibirsk, Russia, 1990; pp. 68–79. ISBN 5-02-029542-6. [Google Scholar]
- Todisco, V.; Fiedler, K.; Puliga, E.; Dinca, V.; Voda, R.; Grill, A. The origin of the Sardinian Blue, Pseudophilotes barbagiae (Lepidoptera, Lycaenidae): Out-of-Europe or out-of-African? Genome 2015, 58 (Suppl. S5), 288. [Google Scholar]
- deWaard, J.R.; Ivanova, N.V.; Hajibabaei, M.; Hebert, P.D.N. Assembling DNA barcodes: Analytical protocols. In Environmental Genomics, Methods in Molecular Biology; Martin, C.C., Ed.; Humana Press: Totowa, NJ, USA, 2008; Volume 410, pp. 275–283, ISBN-13: 978-1588297778. [Google Scholar]
- Lukhtanov, V.A.; Sourakov, A.; Zakharov, E.V.; Hebert, P.D.N. DNA barcoding Central Asian butterflies: Increasing geographical dimension does not significantly reduce the success of species identification. Mol. Ecol. Resour. 2009, 9, 1302–1310. [Google Scholar] [CrossRef] [PubMed]
- Dinca, V.; Montagud, S.; Talavera, G.; Hernandez-Roldan, J.; Munguira, M.L.; Garcia-Barros, E.; Hebert, P.D.; Vila, R. DNA barcode reference library for Iberian butterflies enables a continental-scale preview of potential cryptic diversity. Sci. Rep. 2015, 5, 12395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abe, Y.; Miura, K.; Ito, H.; Yago, M.; Koh, S.-K.; Murata, K.; Yamashita, H. Origins of recently re-established and newly discovered populations of the endangered butterfly Shijimiaeoides divinus (Lepidoptera: Lycaenidae) in Oita Prefecture, Japan. Entomol. Sci. 2016, 19, 458–461. [Google Scholar] [CrossRef] [Green Version]
- Dapporto, L.; Cini, A.; Voda, R.; Dinca, V.; Wiemers, M.; Menchetti, M.; Magini, G.; Talavera, G.; Shreeve, T.; Bonelli, S.; et al. Integrating three comprehensive datasets shows that mitochondrial DNA variation is linked to species traits and paleogeographic events in European butterflies. Mol. Ecol. Resour. 2019, 19, 1623–1636. [Google Scholar] [CrossRef]
- Groza, B.; Vodă, R.; Székely, L.; Vila, R.; Dincă, V. Genetics and extreme confinement of three overlooked butterfly species in Romania call for immediate conservation actions. J. Insect. Conserv. 2021, 25, 137–146. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30 (Suppl. S4), 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misof, B.; Misof, K. A Monte Carlo approach successfully identifies randomness in multiple sequence alignments: A more objective means of data exclusion. Syst. Biol. 2009, 58, 21–34. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Lukhtanov, V.; Pierce, N.; Vila, R. DNA barcodes combined with multilocus data of representative taxa can generate reliable higher-level phylogenies. Syst. Biol. 2022, 71, 382–395. [Google Scholar] [CrossRef] [PubMed]
- Talavera, G.; Lukhtanov, V.A.; Pierce, N.E.; Vila, R. Establishing criteria for higher-level classification using molecular data: The systematics of Polyommatus blue butterflies (Lepidoptera, Lycaenidae). Cladistics 2013, 29, 166–192. [Google Scholar] [CrossRef] [PubMed]
- Pech, P.; Fric, Z.; Konvicka, M.; Zrzavy, J. Phylogeny of Maculinea blues (Lepidoptera: Lycaenidae) based on morphological and ecological characters: Evolution of parasitic myrmecophily. Cladistics 2004, 20, 362–375. [Google Scholar] [CrossRef] [PubMed]
- Wiemers, M.; Chazot, N.; Wheat, C.W.; Schweiger, O.; Wahlberg, N. A complete time-calibrated multi-gene phylogeny of the European butterflies. ZooKeys 2020, 938, 97–124. [Google Scholar] [CrossRef] [PubMed]
- Tshikolovets, V.V.; Kosterin, O.; Gorbunov, P.; Yakovlev, R. The Butterflies of Kazakhstan; Tshikolovets Publications: Pardubice, Czech Republic, 2016; 384p, ISBN 978-80-904900-7-9. [Google Scholar]
- Tshikolovets, V.V.; Yakovlev, R.V.; Bálint, Z. The Butterflies of Mongolia; Tshikolovets Publications: Pardubice, Czech Republic, 2009; 320p, ISBN 978-966-02-5145-8. [Google Scholar]
- Lukhtanov, V.A.; Lukhtanov, A.G. Rare and little-known species of blue butterflies (Lepidoptera, Lycaenidae) from the Southern Altai. Vestnik Zoologii 1988, 1988, 9–12. [Google Scholar]
- Gorbunov, P.Y. The Butterflies of Russia: Classification, Genitalia, Keys for Identification (Lepidoptera: Hesperioidea and Papilionoidea); Thesis: Ekaterinburg, Russia, 2001; 320p, ISBN 5-94131-004--8. [Google Scholar]
- Tuzov, V.K.; Bogdanov, P.V.; Churkin, S.V.; Dantchenko, A.V.; Devyatkin, A.L.; Murzin, V.S.; Samodurov, G.D.; Zhdanko, A.B. Guide to the Butterflies of Russia and Adjacent Territories (Lepidoptera, Rhopalocera); Pensoft: Moscow, Russia, 2000; Volume 2, 580p, ISBN 954-642-095-6. [Google Scholar]
- Bernardi, G. Note sur la variation géographique de l’armure génilale mâle des Iolana (Lep. Lycaenidae). Bull. Soc. Entomol. Fr. 1972, 77, 160–167. [Google Scholar] [CrossRef]
- Dumont, D. Révision du genre Iolana Bethune-Baker, 1914 (Lepidoptera: Lycaenidae). Description d’une nouvelle espèce: Kermani n.sp. Linneana Belg. 2004, 19, 332–357. [Google Scholar]
- Forster, W. Das System der paläarktischen Polyommatini (Lep. Lycaen.). Mitt. Münch. Ent. Ges. 1938, 28, 97–118. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | BOLD/Field ID | GeneBank | Gene | Country | Locality |
|---|---|---|---|---|---|
| Glaucopsyche alexis var. aeruginosa | BPALB161-16 | OP712325 | COI | Israel | Hermon |
| BPALB162-16 | OP712326 | Israel | Hermon | ||
| BPAL2627-14 | OP712327 | Israel | Beit Jan | ||
| BPAL2522-14 | OP712328 | Israel | Nahal Trivon | ||
| BPAL3276-16 | OP712334 | Kazakhstan | Dzhungarian Alatau, Kolbai | ||
| BPAL3274-16 | OP712332 | Kazakhstan | Kolbai | ||
| BPAL3275-16 | OP712333 | Kazakhstan | Kolbai | ||
| BPAL3408-16 | OP712338 | Kazakhstan | Kyzylagash | ||
| Glaucopsyche argali | LOWAM265-11 | OP712339 | COI | Kazakhstan | Kurtshum Mts, Salkyn-Cheku |
| LOWAM268-11 | OP712340 | Kazakhstan | Salkyn-Cheku | ||
| LOWAM267-11 | OP712341 | Kazakhstan | Salkyn-Cheku | ||
| LOWAM266-11 | OP712342 | Kazakhstan | Salkyn-Cheku | ||
| Glaucopsyche laetifica | BPAL3283-16 | OP712335 | COI | Kazakhstan | Ili valley, Koktal |
| BPAL3284-16 | OP712336 | Kazakhstan | Koktal | ||
| BPAL3285-16 | OP712337 | Kazakhstan | Koktal | ||
| Glaucopsyche melanops | BPAL3540-16 | OP712329 | COI | Morocco | Agadir 30.90 N 7.24 W |
| BPAL3541-16 | OP712330 | Morocco | Agadir | ||
| BPAL3546-16 | OP712331 | Morocco | Agadir | ||
| Iolana alfierii | BPAL2358-14 | OP712343 | COI | Israel | Avdat |
| BPAL2524-14 | OP712348 | Israel | Avdat | ||
| BPAL2525-14 | OP712349 | Israel | Avdat | ||
| BPAL2902-15 | OP712350 | Israel | Har-A-Negev | ||
| BPAL2359-14 | OP712352 | Israel | Avdat | ||
| Iolana andreasi | LOWAM286-11 | OP712351 | COI | Iran | Shahkuh |
| Iolana andreasi khayyami | BPAL2452-14 | OP712346 | COI | Iran | Tehran, Polur |
| BPAL2453-14 | OP712347 | Iran | Tehran, Polur | ||
| Iolana kermani | BPAL2450-14 | OP712344 | COI | Iran | Kerman, Kuh-e-Segoh |
| BPAL2451-14 | OP712345 | Iran | Kerman | ||
| Praephilotes anthracias | BPAL3279-16 | OP712323 | COI | Kazakhstan | Matai |
| BPAL3280-16 | OP712324 | Kazakhstan | Matai | ||
| Pseudophilotes abencerragus | BPALB525-18 | OP644300 | COI | Israel | |
| BPALB526-18 | OP644301 | Israel | |||
| BPALB527-18 | OP644302 | Israel | |||
| BPALB528-18 | OP644303 | Israel | |||
| BPAL3567-16 | OP644314 | Morocco | 32.5853 N 6.05611 W | ||
| Pseudophilotes bavius | BPALB030-16 | OP644305 | COI | Russia | Bashkortostan, 54.89 N 53.646 E |
| Pseudophilotes panope | L2-14 | OP679877 | ITS2 | Kazakhstan | Koibyn |
| L2-15 | OP679878 | Kazakhstan | Koibyn | ||
| L2-16 | OP679879 | Kazakhstan | Koibyn | ||
| L2-14 | OP681138 | Wingless | Kazakhstan | Koibyn | |
| L2-14 | OP681135 | EF1α | Kazakhstan | Koibyn | |
| L2-15 | OP681136 | Kazakhstan | Koibyn | ||
| L2-16 | OP681137 | Kazakhstan | Koibyn | ||
| L2-14 | OP678972 | 28S | Kazakhstan | Koibyn | |
| L2-15 | OP678973 | Kazakhstan | Koibyn | ||
| L2-16 | OP678974 | Kazakhstan | Koibyn | ||
| BPALB512-18 | OP644294 | COI | Kazakhstan | Koibyn | |
| BPALB513-18 | OP644295 | Kazakhstan | Koibyn | ||
| BPALB514-18 | OP644296 | Kazakhstan | Koibyn | ||
| BPALB515-18 | OP644297 | Kazakhstan | Koibyn | ||
| BPALB516-18 | OP644298 | Kazakhstan | Koibyn | ||
| BPALB517-18 | OP644299 | Kazakhstan | Koibyn | ||
| BPALB601-19 | OP644310 | Kazakhstan | 25 km NE Atyrau | ||
| BPALB602-19 | OP644311 | Kazakhstan | 21 km NE Atyrau | ||
| BPALB603-19 | OP644312 | Mongolia | Hovd, Arshantyn Nuru | ||
| BPAL3287-16 | OP644315 | Kazakhstan | Koibyn | ||
| Pseudophilotes vicrama | BPALB553-18 | OP644304 | COI | Israel | |
| BPALB284-17 | OP644306 | Tajikistan | |||
| BPALB331-17 | OP644307 | Tajikistan | |||
| BPALB359-17 | OP644308 | Tajikistan | |||
| BPALB470-17 | OP644309 | Israel | |||
| Pseudophilotes jacuticus | BPALB605-19 | OP644313 | COI | Russia | Yakutia, Yakutsk |
| Sequence | Total Length, bp | Number of Variable Sites | Number of Parsimony Informative Sites |
|---|---|---|---|
| COI | 1497 | 454 | 353 |
| COII | 679 | 184 | 116 |
| EF1a | 1161 | 238 | 157 |
| H3 | 327 | 57 | 42 |
| ITS2 | 449 | 104 | 81 |
| wg | 369 | 120 | 67 |
| 28S | 820 | 93 | 65 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lukhtanov, V.A.; Gagarina, A.V. Molecular Phylogeny and Taxonomy of the Butterfly Subtribe Scolitantidina with Special Focus on the Genera Pseudophilotes, Glaucopsyche and Iolana (Lepidoptera, Lycaenidae). Insects 2022, 13, 1110. https://doi.org/10.3390/insects13121110
Lukhtanov VA, Gagarina AV. Molecular Phylogeny and Taxonomy of the Butterfly Subtribe Scolitantidina with Special Focus on the Genera Pseudophilotes, Glaucopsyche and Iolana (Lepidoptera, Lycaenidae). Insects. 2022; 13(12):1110. https://doi.org/10.3390/insects13121110
Chicago/Turabian StyleLukhtanov, Vladimir A., and Anastasia V. Gagarina. 2022. "Molecular Phylogeny and Taxonomy of the Butterfly Subtribe Scolitantidina with Special Focus on the Genera Pseudophilotes, Glaucopsyche and Iolana (Lepidoptera, Lycaenidae)" Insects 13, no. 12: 1110. https://doi.org/10.3390/insects13121110