

Combined Jasmonic Acid and Ethylene Treatment Induces Resistance Effect in Faba Bean Plants Against Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing and Host Plant Cultivation
2.2. Different Concentrations of JA and ET Solutions
2.3. Plant Treatments
2.4. Oviposition Assays
2.5. Leaf Disc Thrip Selectivity Assays
2.6. Enzyme Activity Assays
2.7. Expression Levels of JA/ET Pathway Marker Genes
2.8. Life Table Study
2.9. Data Analysis
3. Results
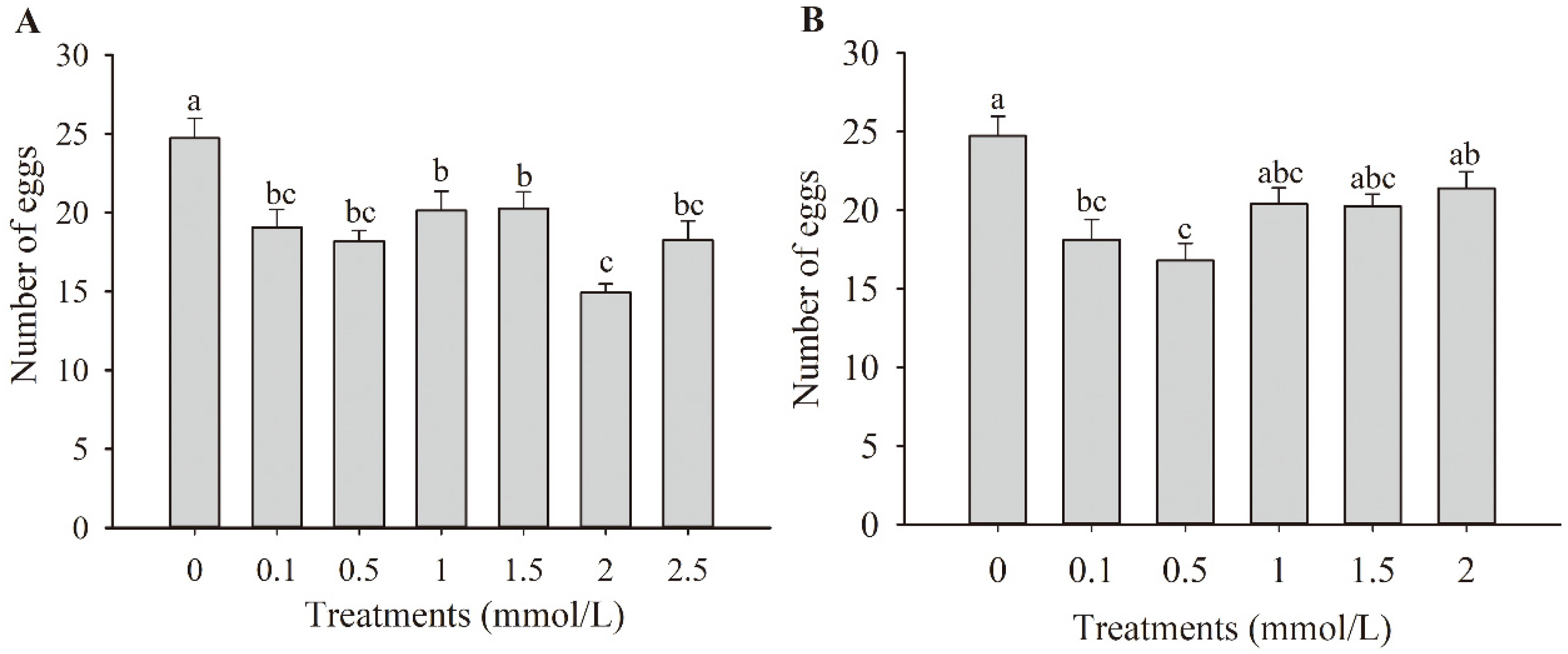
3.1. Effects of JA and ET Treatments on the Oviposition of F. occidentalis
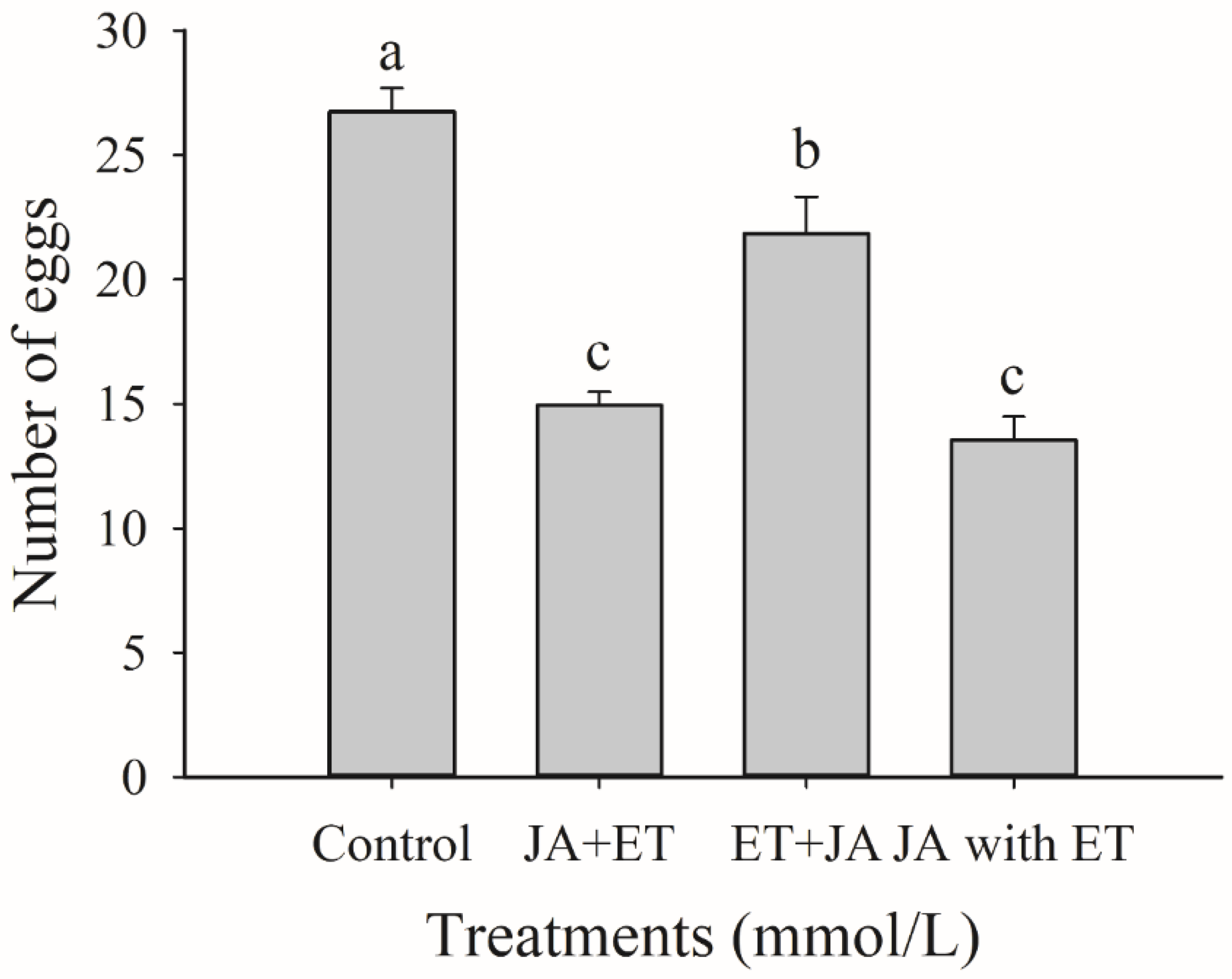
3.2. Effects of Combined JA and ET Treatments on the Oviposition of F. occidentalis
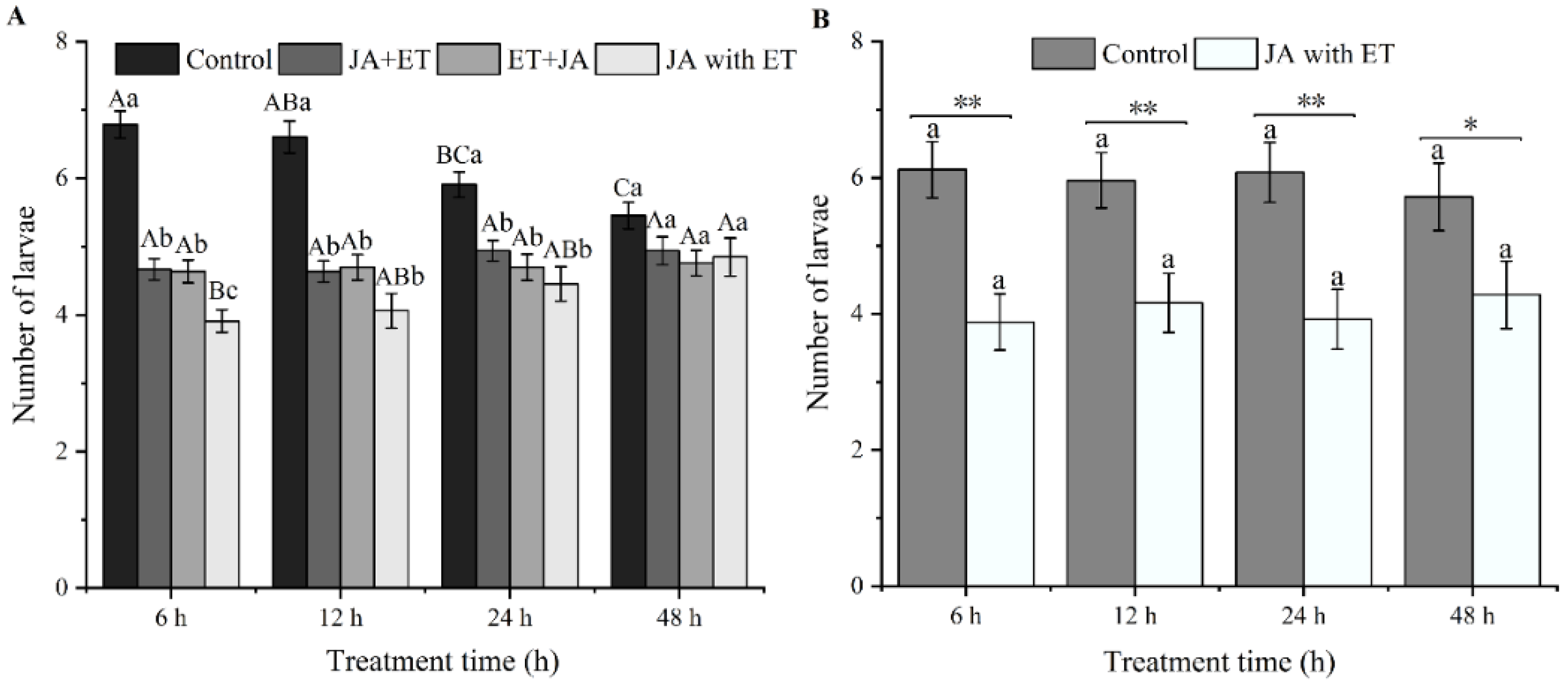
3.3. Effect of Combined JA and ET Treatment on the Oviposition Selectivity of F. occidentalis on Faba Bean
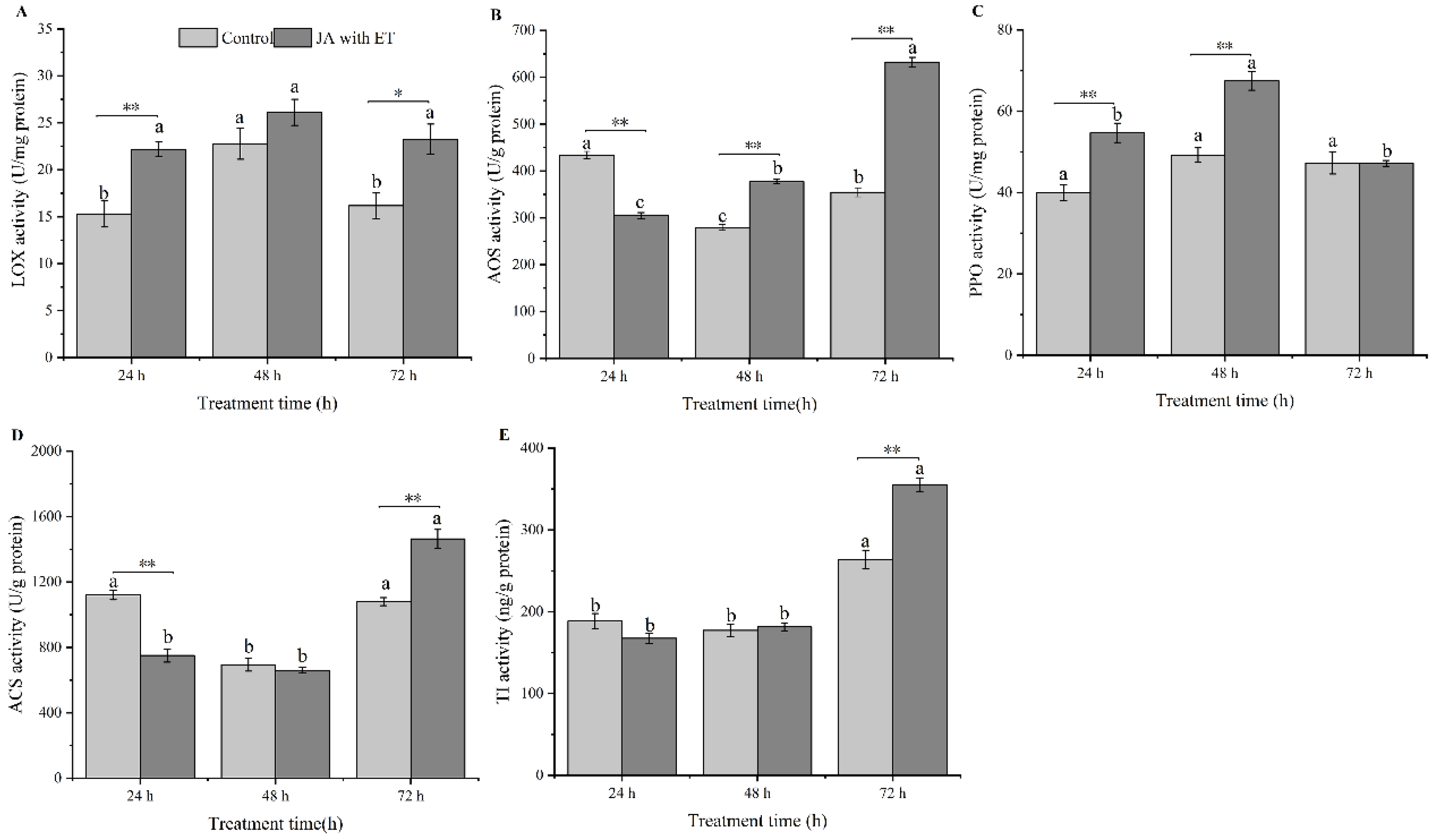
3.4. Effect of the JA with ET Treatment on the Activities of Defense Enzymes in Faba Bean Plants
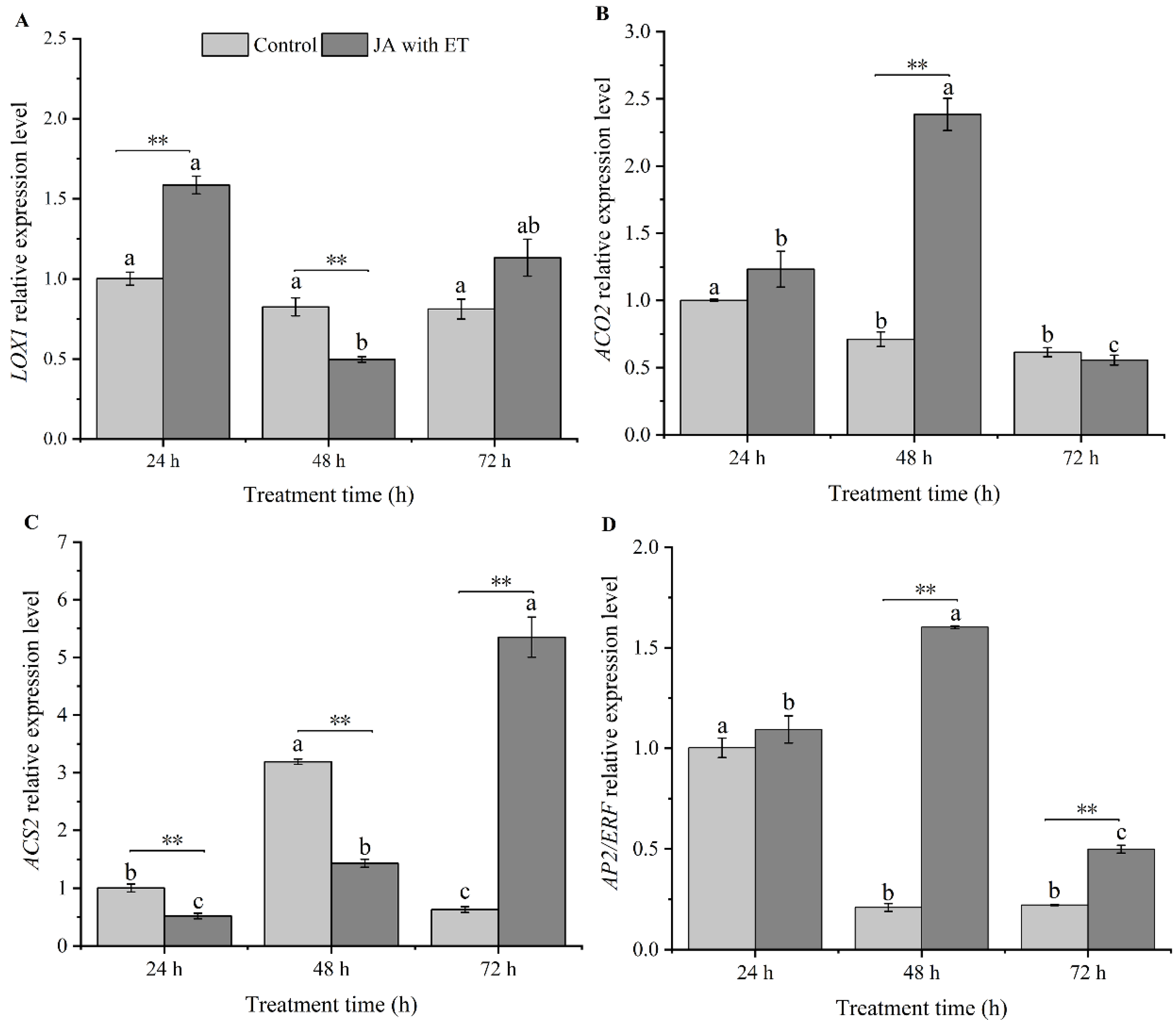
3.5. Effect of the JA with ET Treatment on the Expression of JA/ET Pathway Marker Genes
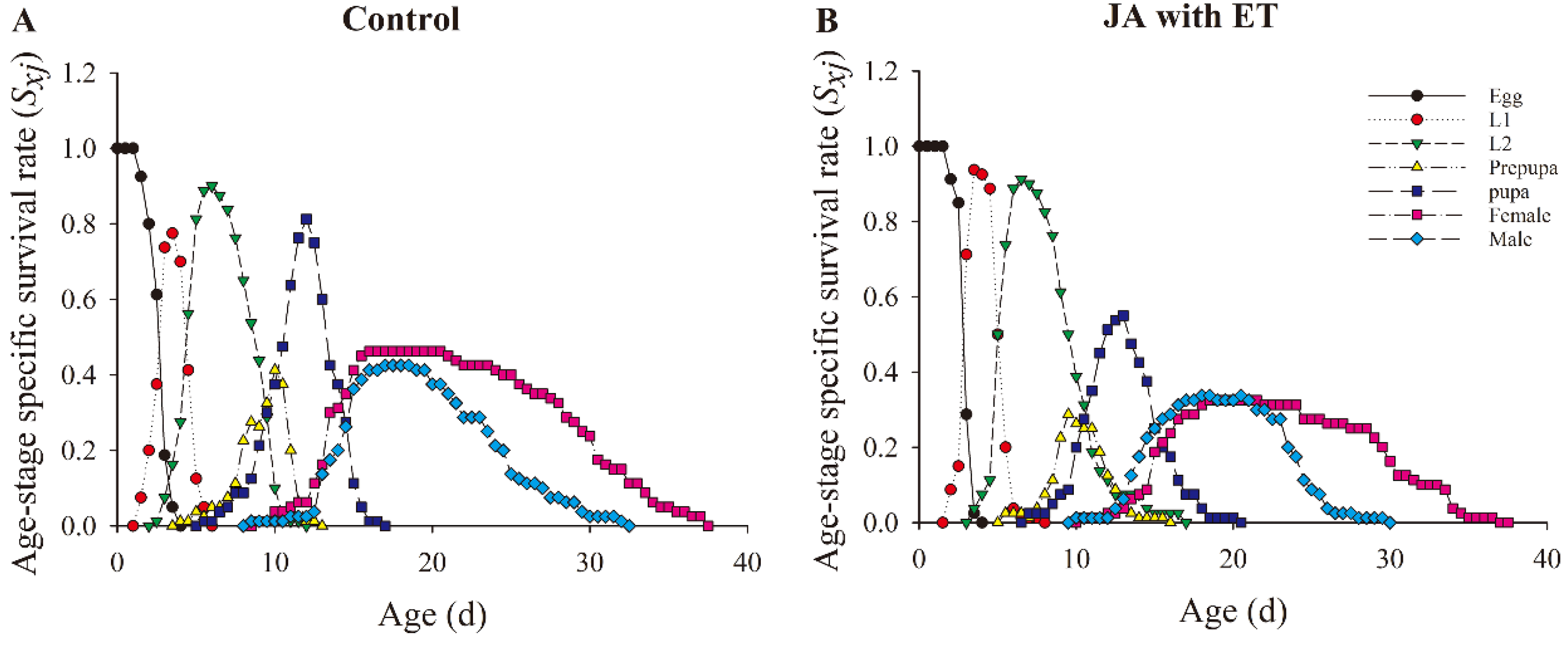
3.6. Effect of the JA with ET Treatment on the Life Parameters of F. occidentalis
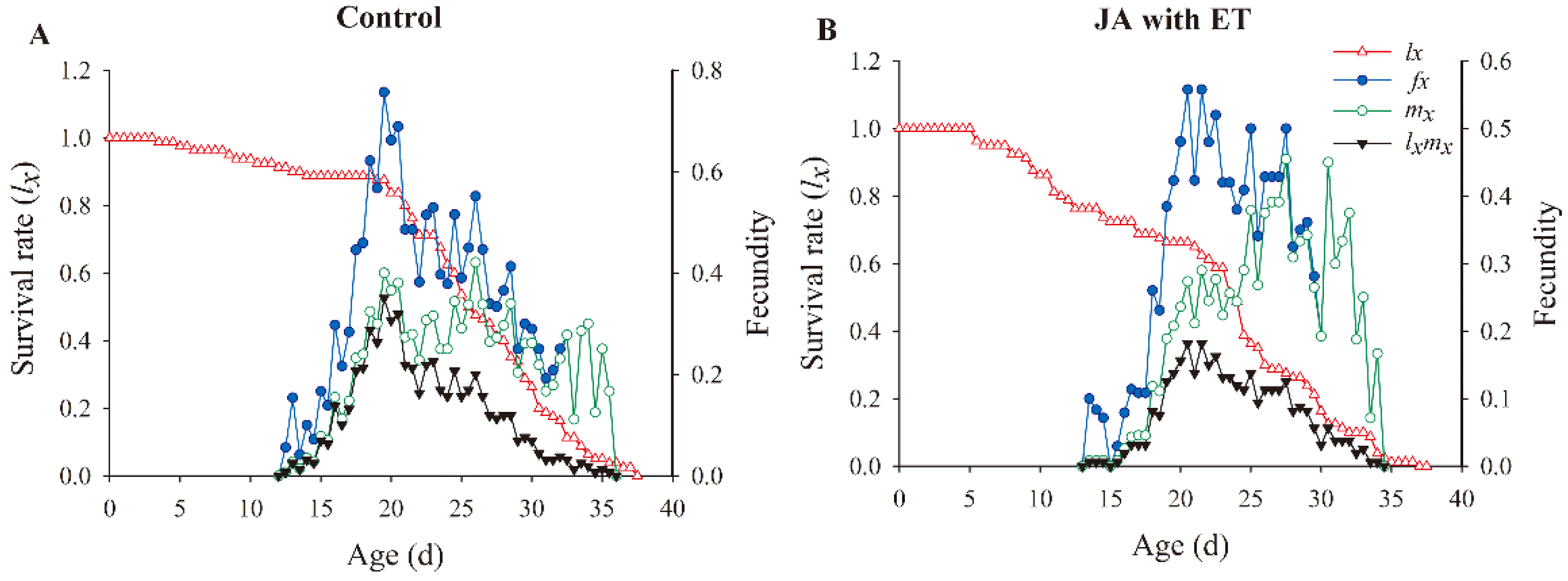
3.7. Effect of the JA with ET Treatment of Faba Bean Plants on the Survival Rate and Fecundity of F. occidentalis
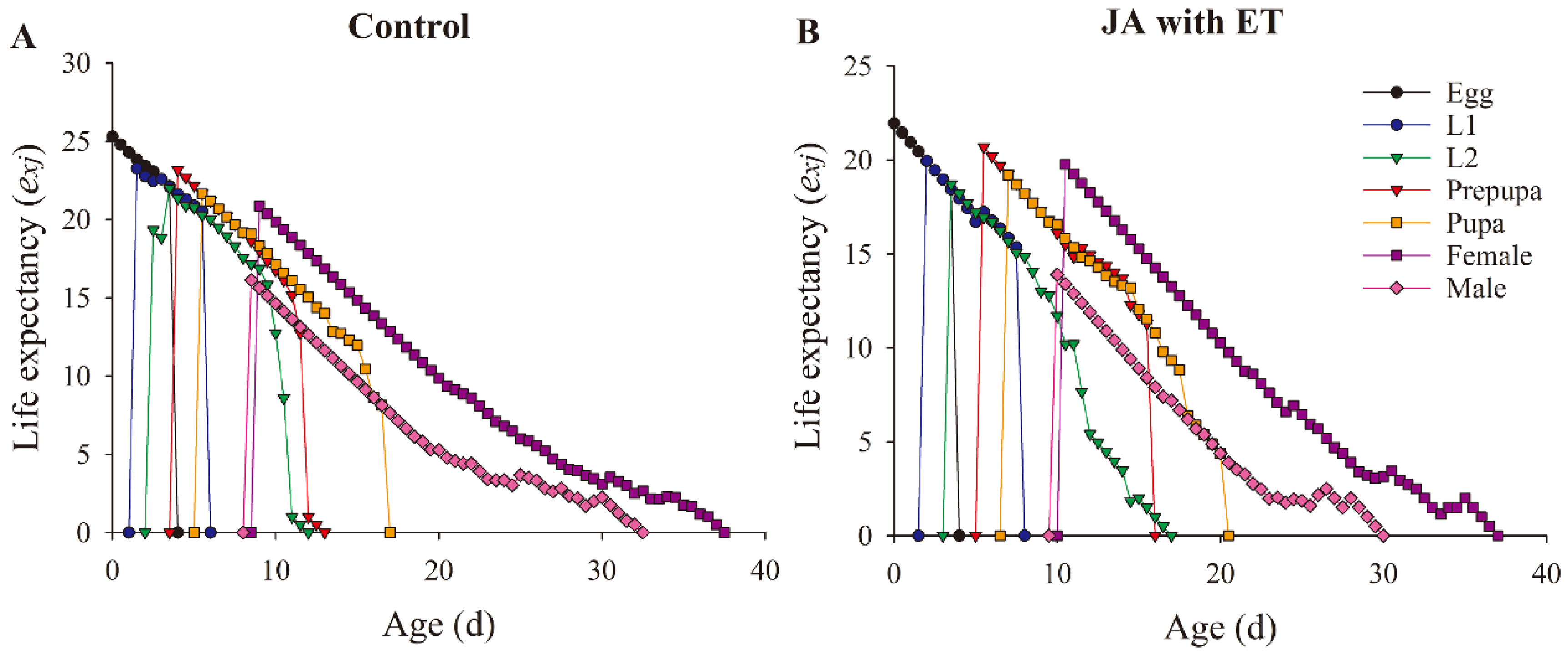
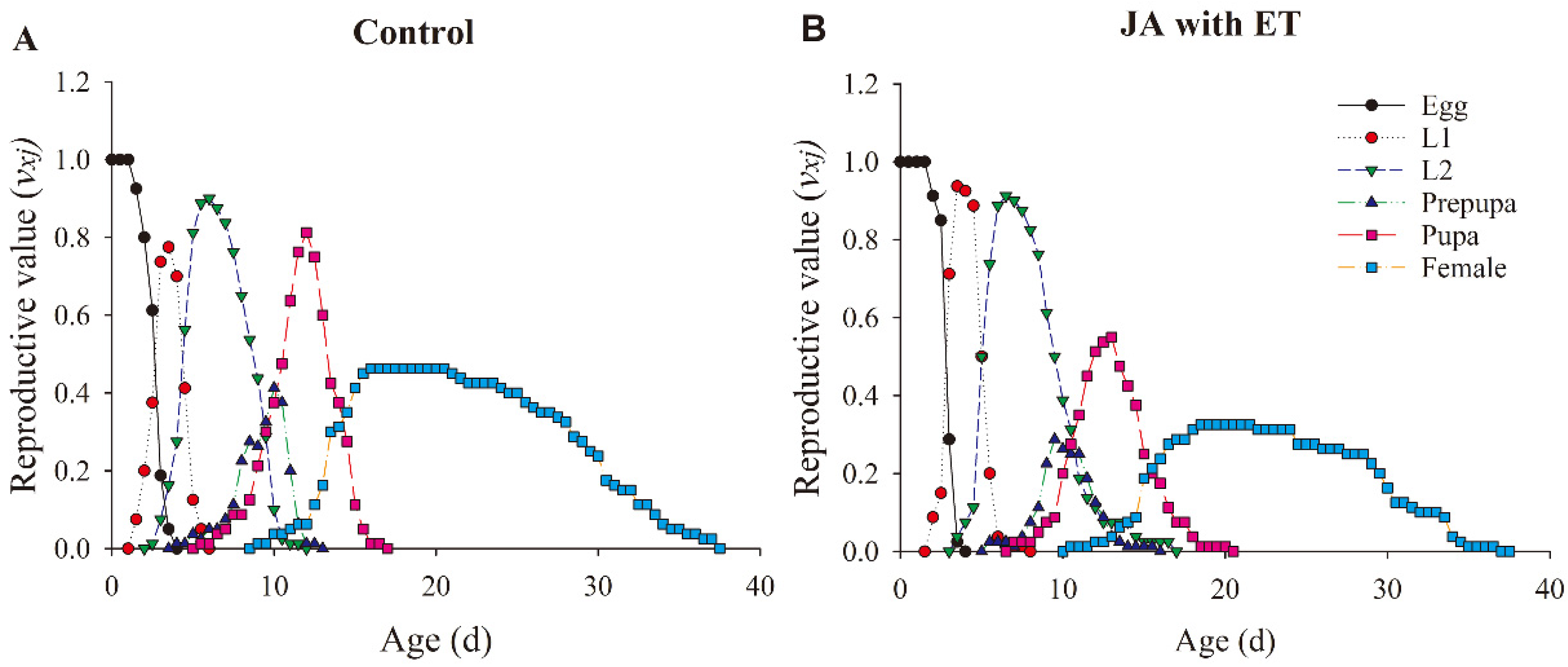
3.8. Effect of the JA with ET Treatment in Faba Bean Plants on the Life Parameters of F. occidentalis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kessler, A.; Baldwin, I.T. Plant responses to insect herbivory: The emerging molecular analysis. Annu. Rev. Plant Biol. 2002, 53, 299–328. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Chung, Y.R. Induction of systemic resistance against insect herbivores in plants by beneficial soil microbes. Front. Plant Sci. 2017, 8, 1816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef]
- Stella de Freitas, T.F.; Stout, M.J.; Sant’Ana, J. Effects of exogenous methyl jasmonate and salicylic acid on rice resistance to Oebalus pugnax. Pest Manag. Sci. 2019, 75, 744–752. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Lee, B. Friends or foes: New insights in jasmonate and ethylene co-actions. Plant Cell Physiol. 2015, 56, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Thaler, J.S.; Stout, M.J.; Karban, R.; Duffey, S.S. Exogenous jasmonates simulate insect wounding in tomato plants (Lycopersicon esculentum) in the laboratory and field. J. Chem. Ecol. 1996, 22, 1767–1781. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, L.; Yang, Y.; Guo, X.; Chen, G.; Xiong, X.; Dong, D.; Li, G. Transcriptome analysis reveals that exogenous ethylene activates immune and defense responses in a high late blight resistant potato genotype. Sci. Rep. 2020, 10, 21294. [Google Scholar] [CrossRef]
- Saniewski, M.; Dziurka, M.; Dziurka, K.; Góraj-Koniarska, J.; Ueda, J.; Miyamoto, K. Methyl jasmonate induces leaf senescence of Ginkgo biloba L.: Relevance to endogenous levels of plant hormones. Plant Growth Regul. 2020, 91, 383–396. [Google Scholar] [CrossRef]
- Hou, S.; Lin, L.; Lv, Y.; Xu, N.; Sun, X. Responses of lipoxygenase, jasmonic acid, and salicylic acid to temperature and exogenous phytohormone treatments in Gracilariopsis lemaneiformis (Rhodophyta). J. Appl. Phycol. 2018, 30, 3387–3394. [Google Scholar] [CrossRef]
- O’Donnell, P.J.; Calvert, C.; Atzorn, R.; Wasternack, C.; Leyser, H.M.O.; Bowles, D.J. Ethylene as a signal mediating the wound response of tomato plants. Science 1996, 274, 1914–1917. [Google Scholar] [CrossRef]
- Schaller, A.; Stintzi, A. Enzymes in jasmonate biosynthesis—Structure, function, regulation. Phytochemistry 2009, 70, 1532–1538. [Google Scholar] [CrossRef]
- Pattyn, J.; Vaughan-Hirsch, J.; Van de Poel, B. The regulation of ethylene biosynthesis: A complex multilevel control circuitry. New Phytol. 2021, 229, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.L.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell. 2002, 14, S131–S151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healey, M.A.; Senior, L.J.; Brown, P.H.; Duff, J. Relative abundance and temporal distribution of adult Frankliniella occidentalis (Pergande) and Frankliniella schultzei (Trybom) on French bean, lettuce, tomato and zucchini crops in relation to crop age. J. Asia-Pac. Entomol. 2017, 20, 859–865. [Google Scholar] [CrossRef]
- Zhang, B.; Qian, W.; Qiao, X.; Xi, Y.; Wan, F. Invasion biology, ecology, and management of Frankliniella occidentalis in China. Arch. Insect Biochem. Physiol. 2019, 102, e21613. [Google Scholar] [CrossRef]
- Reitz, S.R. Biology and ecology of the western flower thrips (Thysanoptera: Thripidae): The making of a pest. Fla. Entomol. 2009, 92, 7–13. [Google Scholar] [CrossRef]
- Zeng, G.; Zhi, J.R.; Ye, M.; Xie, W.; Zhang, T.; Li, D.Y.; Liu, L.; Wu, X.B.; Cao, Y. Life table and preference choice of Frankliniella occidentalis (Thysanoptera: Thripidae) for kidney bean plants treated by exogenous calcium. Insects 2021, 12, 838. [Google Scholar] [CrossRef]
- Lortzing, T.; Steppuhn, A. Jasmonate signalling in plants shapes plant–insect interaction ecology. Curr. Opin. Insect Sci. 2016, 14, 32–39. [Google Scholar] [CrossRef]
- Bera, S.; Blundell, R.; Liang, D.; Crowder, D.W.; Casteel, C.L. The oxylipin signaling pathway is required for increased aphid attraction and retention on virus-infected plants. J. Chem. Ecol. 2020, 46, 771–781. [Google Scholar] [CrossRef]
- Sarde, S.J.; Bouwmeester, K.; Venegas-Molina, J.; David, A.; Boland, W.; Dicke, M. Involvement of sweet pepper CaLOX2 in jasmonate-dependent induced defence against Western flower thrips. J. Integr. Plant Biol. 2019, 61, 1085–1098. [Google Scholar] [CrossRef]
- Kawazu, K.; Mochizuki, A.; Sugeno, W.; Seo, S.; Mitsuhara, I. Differences in the susceptibility of five herbivore species and developmental stages to tomato resistance induced by methyl jasmonate treatment. Arthropod-Plant Interact. 2013, 7, 415–422. [Google Scholar] [CrossRef]
- Abe, H.; Ohnishi, J.; Narusaka, M.; Seo, S.; Narusaka, Y.; Tsuda, S.; Kobayashi, M. Function of jasmonate in response and tolerance of Arabidopsis to thrip feeding. Plant Cell Physiol. 2008, 49, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Cong, C.L.; Zhi, J.R.; Liao, Q.R.; Mo, L.F. Effects of thrips feeding, mechanical wounding, and exogenous methyl salicylate and jasmonic acid on defense enzyme activities in kidney bean leaf. Acta Entomol. Sin. 2014, 57, 564–571. [Google Scholar] [CrossRef]
- Mou, F.; Zhi, J.R.; Cong, C.L.; Liao, Q.R.; Tian, T. Effects of exogenous jasmonic acid and methyl salicylate on developmental duration and population dynamics of Frankliniella occidentalis. Plant Prot. 2014, 40, 70–74. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y. A novel perspective on seed yield of broad bean (Vicia faba L.): Differences resulting from pod characteristics. Sci. Rep. 2014, 31, 6859. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Hou, X.L.; Yue, W.B.; Xie, W.; Zhang, T.; Zhi, J.R. Response of protective enzymes in Western flower thrips (Thysanoptera: Thripidae) to two leguminous plants. Environ. Entomol. 2020, 49, 1191–1197. [Google Scholar] [CrossRef]
- Soria, C.; Mollema, C. Life-history parameters of western flower thrips on susceptible and resistant cucumber genotypes. Entomol. Exp. Appl. 1995, 74, 177–184. [Google Scholar] [CrossRef]
- Gutierrez, N.; Giménez, M.J.; Palomino, C.; Avila, C.M. Assessment of candidate reference genes for expression studies in Vicia faba L. by real-time quantitative PCR. Mol. Breed. 2010, 28, 13–24. [Google Scholar] [CrossRef]
- Abid, G.; M’hamdi, M.; Mingeot, D.; Aouida, M.; Aroua, I.; Muhovski, Y.; Jebara, M. Effect of drought stress on chlorophyll fluorescence, antioxidant enzyme activities and gene expression patterns in faba bean (Vicia faba L.). Arch. Agron. Soil Sci. 2016, 63, 536–552. [Google Scholar] [CrossRef]
- Li, D.Y.; Zhi, J.R.; Yue, W.B.; Zhang, T.; Liu, L. Resistance to spinetoram affects host adaptability of Frankliniella occidentalis (Thysanoptera: Thripidae) based on detoxifying enzyme activities and an age-stage-two-sex life table. Environ. Entomol. 2022, 51, 780–789. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Acad. Sin. Bull. Inst. Zool. 1985, 24, 225–240. [Google Scholar]
- Chi, H.; You, M.; Atlihan, R.; Smith, C.; Kavousi, A.; Özgökçe, M.; Güncan, A.; Tuan, S.; Fu, J.; Xu, Y.; et al. Age-stage, two-sex life table: An introduction to theory, data analysis, and application. Entomol. Generalis 2020, 40, 103–124. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis; Visual BASIC Environment; National Chung Hsing University: Taichung, Taiwan, 2022. [Google Scholar]
- Chi, H. Periodic mass rearing and harvesting based on the theories of both the age-specific life table and the age-stage, two-sex life table. Environ. Entomol. 1994, 23, 535–542. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-Stage, Two-sex life tables of Aphidiusgifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzuspersicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef]
- Zhang, T.; Zhi, J.R.; Zeng, G.; Yue, W.B.; Liu, L. Review of induced plant defenses against the Western flower thrip and adaptations by thrips to counter these. Chin. J. Appl. Ecol. 2021, 58, 479–486. [Google Scholar] [CrossRef]
- Yang, J.; Duan, G.; Li, C.; Liu, L.; Han, G.; Zhang, Y.; Wang, C. The crosstalks between jasmonic acid and other plant hormone signaling highlight the involvement of jasmonic acid as a core component in plant response to biotic and abiotic stresses. Front. Plant Sci. 2019, 10, 1349. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Zhang, X.Y.; Sun, X.Z.; Xu, B.Y. Effect of methyl jasmonate on aphid resistance of chrysanthemum. Chin. J. Appl. Ecol. 2020, 31, 4197–4205. [Google Scholar] [CrossRef]
- Yang, S.Y.; Song, F.F.; Xie, J.C. Resistance of jasmonic acid-mediated cotton seedlings against the relative growth rate of Helicoverpa armiger. J. Northwest A F Univ. (Nat. Sci. Ed.). 2013, 41, 66–74. [Google Scholar] [CrossRef]
- Ma, F.; Yang, X.; Shi, Z.; Miao, X. Novel crosstalk between ethylene- and jasmonic acid-pathway responses to a piercing-sucking insect in rice. New Phytol. 2020, 225, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Schmelz, E.A.; Alborn, H.T.; Tumlinson, J.H. Synergistic interactions between volicitin, jasmonic acid and ethylene mediate insect-induced volatile emission in Zea mays. Physiol. Plant. 2003, 117, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Pangesti, N.; Reichelt, M.; van de Mortel, J.E.; Kapsomenou, E.; Gershenzon, J.; van Loon, J.J.; Dicke, M.; Pineda, A. Jasmonic acid and ethylene signaling pathways regulate glucosinolate levels in plants during rhizobacteria-induced systemic resistance against a leaf-chewing herbivore. J. Chem. Ecol. 2016, 42, 1212–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.Y.; Chen, J.L.; Cheng, D.F.; Sun, J.R.; Liu, Y.; Tian, Z. Biochemical and molecular characterizations of Sitobion avenae-induced wheat defense responses. Crop. Prot. 2009, 28, 435–442. [Google Scholar] [CrossRef]
- Koramutla, M.K.; Kaur, A.; Negi, M.; Venkatachalam, P.; Bhattacharya, R. Elicitation of jasmonate-mediated host defense in Brassica juncea (L.) attenuates population growth of mustard aphid Lipaphis erysimi (Kalt.). Planta 2014, 240, 177–194. [Google Scholar] [CrossRef]
- Xin, Z.; Li, X.; Li, J.; Chen, Z.; Sun, X. Application of chemical elicitor (Z)-3-hexenol enhances direct and indirect plant defenses against tea geometrid Ectropis obliqua. BioControl 2015, 61, 1–12. [Google Scholar] [CrossRef]
- Pre, M.; Atallah, M.; Champion, A.; De Vos, M.; Pieterse, C.M.; Memelink, J. The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol. 2008, 147, 1347–1357. [Google Scholar] [CrossRef] [Green Version]
- Kant, M.R.; Jonckheere, W.; Knegt, B.; Lemos, F.; Liu, J.; Schimmel, B.C.; Villarroel, C.A.; Ataide, L.M.; Dermauw, W.; Glas, J.J.; et al. Mechanisms and ecological consequences of plant defence induction and suppression in herbivore communities. Ann. Bot. 2015, 115, 1015–1051. [Google Scholar] [CrossRef]
- Bhonwong, A.; Stout, M.J.; Attajarusit, J.; Tantasawat, P. Defensive role of tomato polyphenol oxidases against cotton bollworm (Helicoverpa armigera) and beet armyworm (Spodoptera exigua). J. Chem. Ecol. 2008, 35, 28–38. [Google Scholar] [CrossRef]
- Divekar, P.A.; Rani, V.; Majumder, S.; Karkute, S.G.; Molla, K.A.; Pandey, K.K.; Behera, T.K.; Govindharaj, G.P.P. Protease inhibitors: An induced plant defense mechanism against herbivores. J Plant Growth Regul. 2022, 1–17. [Google Scholar] [CrossRef]
- Felton, G.W. Indigestion is a plant’s best defense. Proc. Natl. Acad. Sci. USA 2005, 102, 18771–18772. [Google Scholar] [CrossRef] [PubMed]
- Riley, D.G.; Joseph, S.V.; Srinivasan, R.; Diffie, S. Thrips vectors of tospoviruses. J. Integ. Pest Manag. 2011, 2, I1–I10. [Google Scholar] [CrossRef]
- Basu, S.; Clark, R.E.; Bera, S.; Casteel, C.L.; Crowder, D.W. Responses of pea plants to multiple antagonists are mediated by order of attack and phytohormone crosstalk. Mol. Ecol. 2021, 30, 4939–4948. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primers (5′-3′) | Reverse Primers (5′-3′) |
|---|---|---|
| ELF1A | GTGAAGCCCGGTATGCTTGT | CTTGAGATCCTTGACTGCAACATT |
| LOX1 | AGTCCTCAAGTTTCCGCCAC | GGGAGCGAATTAAGCCTGGA |
| ACO2 | GACGACAAAGTCAGTGGCCT | TCACTCGGTGCTCTATGCTT |
| ACS2 | CAGCCGCAAAATGTCGAGTT | AACCTCTTCGCACTCTCAGC |
| AP2/ERF | CACCGCCGTTTTCTATCTCC | TAACAACGGCAGCGTTTTCA |
| Index | Control | JA with ET | p | |||
|---|---|---|---|---|---|---|
| n | Mean ± SE | n | Mean ± SE | |||
| Egg stage (d) | 80 | 2.79 ± 0.07 | 80 | 3.04 ± 0.05 | 0.0034 | ** |
| First instar stage (d) | 80 | 1.73 ± 0.03 | 78 | 2.23 ± 0.06 | 0 | ** |
| Second instar stage (d) | 73 | 4.16 ± 0.11 | 58 | 4.42 ± 0.18 | 0.21 | ns |
| Prepupa stage (d) | 72 | 1.38 ± 0.03 | 57 | 1.46 ± 0.04 | 0.11 | ns |
| Pupa stage (d) | 71 | 3.68 ± 0.08 | 55 | 3.83 ± 0.10 | 0.22 | ns |
| Immature | 71 | 13.72 ± 0.19 | 55 | 14.91 ± 0.26 | 0.0002 | ** |
| Female adult stage (d) | 37 | 16.42 ± 0.64 | 26 | 15.08 ± 0.68 | 0.15 | ns |
| Male adult stage (d) | 34 | 10.60 ± 0.55 | 29 | 9.29 ± 0.36 | 0.047 | * |
| Per female oviposition | 37 | 12.39 ± 0.57 | 26 | 10.08 ± 0.67 | 0.0098 | ** |
| Adult preoviposition period (d) | 37 | 3.05 ± 0.10 | 26 | 3.37 ± 0.09 | 0.025 | * |
| Total preoviposition period (d) | 37 | 16.49 ± 0.28 | 26 | 18.56 ± 0.37 | 0 | ** |
| Female longevity (d) | 37 | 29.85 ± 0.70 | 26 | 30.27 ± 0.73 | 0.68 | ns |
| Male longevity (d) | 34 | 24.63 ± 0.59 | 29 | 23.95 ± 0.46 | 0.36 | ns |
| Sex ratio of offspring (%) | 929 | 1.05 ± 0.086 | 523 | 1.00 ± 0.96 | 0.79 | ns |
| Population Parameter | Control | JA with ET | p | |||
|---|---|---|---|---|---|---|
| n | Mean ± SE | n | Mean ± SE | |||
| Intrinsic rate of increase (r) (d1) | 80 | 0.08 ± 0.01 | 80 | 0.05 ± 0.01 | 0.003 | ** |
| Finite rate of increase (λ) (d1) | 80 | 1.08 ± 0.01 | 80 | 1.05 ± 0.01 | 0.003 | ** |
| Net reproductive rate (R0) | 80 | 5.73 ± 0.74 | 80 | 3.30 ± 0.57 | 0.009 | ** |
| Mean generation time (T) (d1) | 80 | 22.01 ± 0.37 | 80 | 23.77 ± 0.42 | 0.002 | ** |
| Gross reproductive rate (GRR) | 80 | 10.76 ± 1.03 | 80 | 9.20 ± 0.70 | 0.210 | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.-L.; Zhang, T.; Zhi, J.-R.; Tuo, L.; Yue, W.-B.; Li, D.-Y.; Liu, L. Combined Jasmonic Acid and Ethylene Treatment Induces Resistance Effect in Faba Bean Plants Against Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Insects 2022, 13, 1073. https://doi.org/10.3390/insects13111073
Jia Y-L, Zhang T, Zhi J-R, Tuo L, Yue W-B, Li D-Y, Liu L. Combined Jasmonic Acid and Ethylene Treatment Induces Resistance Effect in Faba Bean Plants Against Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae). Insects. 2022; 13(11):1073. https://doi.org/10.3390/insects13111073
Chicago/Turabian StyleJia, Yu-Lian, Tao Zhang, Jun-Rui Zhi, Lu Tuo, Wen-Bo Yue, Ding-Yin Li, and Li Liu. 2022. "Combined Jasmonic Acid and Ethylene Treatment Induces Resistance Effect in Faba Bean Plants Against Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae)" Insects 13, no. 11: 1073. https://doi.org/10.3390/insects13111073