
Newly Discovered Components of Dendrolimus pini Sex Pheromone
 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Collection of Female Volatiles
2.3. Chemical Analyses of Female Emissions
2.4. Chemicals, Standards, and Lure Components
| (Z5)-dodecen-1-yl acetate | (Z5)-12:OAc |
| (Z5)-tetradecen-1-yl acetate | (Z5)-14:OAc; |
| (Z5)-decenal | (Z5)-10:Ald |
| (Z5)-decen-1-yl acetate | (Z5)-10:OAc |
| (Z5)-dodecenal | (Z5)-12:Ald |
| (Z5)-dodecen-1-ol | (Z5)-12:OH |
| (Z5)-tetradecenal | (Z5)-14:Ald |
| (Z5)-tetradecen-1-ol | (Z5)-14:OH |
| (Z5,E7)-dodecadienal | (Z5,E7)-12:Ald |
| (E5,Z7)-dodecadienal | (E5,Z7)-12:Ald |
| (Z5,Z7)-dodecadienal | (Z5,Z7)-12:Ald |
| (E5,E7)-dodecadienal | (E5,E7)-12:Ald |
| (Z5,E7)-dodecadien-1-ol | (Z5,E7)-12:OH |
| (E5,Z7)-dodecadien-1-ol | (E5,Z7)-12:OH |
| (Z5,Z7)-dodecadien-1-ol | (Z5,Z7)-12:OH |
| (E5,E7)-dodecadien-1-ol | (E5,E7)-12:OH |
2.5. Steam Distillation of SPEO in a Deryng Apparatus
2.6. Identification of SPEO Components by GC/MS
2.7. Preparation of Lures and Dispensers
2.8. Wind Tunnel
2.9. Field Experiments
2.10. Statistical Analyses
3. Results
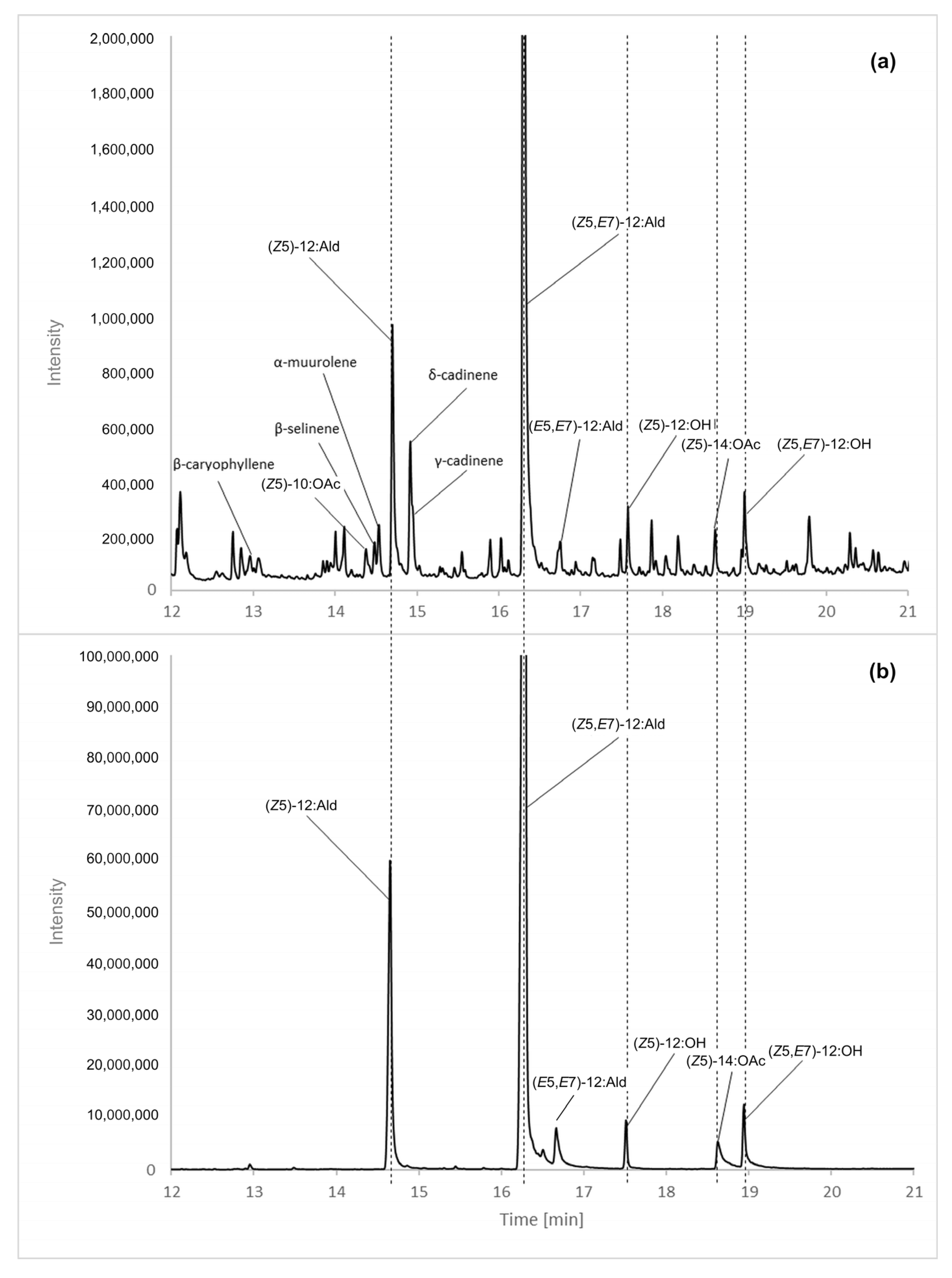
3.1. Identification of Female Volatiles
- (Z5)-10:OAc with: a diagnostic ion at m/z 138 [M-CH3COOH]+·, and a base peak at m/z 67 [C5H7]+;
- (Z5)-12:Ald with: a molecular ion at m/z 182 [M]+·, diagnostic ions at m/z 164 [M-H2O]+· and m/z 138 [M-C2H3O]+·, and a base peak at m/z 67 [C5H7]+;
- (Z5,E7)-12:Ald with: a molecular ion at m/z 180 [M]+·, diagnostic ions at m/z 162 [M-H2O]+· and m/z 137 [M-C2H3O]+·, a diagnostic ion for a 5,7-diene residue at m/z 123 [C9H15]+· [34], and a base peak at m/z 79 [C6H7]+;
- (E5,E7)-12:Ald with: a molecular ion at m/z 180 [M]+·, diagnostic ions at m/z 162 [M-H2O]+· and m/z 137 [M-C2H3O]+·, a diagnostic ion for the presence of a 5,7-diene residue at m/z 123 [C9H15]+·, and a base peak at m/z 79 [C6H7]+;
- (Z5)-12:OH with a base peak at m/z 67 [C5H7]+;
- (Z5)-14:OAc with: a hardly detectable cation m/z 255 [M+H]+, a diagnostic ion at m/z 194 [M-CH3COOH]+· and a base peak at m/z 67 [C5H7]+; and

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rt min | Compound | KI Calculated | KI from Literature |
|---|---|---|---|
| 12.95 | β-caryophyllene | 1592 | 1594 [42] |
| 14.38 | (Z5)-10:OAc | 1723 | no data |
| 14.48 | β-selinene | 1718 | 1711 [42] |
| 14.54 | α-muurolene | 1724 | 1714 [42] 1725 [43] |
| 14.70 | (Z5)-12:Ald | 1739 | 1724 [44] |
| 14.92 | δ-cadinene | 1760 | 1749 [42] |
| 14,95 | γ-cadinene | 1763 | 1752 [42] |
| 16.30 | (Z5,E7)-12:Ald | 1883 | 1884 [45] |
| 16.75 | (E5,E7)-12:Ald | 1925 | 1925 [45] |
| 17.58 | (Z5)-12:OH | 2007 | 2007 [44] |
| 18.64 | (Z5)-14:OAc | 2121 | 2103 [44] |
| 19.01 | (Z5,E7)-12:OH | 2153 | 2161 [45] |
3.2. Wind-Tunnel Experiments
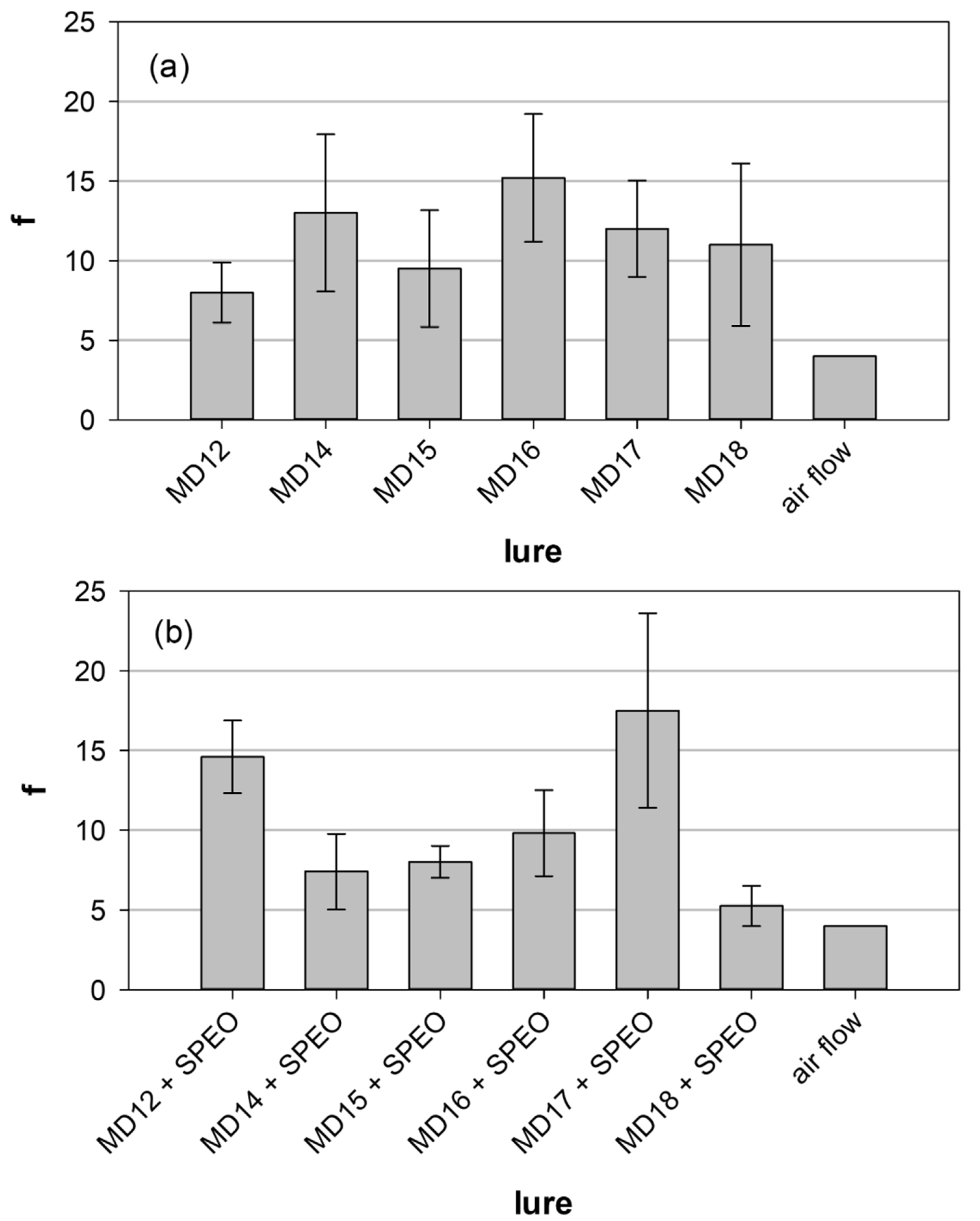
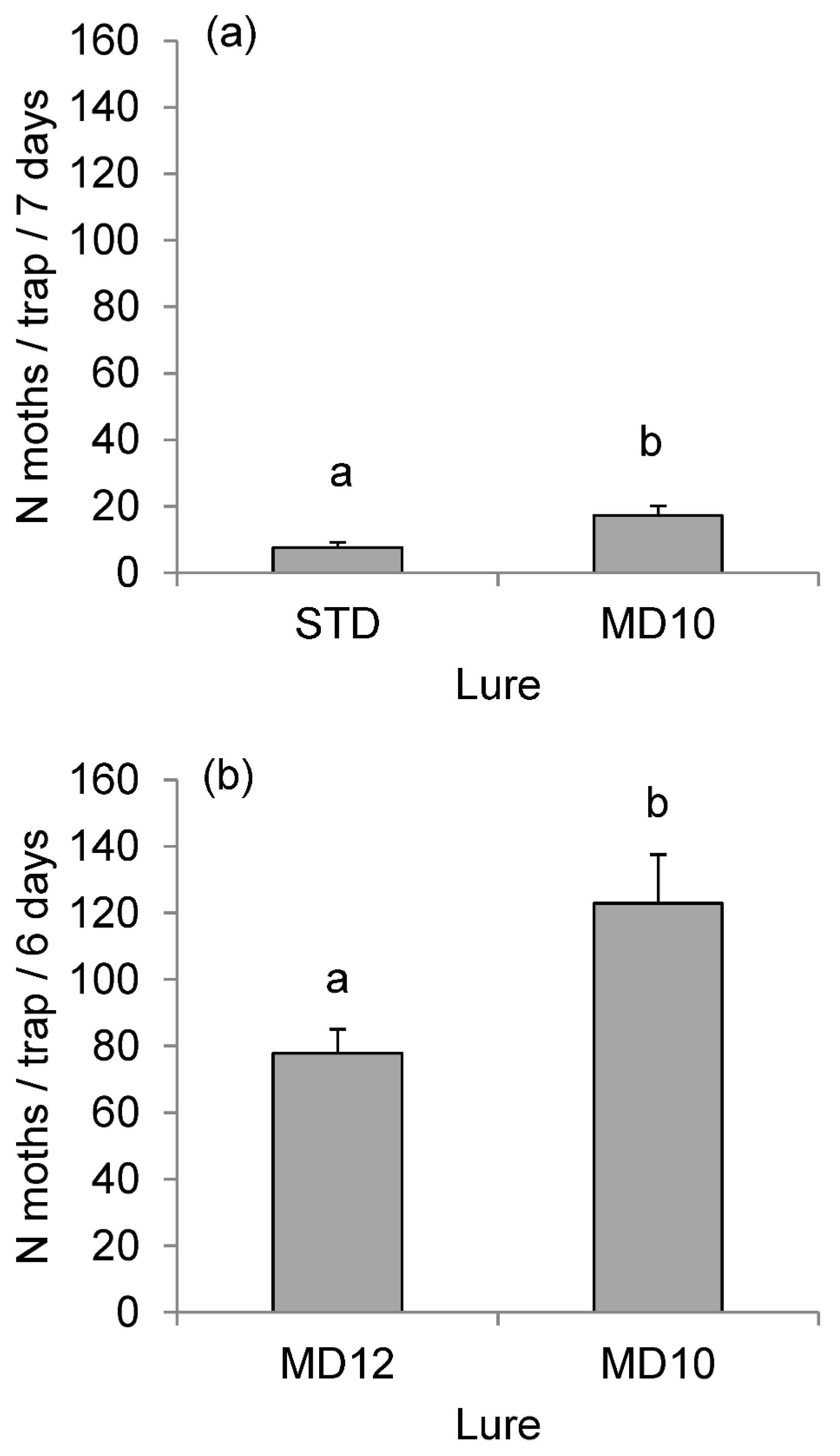
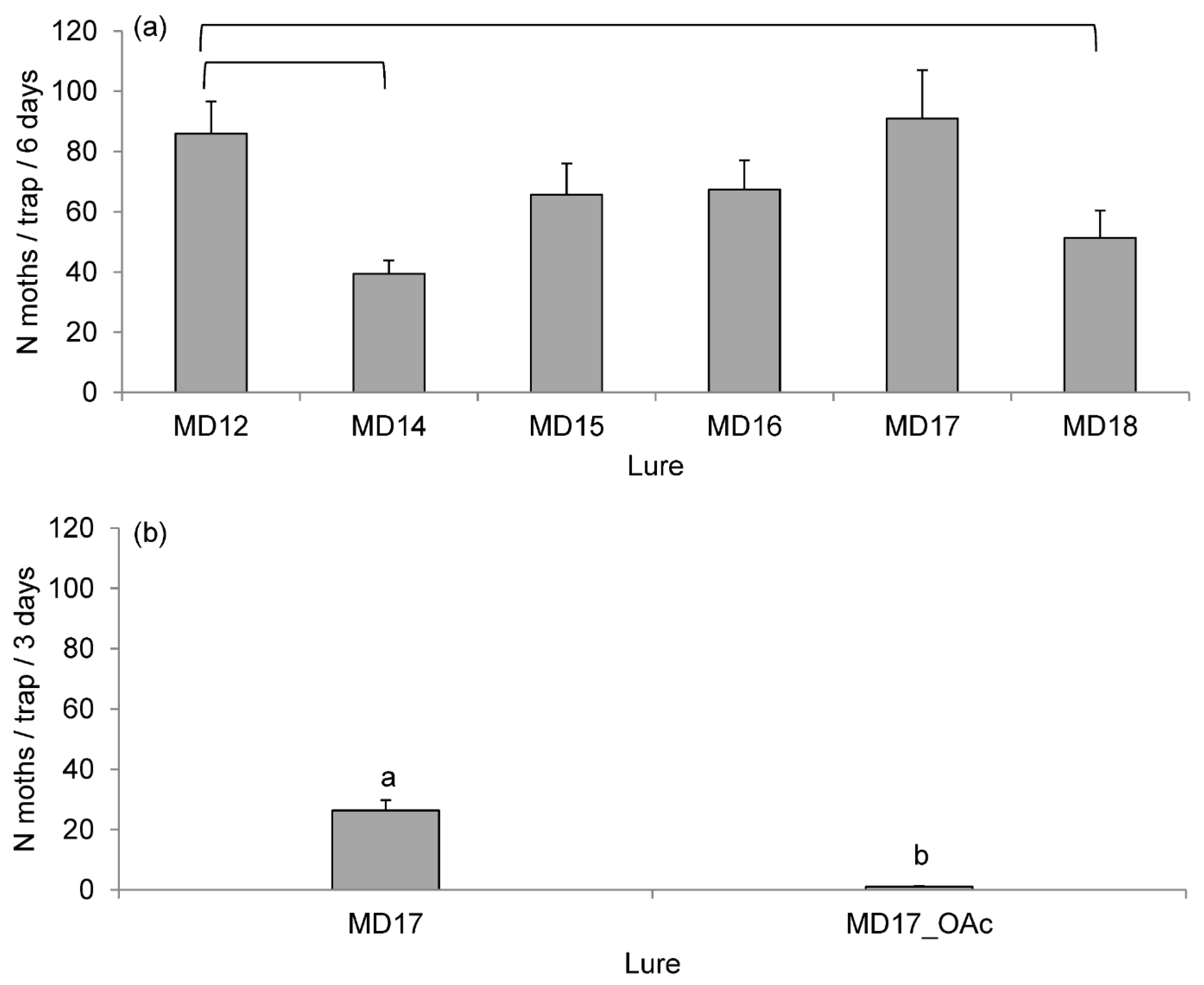
3.3. Field Experiments
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- US Department of Agriculture’s Animal and Plant Health Inspection Service. Plant Protection and Quarantine. In New Pest Response Guidelines: Dendrolimus Pine Moths; US Department of Agriculture’s Animal and Plant Health Inspection Service: Washington, DC, USA, 2012. [Google Scholar]
- Skrzecz, I.; Ślusarski, S.; Tkaczyk, M. Integration of science and practice for Dendrolimus pini (L.) management—A review with special reference to Central Europe. Forest. Ecol. Manag. 2020, 455, 117697. [Google Scholar] [CrossRef]
- LaForest, J.H.; Douce, G.K.; Minteer, C.R.; Bargeron, C.T.; Evans, C.W.; Moorhead, D.J.; Sanders, C. Invasive Insects of Concern to Georgia; University of Georgia, Center for Invasive Species and Ecosystem Health: Athens, GA, USA, 2008. [Google Scholar]
- Molet, T. CPHST Pest Datasheet for Dendrolimus pini. 2012. Available online: http://pest.ceris.purdue.edu/services/approvedmethods/sheet.php?v=1966&from=2023 (accessed on 15 November 2022).
- Sierpińska, A. Towards an Integrated Management of Dendrolimus pini L.; US Department of Agriculture, Forest Service, Northeastern Research Station: Newtown Square, PA, USA, 1998; pp. 129–142.
- Lesniak, A. Climatic and meteorological conditions of the pine moth (Dendrolimus pini L.) outbreaks. Ekol. Pol. 1976, 24, 515–547. [Google Scholar]
- PGLLP. Instrukcja Ochrony Lasu Tom II; Centrum Informacyjne Lasów Państwowych: Warszawa, Poland, 2012.
- Zhang, A.-B.; Wang, Z.-J.; Tan, S.-J.; Li, D.-M. Monitoring the masson pine moth, Dendrolimus punctatus (Walker) (Lepidoptera: Lasiocampidae) with synthetic sex pheromone-baited traps in Qianshan County, China. Appl. Entomol. Zool. 2003, 38, 177–186. [Google Scholar] [CrossRef]
- Bestmann, H.J.; Koschatzky, K.H.; Platz, H.; Süß, J.; Vostrowsky, O.; Knauf, W.; Burghardt, G.; Schneider, I. Pheromone, XL. Stereoselektive synthese des Pheromonkomplexes von Lasio-campidae-Arten (Lepidoptera); ein Sexuallockstoff für den Kiefernspinner Dendrolimus pini. Liebigs Ann. Chem. 1982, 1982, 1359–1365. [Google Scholar] [CrossRef]
- Priesner, E.; Bogenschütz, H.; Albert, R.; Reed, D.W.; Chisholm, M.D. Identification and field evaluation of a sex pheromone of the European pine moth. Z. Naturforsch C 1984, 39, 1192–1195. [Google Scholar] [CrossRef] [Green Version]
- Kovalev, B.G.; Bolgar, T.S.; Zubov, P.A.; Zharkov, D.G.; Golosova, M.; Nesterov, E.A.; Tvaradze, M.S. Identification of additional components of the sex pheromone of Dendrolimus pini. Chem. Nat. Compd. 1993, 29, 135–136. [Google Scholar] [CrossRef]
- Baranchikov, Y.N.; Petko, V.M.; Ponomarev, V.L. The Russians are coming—Aren’t they? Siberian moth in European forests. In Proceedings of the 17th U.S. Department of Agriculture Interagency Research Forum on Gypsy Moth and Other Invasive Species 2006; Gen. Tech. Rep. NRS-P-10. US Department of Agriculture: Newtown Square, PA, USA, 2007; pp. 18–20. [Google Scholar]
- Johansson, B.G.; Anderbrant, O.; Sierpinski, A. Multispecies trapping of six pests of scots pine in Sweden and Poland. J. Appl. Entomol. 2002, 126, 212–216. [Google Scholar] [CrossRef]
- Ostrauskas, H.; Ivinskis, P. Moths caught in pheromone traps during search for Dendrolimus pini and D. sibiricus (Lepidoptera, Lasiocampidae) in Lithuania. Acta Zool Lith. 2011, 21, 239–243. [Google Scholar] [CrossRef]
- Ponomarev, B.L.; Baranchikov, Y.N.; Marchenko, Y.I.; Ostrauskas, H.H. Field testing of Dendrolimus pini L. sex pheromone. Lesnoy Vestnik 2006, 2, 89–91. [Google Scholar]
- Saunders, D.S. Insect photoperiodism: Seeing the light. Physiol. Entomol. 2012, 37, 207–218. [Google Scholar] [CrossRef]
- Kováts, E.; Keulemans, A.I.M. The Kováts retention index system. Anal. Chem. 1964, 36, 31A–41A. [Google Scholar] [CrossRef]
- Zenkevich, I.G. Kovats’ retention index system. In Encyclopedia of Chromatography, 3rd ed.; Cazes, J., Ed.; CRC Press, Taylor and Francis: Boca Raton, FL, USA, 2010; pp. 1304–1310. [Google Scholar]
- Stille, J.K.; Groh, B.L. Stereospecific cross-coupling of vinyl halides with vinyl tin reagents catalyzed by palladium. J. Am. Chem. Soc. 1987, 109, 813–817. [Google Scholar] [CrossRef]
- Stille, J.K.; Simpson, J.H. Stereospecific palladium-catalyzed coupling reactions of vinyl iodides with acetylenic tin reagents. J. Am. Chem. Soc. 1987, 109, 2138–2152. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Ioannou, E.; Koutsaviti, A.; Tzakou, O.; Roussis, V. The genus Pinus: A comparative study on the needle essential oil composition of 46 pine species. Phytochem. Rev. 2014, 13, 741–768. [Google Scholar] [CrossRef]
- Orav, A.; Kailas, T.; Liiv, M.; Aav, R. Capillary gas chromatographic analysis of the monoterpenoic fracuion of Estonian conifer needle oil. Proc. Estonian Acad. Sci. Chem. 1995, 44, 149–155. [Google Scholar]
- Venskutonis, P.R.; Vyskupaityte, K.; Plausinaitis, R. Composition of essential oils of Pinus sylvestris L. from different locations of Lithuania. J. Essent Oil Res. 2000, 12, 559–565. [Google Scholar] [CrossRef]
- Sukovata, L.; Asztemborska, M.; Rudziński, K.J.; Cieślak, M.; Staszek, D.; Janiszewski, W.; Szmigielski, R.; Kolk, A.; Raczko, J. Effect of dispenser type, trap design and placement on catches of pine-tree lappet moth, Dendrolimus pini. Phytoparasitica 2020, 48, 63–74. [Google Scholar] [CrossRef]
- Bolker, B.; Brooks, M.; Clark, C.; Geange, S.; Poulsen, J.; Stevens, H.; White, J.-S. Generalized Linear Mixed Models: A Practical Guide for Ecology and Evolution. Trends Ecol. Evol. 2009, 24, 127–135. [Google Scholar] [CrossRef]
- Mangiafico, S. Summary and Analysis of Extension Program Evaluation in R; Rutgers Cooperative Extension: New Brunswick, NJ, USA, 2016. [Google Scholar]
- Zuur, A.; Ieno, E.; Walker, N.; Saveliev, A.; Smith, G. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 10 October 2022).
- RStudio Team. RStudio: Integrated Development for R. RStudio Inc.; RStudio Team: Boston, MA, USA, 2015. [Google Scholar]
- Brooks, M.; Kristensen, K.; van Benthem, K.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.; Mächler, M.; Bolker, B. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Estimated Marginal Means, Aka Least-Squares Means. v. 1.4.5, R Package (>=3.2). 2020. Available online: https://zbmath.org/software/29805 (accessed on 10 October 2022).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.2.7. Available online: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 6 February 2020).
- Ando, T.; Yamakawa, R. Analyses of lepidopteran sex pheromones by mass spectrometry. Trend Anal. Chem. 2011, 30, 990–1002. [Google Scholar] [CrossRef]
- Ando, T.; Katagiri, Y.; Uchiyama, M. Mass spectra of dodecadienic compounds with a conjugated double bond, lepidopterous sex pheromones. Agric. Biol. Chem. Tokyo 1985, 49, 413–421. [Google Scholar] [CrossRef]
- Rotundo, G.; Germinara, G.; Cristofaro, A. Chemical, electrophysiological, and behavioral investigations on the sex pheromone of lackey moth, Malacosoma neustrium. J. Chem. Ecol. 2004, 30, 2057–2069. [Google Scholar] [CrossRef] [PubMed]
- Vu, M.H.; Ando, T.; Takahashi, N.; Tatsuki, S.; Yamane, A.; Ikeda, T.; Yamazaki, S. Identification of the female sex pheromone of the pine moth. Agric. Biol. Chem. Tokyo 1980, 44, 231–233. [Google Scholar] [CrossRef]
- Blomquist, G.J.; Vogt, R.G. Biosynthesis and detection of pheromones and plant volatiles—Introduction and overview. In Insect Pheromone Biochemistry and Molecular Biology; Blomquist, G.J., Vogt, R.G., Eds.; Academic Press: London, UK, 2003; pp. 3–18. [Google Scholar]
- Bruyne, M.; Baker, T.C. Odor detection in insects: Volatile codes. J. Chem. Ecol. 2008, 34, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Sukovata, L.; Kolk, A.; Jaroszynska, J.; Krajewska, U.; Purzynska, A.; Isidorov, V. Host-tree preferences of the pine moth (Lepidoptera: Lasiocampidae) and pine beauty moth (Lepidopera: Noctuidae) larvae in relation to needle quality. In Proceedings of the Ecology, Survey and Management of Forest Insects, Krokow, Poland, 1–5 September 2003; pp. 98–106. [Google Scholar]
- Szmigielski, R.; Cieslak, M.; Rudziński, K.; Maciejewska, B. Identification of volatiles from Pinus silvestris attractive for Monochamus galloprovincialis using a SPME-GC/MS platform. Environ. Sci. Pollut. R 2012, 19, 2860–2869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, T.Y.; Eiserich, J.P.; Shibamoto, T. Volatile compounds isolated from edible Korean chamchwi (Aster scaber Thunb). J. Agric. Food Chem. 1993, 41, 1693–1697. [Google Scholar] [CrossRef]
- Vernin, G.; Boniface, C.; Metzger, J.; Ghiglione, C.; Hammoud, A.; Suon, K.-N.; Fraisse, D.; Párkányi, C. GC-MS-SPECMA bank analysis of JUniperus communis needles and berries. Phytochemistry 1988, 27, 1061–1064. [Google Scholar] [CrossRef]
- Marques, F.d.A.; McElfresh, J.S.; Millar, J.G. Kováts retention indexes of monounsaturated C12, C14, and C16 alcohols, acetates and aldehydes commonly found in lepidopteran pheromone blends. J. Brazil Chem. Soc. 2000, 11, 592–599. [Google Scholar] [CrossRef]
- Kong, X.; Zhang, Z.; Zhao, C.; Wang, H. Female sex pheromone of the yunnan pine caterpillar moth Dendrolimus Houi: First (E,Z)-isomers in pheromone components of Dendrolimus spp. J. Chem. Ecol. 2007, 33, 1316–1327. [Google Scholar] [CrossRef]
- Priesner, E.; Altenkirch, W.; Baltensweiler, W.; Bogenschütz, H. Evaluation of (Z)-5-decen-1-ol as an attractant for male larch casebearer moths, Coleophora laricella. Z. Naturforsch C 1982, 37, 953. [Google Scholar] [CrossRef]
- Witzgall, P.; Priesner, E. Wind-tunnel study on attraction inhibitor in male Coleophora laricella Hbn. (Lepidoptera: Coleophoridae). J. Chem. Ecol. 1991, 17, 1355–1362. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-D.; Löfstedt, C. Functional evolution of a multigene family: Orthologous and paralogous pheromone receptor genes in the turnip moth, Agrotis segetum. PLoS ONE 2013, 8, e77345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Löfstedt, C.; Linn, C.E.; Löfqvist, J. Behavioral responses of male turnip moths,Agrotis segetum, to sex pheromone in a flight tunnel and in the field. J. Chem. Ecol. 1985, 11, 1209–1221. [Google Scholar] [CrossRef]
- Beevor, P.S.; Youm, O.; Hall, D.; Cork, A. Identification and field evaluation of components of female sex pheromone of millet stem borer, Coniesta ignefusalis. J. Chem. Ecol. 1999, 25, 2643–2663. [Google Scholar] [CrossRef] [Green Version]
- Capizzi, A.; Tonini, C.; Arsura, E.; Guglielmetti, G.; Massardo, P.; Piccardi, P. Sex pheromone components of the European goat moth,Cossus cossus. J. Chem. Ecol. 1983, 9, 191–200. [Google Scholar] [CrossRef]
- Bestmann, H.J.; Brosche, T.; Koschatzky, K.H.; Michaelis, K.; Platz, H.; Vostrowsky, O.; Knauf, W. Pheromone XXX. Identifizierung eines neuartigen Pheromonkomplexes aus der Graseule Scotia exclamationis (Lepidoptera). Tetrahedron Lett. 1980, 21, 747–750. [Google Scholar] [CrossRef]
- Struble, D.L.; Arn, H. Combined gas chromatography and electroantennogram recording of insect olfactory responses. In Techniques in Pheromone Research; Hummel, H.E., Miller, T.A., Eds.; Springer: New York, NY, USA, 1984; pp. 161–178. [Google Scholar]
- Arn, H.; Städler, E.; Rauscher, S.; Buser, H.R.; Mustaparta, H.; Esbjerg, P.; Zethner, O.; Philipsen, H.; Struble, D.L.; Bues, R. Multicomponent sex pheromone in Agrotis segetum: Preliminary analysis and field evaluation. Z. Naturforsch C 1980, 35, 986. [Google Scholar] [CrossRef]
- Bestmann, H.J.; Vostrowsky, O.; Koschatzky, K.H.; Platz, H.; Brosche, T.; Kantardjiew, I.; Rheinwald, M.; Knauf, W. (Z)-5-Decenylacetat, ein Sexuallockstoff für Männchen der Saateule Agrotis segetum (Lepidoptera). Angew. Chem. 1978, 90, 815–816. [Google Scholar] [CrossRef]
- Wu, W.; Bengtsson, M.; Hansson, B.S.; Liljefors, T.; Löfstedt, C.; Prestwich, G.D.; Sun, W.-C.; Svensson, M. Electrophysiological and behavioral responses of turnip moth males, Agrotis segetum to fluorinated pheromone analogs. J. Chem. Ecol. 1993, 19, 143–157. [Google Scholar] [CrossRef]
- Wu, W.; Cottrell, C.B.; Hansson, B.S.; Löfstedt, C. Comparative study of pheromone production and response in Swedish and Zimbabwean populations of turnip moth, Agrotis segetum. J. Chem. Ecol. 1999, 25, 177–196. [Google Scholar] [CrossRef]
- Xiang, Y.-Y.; Yang, M.-F.; Li, Z.-Z. Sex pheromone components of the female black cutworm moth in China: Identification and field trials. Zool. Res. 2009, 30, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Priesner, E.; Witzgall, P. Modification of pheromonal behaviour in wild Coleophora laricella male moths by (Z)-5-decenyl acetate, an attraction-inhibitor. Z. Angew. Entomol. 1984, 98, 118–135. [Google Scholar] [CrossRef]
- Chang, H.; Liu, Y.; Ai, D.; Jiang, X.; Dong, S.; Wang, G. A pheromone antagonist regulates optimal mating time in the moth Helicoverpa armigera. Curr. Biol. 2017, 27, 1610–1615. [Google Scholar] [CrossRef] [PubMed]
- Klun, J.A.; Baranchikov, Y.N.; Mastro, V.C.; Hijji, Y.; Nicholson, J.; Ragenovich, I.; Vshivkova, T.A. A sex attractant for the siberian moth Dendrolimus superans sibiricus (Lepidoptera: Lasiocampidae). J. Entomol. Sci. 2000, 35, 158–166. [Google Scholar] [CrossRef]
- Kong, X.; Zhao, C.; Gao, W. Identification of sex pheromones of four economically important species in genus Dendrolimus. Chin. Sci. Bull. 2001, 46, 2077–2081. [Google Scholar] [CrossRef]
- Kong, X.-B.; Zhao, C.-H.; Wang, R. Sex pheromone of the larch caterpillar moth, Dendrolimus superans, from northeastern China. Entomol. Exp. Appl. 2007, 124, 37–44. [Google Scholar] [CrossRef]
- Zhao, C.; Li, Q.; Guo, X.; Wang, X. New components of sex pheromone in the pine caterpillar moth, Dendrolimus punctatus: Identification of chemical structures and field tests. Acta Entomol. Sin. (Kunchong Xuebao) 1993, 36, 247–250. [Google Scholar]
- Kong, X.-B.; Liu, K.-W.; Wang, H.-B.; Zhang, S.-F.; Zhang, Z. Identification and behavioral evaluation of sex pheromone components of the chinese pine caterpillar moth, Dendrolimus tabulaeformis. PLoS ONE 2012, 7, e33381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, K.-W. Sex Pheromone Identification of Chinese Pine Moth Dendrolimus tabulaeformis and the Release Rates of Pine Caterpillar Moths Pheromones from Dispensers. Master’s Thesis, Shandong Agricultural University, Taian, China, 2012. [Google Scholar]
- Kong, X.-B.; Sun, X.-L.; Wang, H.-B.; Zhang, Z.; Zhao, C.-H.; Booij, K.J.H. Identification of components of the female sex pheromone of the simao pine caterpillar moth, Dendrolimus kikuchii Matsumura. J. Chem. Ecol. 2011, 37, 412–419. [Google Scholar] [CrossRef]
- Kong, X.-B.; Zhao, C.-H.; Sun, Y.-P.; Feng, S.-Q.; Wu, H.-S. Identification of minor components of the pine caterpillar moth, Dendrolimus spectabilis sex pheromone: Components, electrophysiological activity and field effects. Acta Entomol. Sin. 2003, 46, 131–137. [Google Scholar]
- Pletnev, V.A.; Ponomarev, V.L.; Vendilo, N.V.; Kurbatov, S.A.; Lebedeva, K.V. Search of the pheromone of Siberian silkworm Dendrolimus superans sibiricus (Lepidoptera: Lasiocampidae). Agrokhimiya 2000, 6, 67–72. [Google Scholar]
- Kong, X.; Zhao, L.; Zhang, Z.; Wang, H.; Bai, F.; Yu, G. Solid-phase microextraction of pheromone components of Dendrolimus houi(Lepidoptera:Lasiocampidae) and their characterization by gas chromatography—Mass spectrometry and GC-electroantennographic detection. Acta Entomol. Sin. (Kunchong Xuebao) 2010, 53, 857–863. [Google Scholar]
- Kong, X.; Zhang, Z.; Wang, H.; Zhao, C. Study on trace component in sex pheromones of Dendrolimus spp. Chin. J. Chromatogr. (Se Pu) 2005, 23, 370–373. [Google Scholar]
- Laboratory_of_Insect_Pheromone. Isolaton, identification and synthesis of EAG-active components of the pine catepillar moth sex pheromone. Chin. Sci. Bull. 1979, 24, 1004–1008. (In Chinese) [Google Scholar] [CrossRef]
- Zhao, C.-H.; Li, Q.; Gao, W. Stimulation of sex pheromone production by PBAN-like substance in the pine caterpillar moth, Dendrolimus punctatus (Lepidoptera: Lasiocampidae). Arch. Insect Biochem. Physiol. 2002, 49, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Ando, T.; Vu, M.H.; Yoshida, S.; Takahashi, N.; Tatsuki, S.; Katagiri, K.; Yamane, A.; Ikeda, T.; Yamazaki, S. (5Z, 7E)-5,7-Dodecadien-1-ol: Female sex pheromone of the pine moth Dendrolimus spectabilis Butler. Agric. Biol. Chem. Tokyo 1982, 46, 709–715. [Google Scholar] [CrossRef] [Green Version]
- Howse, P.E. Factors controlling responses of insects to pheromones. In Insect Pheromones and Their Use in Pest Management; Howse, P., Stevens, I., Jones, O., Eds.; Chapman & Hall: London, UK, 1998; pp. 69–104. [Google Scholar]
- Trona, F.; Anfora, G.; Balkenius, A.; Bengtsson, M.; Tasin, M.; Knight, A.; Janz, N.; Witzgall, P.; Ignell, R. Neural coding merges sex and habitat chemosensory signals in an insect herbivore. Proc. R. Soc. Lond. B Biol. Sci. 2013, 280, 20130267. [Google Scholar] [CrossRef] [Green Version]
- Sukovata, L.; Dziuk, A.; Parratt, M.; Bystrowski, C.; Dainton, K.; Polaszek, A.; Moore, R. The importance of trap type, trap colour and capture liquid for catching Dendrolimus pini and their impact on by-catch of beneficial insects. Agric. For. Entomol. 2020, 22, 319–327. [Google Scholar] [CrossRef]
- Levi-Zada, A.; Fefer, D.; Anshelevitch, L.; Litovsky, A.; Bengtsson, M.; Gindin, G.; Soroker, V. Identification of the sex pheromone of the lesser date moth, Batrachedra amydraula, using sequential SPME auto-sampling. Tetrahedron Lett. 2011, 52, 4550–4553. [Google Scholar] [CrossRef]




| Components | Lure Composition (mg per Dispenser) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| STD b | MD10 | MD12 | MD14 | MD15 | MD16 | MD17 | MD18 | MD17OAc | |
| (Z5,E7)-12:Ald | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 | 0.6 |
| (Z5,E7)-12:OH | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 | 0.4 |
| (Z5)-12:Ald | - | 0.15 | 0.15 | - | 0.15 | 0.15 | 0.15 | - | 0.15 |
| (Z5)-12:OH | - | 0.15 | 0.15 | 0.15 | - | 0.15 | 0.15 | - | 0.15 |
| (Z5)-10:OH | - | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | - | 0.05 | - |
| (Z5)-14:OAc | - | 0.15 | 0.15 | 0.15 | 0.15 | - | 0.15 | 0.15 | 0.15 |
| (Z5)-10:OAc | - | - | - | - | - | - | - | - | 0.15 |
| SPEO | - | 1 | - | - | - | - | - | - | - |
| Field Expt 1 | |||||||||
| Field Expt 2 | |||||||||
| Field Expt 3 | |||||||||
| Field Expt 4 | |||||||||
| Category Index i | Behavior Category Ri | Category Weight, wi |
|---|---|---|
| 1 | No reaction | 0 |
| 2 | Wing-fanning | 1 |
| 3 | Flight initiation | 4 |
| 4 | Approaching the lure, departure | 9 |
| 5 | Landing close to the lure, departures, returns | 16 |
| 6 | Landing close to the lure and staying there for a long time (>10 s) | 25 |
| 7 | Landing directly on the lure, attempt to copulate | 36 |
| Compound | D. pini | D. houi | D. kikuchii | D. punctatus | D. spectabilis | D. superans | D. superans sibiricus | D. tabulaeformis |
|---|---|---|---|---|---|---|---|---|
| (Z5)-10:OAc | p | |||||||
| (Z5)-12:Ald | p | r [62,63] | ||||||
| (Z5)-12:OH | p | r [45] c | r [64] c | r [62,63] | r [65,66] | |||
| (Z5)-12:OAc | r [67] | r [64] c | r [65,66] | |||||
| (Z5,E7)-12:Ald | r [10,11], p | r [62,68] | r [62,63] | r [61] ac, r [69] | ||||
| (E5,Z7)-12:Ald | r [45,70] c | |||||||
| (E5,E7)-12:Ald | p b | |||||||
| (Z5,E7)-12:OH | r [11], p | r [45] c | r [62,67,71] c | r [64,72,73] | r [37,62,68,74] | r [62,63] | r [61] ac, r [69] | r [62,65,66] |
| (E5,Z7)-12:OH | r [45,70] | |||||||
| (E5,E7)-12:OH | r [45] c | r [37] c, r [74] | ||||||
| (Z5,E7)-12:OAc | r [45] c | r [62,67,71] | r [64,72,73] | r [62,68] | r [69] c | r [62,65,66] | ||
| (E5,Z7)-12:OAc | r [45,70] | |||||||
| (Z5,Z7)-12:OAc | r [45] c | |||||||
| (E5,E7)-12:OAc | r [45] c | |||||||
| (Z5,E7)-12:OPr | r [64,72,73] | r [68] c | r [62,65] | |||||
| (Z5)-14:OAc | p |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rudziński, K.J.; Staszek, D.; Asztemborska, M.; Sukovata, L.; Raczko, J.; Cieślak, M.; Kolk, A.; Szmigielski, R. Newly Discovered Components of Dendrolimus pini Sex Pheromone. Insects 2022, 13, 1063. https://doi.org/10.3390/insects13111063
Rudziński KJ, Staszek D, Asztemborska M, Sukovata L, Raczko J, Cieślak M, Kolk A, Szmigielski R. Newly Discovered Components of Dendrolimus pini Sex Pheromone. Insects. 2022; 13(11):1063. https://doi.org/10.3390/insects13111063
Chicago/Turabian StyleRudziński, Krzysztof J., Dorota Staszek, Monika Asztemborska, Lidia Sukovata, Jerzy Raczko, Marek Cieślak, Andrzej Kolk, and Rafał Szmigielski. 2022. "Newly Discovered Components of Dendrolimus pini Sex Pheromone" Insects 13, no. 11: 1063. https://doi.org/10.3390/insects13111063